厭氧處理條件下紫娟茶化學成分的變化及其對體外ACE活性的影響
2023-08-30 14:05:38楊高中施江張悅彭群華林智呂海鵬
茶葉科學 2023年4期
楊高中,施江,張悅,彭群華,林智*,呂海鵬*
厭氧處理條件下紫娟茶化學成分的變化及其對體外ACE活性的影響
楊高中1,2,施江1,張悅1,彭群華1,林智1*,呂海鵬1*
1. 中國農業科學院茶葉研究所/農業農村部特種經濟動植物生物學與遺傳育種重點實驗室,浙江 杭州 310008;2. 中國農業科學院研究生院,北京 100081
以紫娟茶樹鮮葉為研究對象,對厭氧處理后制備的凍干樣、綠茶、紅茶