上海大金山島植被功能組成對草本植物、土壤動物和細菌多樣性的影響
2023-09-11 08:42:42楊柏鈺賽碧樂楊安娜張增可李曾燕王萬勝杜運才于再平閻恩榮
生態學報 2023年16期
關鍵詞:物種
楊柏鈺,賽碧樂,楊安娜,張增可,李曾燕,張 立,牟 凌,王萬勝,杜運才,劉 海,于再平,郭 超,閻恩榮,*
1 華東師范大學生態與環境科學學院,上海 200241 2 上海市金山區海洋海塘管理所,上海 201508 3 浙江舟山市普陀山林場, 舟山 316107
生物多樣性是生態系統功能形成的基礎[1]。在森林生態系統中,生態系統功能是由不同營養級生物多樣性共同維持的[2—3]。植物多樣性決定了生態系統生產力大小和物質返還量[4],土壤動物和微生物多樣性驅動了凋落物分解和養分循環過程[5]。在此過程中,由于上行效應和下行效應,不同營養級或同一營養級內生物間也會相互影響[6—7]。例如,樹木多樣性既會通過改變群落結構組成、林下光線與水分資源分配而改變草本多樣性[8—9],也會通過改變輸入的葉片與枯木凋落物的數量與質量而改變土壤動物和微生物多樣性[10—13];同樣,由于負反饋作用,土壤動物和微生物多樣性也會通過調節養分周轉和土壤肥力,反過來影響草本和樹木多樣性[14—15]。因此,植物多樣性與土壤動物和微生物多樣性之間存在著密切的生態關聯[16—17]。但是,我們對于不同營養級生物多樣性相互關系及其驅動機制的理解仍處在探索階段[18]。
過去二十多年中,生態學家探索了不同生態系統中草本或木本植物多樣性對土壤動物及微生物多樣性的影響[19—22]。研究發現,在溫帶草原中,由于食物資源的互補效應,隨著植物多樣性的增加,蚯蚓和線蟲等土壤動物多樣性顯著增加[5];在干旱荒漠生態系統中,木本植物豐度促進了真菌等微生物多樣性[20]。總體而言,雖然這些研究極大的提升了我們關于不同營養級生物多樣性間相互作用的理解,但卻往往從單一營養級的簡單因果關聯的角度出發,忽略了多營養級之間的相互反饋[6—7, 23]。同時,根據生物級聯效應(cascading effect),上一營養級對下一營養級生物的作用過程可能同時包括了直接影響與間接調控[24]。然而,目前關于生物級聯效應對多營養級多樣性關系的影響仍沒有明確的研究結論。
植物多樣性對土壤動物和微生物多樣性的影響主要取決于其返回土壤的凋落物基質的數量、質量與多樣性[12],也就是說,植物多樣性的功能屬性組成是調控土壤動物和微生物多樣性的主要因素之一[25—27]。研究表明,在熱帶和亞熱帶森林中,常綠與落葉樹種的占比既會通過改變林下光斑大小,從而影響草本多樣性[28—29],也會通過改變凋落物基質組成多樣性,從而影響土壤動物和微生物多樣性[10, 30—31]。同時,部分研究表明,樹木物種多樣性對土壤動物與微生物的影響較弱[32]。基于此,本文預測,相對于樹木物種多樣性而言,亞熱帶森林中落葉樹木的占比(也就是功能組成)在調控林下草本、土壤動物與微生物多樣性方面扮演更為重要的角色。由于不同營養級生物間既存在上行效應,也存在下行效應[6],加之級聯效應所產生的直接與間接調控[24],本文提出科學預測1: 森林群落樹木的功能組成(即落葉樹木占比)既可以直接影響土壤動物和微生物多樣性,也可以通過草本多樣性的正向級聯效應而間接調控土壤動物和微生物多樣性。
另外,作為腐食食物鏈的主要構成,土壤動物和微生物群落間也存在著緊密的競爭、互惠合作與捕食關系[33—34]。先前基于亞熱帶森林的研究表明,土壤無脊椎動物可通過啃食樹木木質殘體而為微生物提供食物來源[11]。因此,本文進一步預測(科學預測2):群落中落葉木本植物占比的增加會加劇土壤動物與微生物之間的負向關系。具體而言,由于存在食物資源競爭關系,因此,隨土壤動物多樣性增加,微生物多樣性下降。
上海大金山島自1993年建立保護區起,封山育林30余年,探究植被功能組成和物種多樣性對土壤動物及細菌多樣性的調控作用,以及土壤動物和細菌多樣性對植被物種多樣性的反饋影響,對于揭示森林多營養級生物多樣性的保育具有重要意義。綜上,為檢驗所提出的科學預測,本研究以上海大金山島包含不同落葉木本植物占比的13個植物群落為對象,在分析植物群落樹木物種多樣性、草本物種多樣性、土壤動物和細菌多樣性關系的基礎上,探索土壤養分梯度上,群落中落葉樹木占比對草本、土壤動物和細菌多樣性的影響效應,從而為保護大金山島的生物多樣性提供理論支撐。
1 方法
1.1 研究區概況和樣地選擇
大金山島(30°41′42″ N, 121°24′25″ E)地處上海金山區附近的杭州灣, 位于中亞熱帶北緣, 是金山三島海洋生態自然保護區的核心區。大金山島總面積為0.229 km2, 主峰高103.4 m, 其年平均氣溫15.8 ℃, 土壤為褐色山地黃壤, 降水充沛, 屬海洋性氣候。大金山島植被從外貌水平可劃分為草甸、灌草叢、落葉闊葉林、落葉常綠闊葉混交林和常綠闊葉林5個類型,群落物種組成中落葉植物按一定比例下降[35]。
2016年7月,選擇灌草叢、落葉闊葉林、落葉常綠闊葉混交林和常綠闊葉林植被類型,根據分布面積大小,選擇其中的典型植物群落建立了13個長期固定觀測樣地,落葉闊葉灌叢包括3個日本野桐(Mallotustenuifolius)群落樣地和1個小葉女貞(Ligustrumquihoui)群落樣地,落葉闊葉林包括麻櫟(Quercusacutissima)群落、樸樹(Celtissinensis)和絲棉木(Euonymusmaackii)群落樣地各1個,常綠落葉闊葉混交林群落包括椿葉花椒(Zanthoxylumailanthoides)-柃木(Euryajaponica)群落樣地1個,常綠闊葉林包括普陀樟 (Cinnamomumjaponicum)群落和紅楠(Machilusthunbergii)群落樣地各1個,青岡(Cyclobalanopsisglauca)群落樣地3個。其中,由于小葉女貞群落分布范圍小,樣地面積為100 m2,其余群落類型的樣地面積均為400 m2。另外,由于日本野桐群落和青岡群落分布面積較廣,根據坡位和地形變化,每個類型各建立了3個同樣面積的長期監測樣地。
樣地建成后,根據統計樣方法對每個樣地進行群落學特征調查,對于胸徑大于1 cm,且株高大于0.5 m的所有木本植物,逐木確定物種名稱,測量胸徑、樹高與冠幅。對于草本層,按照典型樣地調查法,確定物種組成,記錄各物種的個體數、平均蓋度與株高。各樣地的群落學特征見表1。
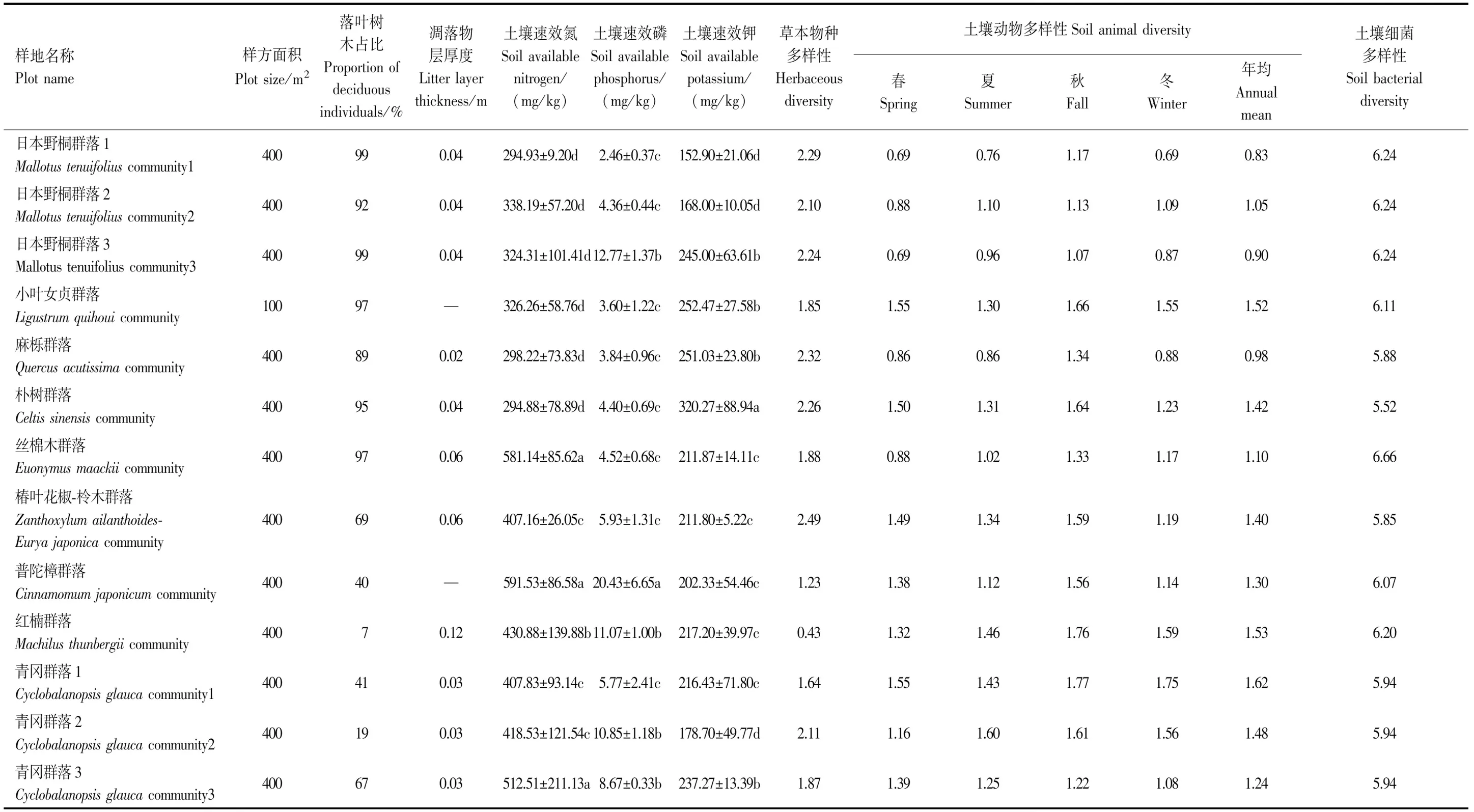
表1 大金山島13個典型植被樣地的基本特征
1.2 土壤理化因子測定及土壤無脊椎動物群落調查
在完成植物群落特征調查后,在每個樣地中,隨機設置8—10個采樣點,利用土鉆提取0—20 cm土樣,帶回實驗室自然風干后進行土壤理化性質分析。樣品風干后,去除石粒和根系等植物殘體,用研缽磨碎,過60目網篩,用以測定土壤速效養分。土壤速效鉀含量采用乙酸銨浸提-火焰光度法測定,土壤速效磷和速效氮含量分別采用氟化銨-鹽酸浸提法和聯合浸提-比色法進行測定[36]。
本文在2017年春季(3月)、夏季(6月)、秋季(9月)和冬季(12月),分四個季節調查土壤動物群落。采樣時,在每個固定樣地中,按照梅花樁法設置了5個采樣點,利用直徑為5 cm的不銹鋼環刀,對表層土壤(0—10 cm)取樣,然后將土柱放至自封袋,在4 ℃條件下運回實驗室處理。在實驗室,利用Tullgren漏斗法(25 w,72 h)分離中小型土壤動物[37]。分離出的土壤動物保存在75%的酒精中,利用體式顯微鏡(Leica, DVM6c, Germany)鑒定和計數。根據尹文英等,所有土壤動物鑒定至目水平[38—39]。
1.3 土壤細菌群落調查
在2017年夏季(6月)土壤動物取樣的同時,也按照梅花樁5點取樣法,采集土壤細菌調查土樣。取樣時,去除土壤表面葉片與枯枝凋落物,利用土鉆收集表層0—10 cm土樣,利用4℃保溫盒快速運回實驗室。在實驗室,每個樣地的5份土樣混合,過2 mm篩,再選取3份,采用MoBio powerSoil DNA提取試劑盒(MoBio Laboratories, Carlsbad, CA, USA)提取土壤微生物DNA,并以16S核糖體rRNA基因V4區為目標區域,采用引物515F(5′-GTGCCAGCMGCCG CGGTAA-3′)和806R (5′-GGACTACHVGGGTWTCTA AT-3′) 進行PCR擴增。PCR反應程序為在94 ℃溫度條件下擴增35個循環,其中包括在94 ℃溫度條件下變性10 s以及在52 ℃溫度條件下復性30 s,最后在72 ℃溫度條件下延伸45 s,共計5 mins。最后保持72 ℃溫度條件不變,將產物延伸10 mins。將得到的PCR產物純化并在llumina Nova 6000平臺上進行擴增子測序。使用Usearch v10軟件將原始數據進行拼接和質控,使用代碼“-fastq_mergepairs”合并雙端序列,使用cutadapt軟件去掉序列中的引物,將長度小于300 bp的序列和低質量序列舍棄,最后去除重復序列,得到有效拼接片段。隨后使用UPARSE算法對超過97%相似度閾值的OTU (Operational Taxonomic Units)進行分類學分析,其中,通過檢測并刪除嵌合體,形成了最初的OTU表并比對SILVA 132數據庫,采用RDP分類器(置信閾值為0.8)對每個OTU進行物種注釋。為保持測序深度的一致,對樣品的序列總數進行抽平處理,抽平后每個樣品的細菌序列總數為30610條。最后,得到了OTU水平上的細菌群落組成與結構信息。
1.4 群落落葉成分比例和生物多樣性指數計算
本研究選取落葉樹木個體數占群落總個體數的占比(DP),用以反映群落中樹木功能成分比例對不同營養級生物多樣性的影響,計算依據公式1:
(1)
式中,Nd/e為每個樣方中落葉樹種/常綠樹種個體數,N為每個樣方中樹木總個體數。
如公式2所示,本研究采用香濃維納指數(Shannon-Wiener,H′)反映樹木和草本多樣性、土壤動物多樣性及細菌多樣性。
(2)
式中,Pi=Ni/N,Ni為第i物種的相對多度,N是所有物種的總個體數。
1.5 統計分析
首先,利用Pearson相關分析檢驗了土壤養分含量、樹木物種多樣性、落葉樹木占比、草本物種多樣性、土壤動物多樣性與細菌多樣性之間是否具有關聯性。其中,前人研究表明[16,40],土壤速效養分常常集中分布在土壤表層(0—20 cm),且在相鄰年際間差異較小,因此為了捕捉到群落整體的土壤速效養分情況對多營養級多樣性的調控作用,本文將土壤速效養分、落葉木本植物占比、土壤動物多樣性、微生物多樣性放在一起探索其關聯[16—17]。隨后,為了探究林分不同生物類群多樣性之間的相互作用,利用結構方程模型(Structural Equation Modeling, SEM)探究群落落葉樹種比例如何通過改變草本層物種多樣性而間接影響土壤動物多樣性及細菌多樣性。前人研究表明,雖然森林生態系統中上行效應與下行效應并存,但往往以上行效應為主[7]。因此,本文SEM中以木本植物與草本植物等低營養級物種多樣性作為效應變量,以土壤動物與細菌等高營養級類群作為響應變量。同時,由于同一營養級間的相互作用,且占據空間資源的優勢類群作用更大,SEM包括了木本植物多樣性對草本物種多樣性的直接影響路徑,以及土壤動物多樣性對細菌多樣性的直接影響路徑[7]。考慮到不同營養級間的級聯作用,SEM納入了木本植物通過草本物種多樣性對土壤動物與細菌的間接路徑[24]。另外,土壤養分是影響各營養級生物多樣性的主要因素之一,本文分析發現,落葉樹種占比與土壤速效磷顯著相關,而與速效氮和速效鉀無顯著關系(詳見結果2),因此,SEM結構中,土壤速效磷也被納入作為影響各營養級生物多樣性的效應變量。最后,由于土壤動物的季節變化較大,為更具代表性,本研究SEM選擇年均土壤動物多樣性(H′)。
模型分析過程中,依據Pearson相關分析結果作為模型變量篩選準則,選擇與不同類群物種多樣性相關關系顯著(P<0.10)且相關系數值較大的變量進入SEMs。本文利用信息標準指數(AIC),卡方擬合優度檢驗(P),擬合優度指數(GFI)和近似誤差的均方根(SRMR)等作為SEMs擬合優度的評估指標[41]。當AIC值較小,P> 0.05,GFI >0.90,SRMR<0.08時表明方程擬合結果較好且可信度較高。在對所有變量進行分析前,對數據進行了對數轉換(ln)。所有的數據分析均在R4.1.2中進行。
2 結果
2.1 植物群落功能組成特征和土壤養分含量
如表1所示,13個群落的落葉木本植物占比變化范圍較大(7%—99%),其中日本野桐群落1,3占比最高,小葉女貞群落、絲棉木群落和樸樹群落次之,紅楠群落最低。凋落物層厚度變化范圍在0.02—0.12 m之間。土壤速效氮和速效磷含量在普陀樟群落最高,紅楠群落次之,日本野桐群落1最低。土壤速效鉀含量在樸樹群落最高,小葉女貞群落和麻櫟群落次之,日本野桐群落1最低。
2.2 落葉木本植物占比、草本物種多樣性、土壤動物多樣性與細菌多樣性間的關系
由表2可見,土壤速效磷含量與落葉木本植物占比顯著正相關(P<0.05),土壤速效氮和速效鉀含量與落葉木本植物占比無顯著相關性(P>0.10)。土壤速效養分也與各生物類群多樣性無顯著關聯性(P>0.10)。
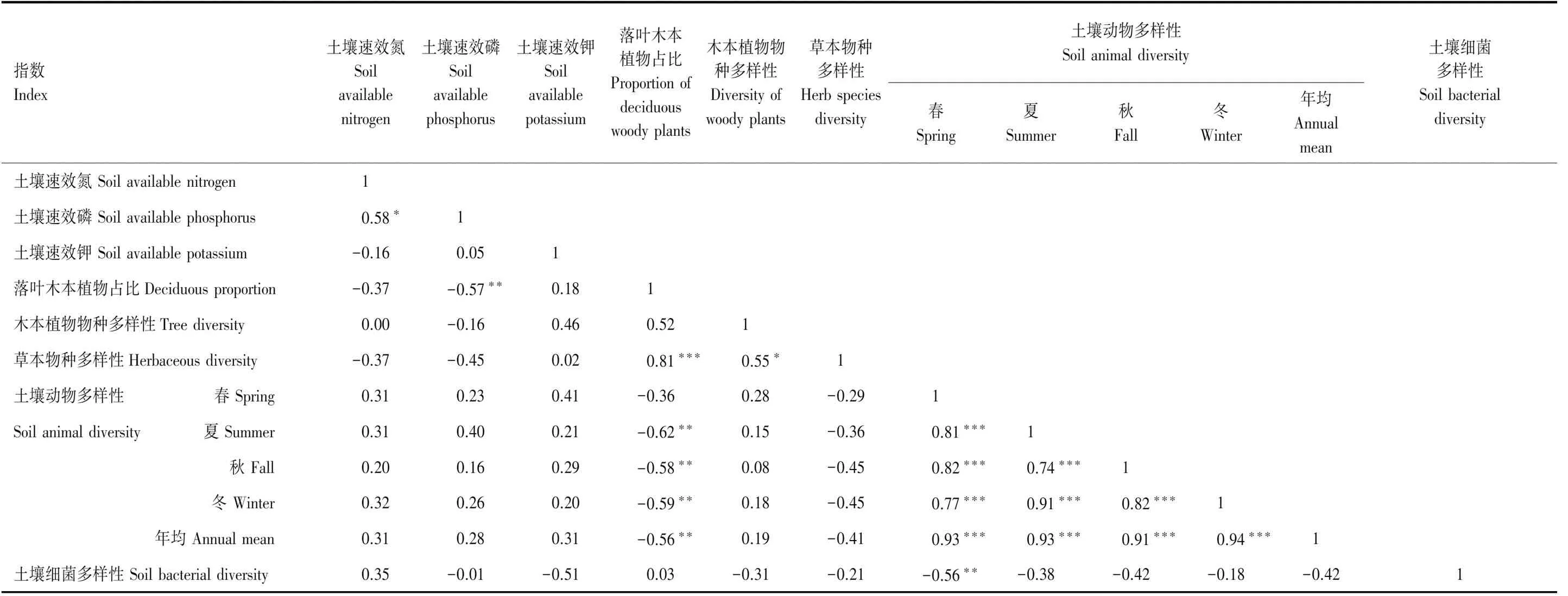
表2 土壤速效養分、落葉木本植物占比和物種多樣性、草本物種多樣性、土壤動物和細菌多樣性之間的相關關系
落葉木本植物占比與草本物種多樣性顯著正相關(P<0.01),與土壤類群多樣性(夏、秋、冬季和年均)顯著負相關(P<0.05),但與春季土壤動物類群多樣性和細菌多樣性無顯著相關性(P>0.10)。木本植物多樣性僅與草本物種多樣性顯著正相關(P<0.10),而與土壤動物和細菌多樣性無顯著相關性(P>0.10)。草本物種多樣性也與土壤動物和細菌多樣性無顯著相關關系(P>0.10),土壤動物多樣性與細菌多樣性間無顯著關系(P>0.10)。
2.3 群落落葉木本植物占比對土壤動物和細菌多樣性的直接和間接影響
SEM方程模型擬合效果較好(AIC=-8.438;P=0.845; GFI=0.988; SRMR=0.022),分別解釋了群落落葉樹木占比、草本層多樣性、土壤動物多樣性與細菌多樣性變異的33%,66%,32%和34%(圖1)。如圖1和表3所示,土壤速效磷含量的增加顯著抑制群落落葉樹木占比(P<0.05),而對草本層、土壤動物與細菌多樣性無顯著影響(P>0.10)。落葉木本植物占比的增加顯著促進草本層多樣性,顯著抑制土壤動物多樣性(P<0.05);雖然落葉木本植物占比對土壤細菌多樣性無直接影響,但間接通過增加草本層多樣性而顯著抑制細菌多樣性(r=-0.47,P<0.05)。此外,草本多樣性與土壤動物多樣性均會直接抑制細菌多樣性(P<0.10)。需要指出的是,本研究也擬合了草本物種多樣性與土壤動物多樣性間的作用路徑,但模型被拒絕,間接說明草本物種多樣性對土壤動物多樣性的直接影響遠遠小于落葉木本植物占比的影響。
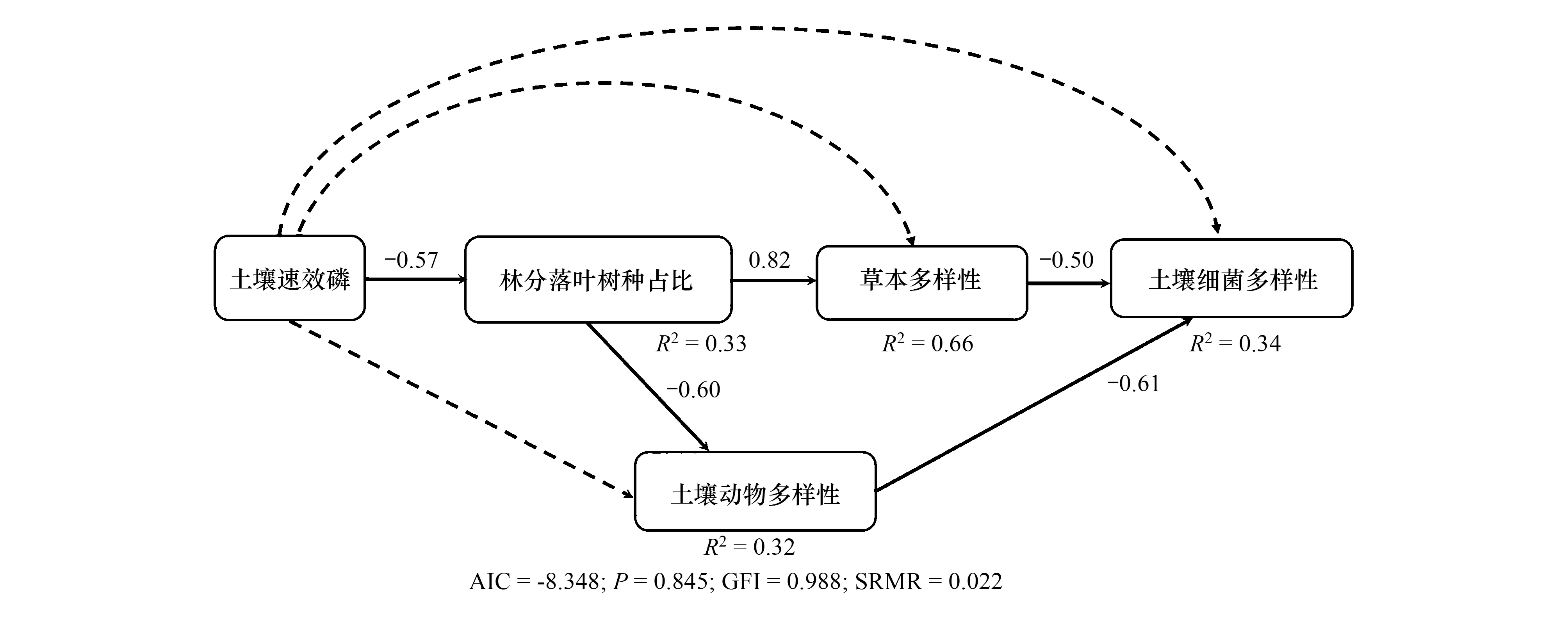
圖1 群落土壤速效磷和落葉木本植物占比對草本、土壤動物與細菌多樣性的直接與間接影響Fig.1 Direct and indirect effects of soil available phosphorus content and proportion of deciduous woody plants on herbaceous species diversity, soil animal diversity and soil bacterial diversityAIC:信息標準指數;P:卡方擬合優度檢驗;GFI:擬合優度指數;SRMR:近似誤差的均方根

表3 土壤速效磷、群落落葉樹木占比對草本、土壤動物與細菌多樣性的影響路徑和效應值
3 討論
本研究發現,大金山島植被中,落葉木本植物占比不僅對草本層物種多樣性和土壤動物多樣性分別產生直接的正和負效應,也會通過增加草本植物多樣性而間接降低土壤細菌多樣性。該結果支持本研究的科學假說,即:相對于樹木物種多樣性而言,亞熱帶森林的落葉樹木占比(也就是群落落葉種功能組成)在調控林下草本、土壤動物與細菌多樣性方面扮演著更為重要的角色。事實上,大金山島位于中亞熱帶北緣,地帶性植被類型為常綠落葉闊葉混交林和落葉闊葉林,與低緯度的典型常綠闊葉林不同,落葉物種是大金山島植被組成的主體成分,是群落結構和生態過程的主要塑造者[42]。本文從多營養級角度證實了落葉木本植物占比對森林不同營養級多樣性關系的調控作用,落葉樹種占比是亞熱帶東部海島生態系統不同營養級生物多樣性維持的主要驅動因素[43]。
落葉木本植物占比與土壤動物多樣性間的負向關系、草本物種多樣性與土壤細菌多樣性的負向關系以及土壤動物和細菌多樣性間的負向關系均反應了低營養級生物對高營養級生物密切的上行調控作用[7]。落葉木本植物占比對土壤動物多樣性的負效應可能是源于選擇效應(selection effect)[5, 44—45],即:雖然落葉植物的群落優勢效應可返還大量基質質量高的凋落物到地表,但降低了凋落物基質多樣性,這樣的格局有利于喜食高質量凋落物的專性類群的生長與繁殖,而不利于維持食性更為多樣的土壤動物類群[46]。結合本研究中木本植物多樣性與土壤動物多樣性間無顯著關聯的結果,可以得出落葉木本植物占比所主導的選擇效應決定了土壤動物多樣性的分布格局。
本研究發現,落葉木本植物占比并不直接影響土壤細菌多樣性,而是間接通過增加草本多樣性而抑制土壤細菌多樣性。一方面,該結果說明,落葉木本植物占比對土壤細菌多樣性的上行調控更多來自于其促進草本植物多樣性之后的生物級聯效應[7],即:間接效應,另一方面說明,草本植物多樣性是塑造大金山島土壤細菌多樣性的主要驅動力。本研究中,落葉木本植物占比與草本物種多樣性存在積極的關聯格局,其主要原因是,隨著落葉木本植物占比的增多,尤其在非生長旺季,群落內的透光性增強,這樣可促進陽性草本植物的適合度,從而增加草本植物物種多樣性[28—29]。本文在調查中發現,相較于青岡群落樣地1,2,3,落葉木本植物占比較高的野桐群落1,3及小葉女貞群落的陽性植物居多,例如絡石、酢漿草與求米草等。同時本文也發現,木本植物物種多樣性的增加會顯著促進與草本植物物種多樣性,這也從另一個側面說明,木本植物物種多樣性的增加會增加林下光資源的異質性,從而有利于維持光適應策略更為多樣的草本植物[8, 47]。那么,隨著草本物種多樣性的增加,土壤養分資源消耗過快,加劇了不同細菌種類之間的養分競爭[48],使得細菌多樣性下降,因此草本物種多樣性與土壤細菌多樣性間表現出了負向關系。
落葉木本植物比例增加所導致的林下草本植物繁榮不僅加劇了不同土壤細菌間的養分資源競爭,也可能會加劇土壤動物與細菌的養分競爭[11, 34]。本研究發現,土壤動物與細菌多樣性間存在顯著的負相關關系,這一格局說明,土壤動物與細菌間不僅僅存在互利作用,即正相關關系,也存在資源競爭格局[7, 11]。但在這里需要指出,土壤動物對細菌多樣性的抑制作用也可能與土壤動物與細菌的食性有關。由于本文土壤動物的鑒定水平僅為目水平,無法深入探討不同食性的土壤動物與細菌之間的資源競爭格局。綜上,落葉木本植物占比的增加不僅會顯著改變群落內部的光線供給性,從而影響草本物種多樣性格局[28],更會通過改變凋落物基質多樣性而調控土壤養分周轉和供給等功能,從而改變土壤動物和細菌多樣性格局[10, 30—31]。
4 結論
本研究表明,落葉木本植物占比是大金山島植被系統不同營養級生物多樣性的主要塑造者,其通過選擇效應顯著抑制土壤動物多樣性,但會通過增加林下透光性而提升草本多樣性,并通過促進草本多樣性而間接減少土壤細菌多樣性。該研究結果有助于深入理解中亞熱帶北緣森林生態系統多營養級生物多樣性的維持機制。需要指出,考慮到本研究土壤動物的鑒定水平,加之本研究結構方程模型分析顯示,土壤速效養分含量并未對土壤動物和細菌多樣性產生直接和間接影響,而只影響落葉木本植物占比,因此,未來有關土壤動物和細菌多樣性負向關系的維持機制需要使用分類水平更高的多營養級生物多樣性數據來進行更深入的探索。最后,本研究僅從數量關系角度揭示了各營養級生物多樣性間(alpha多樣性)的關聯性,未來應考慮利用控制實驗等手段確認落葉木本植物占比在森林生物多樣性(alpha, beta多樣性)維持中的生態機制。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
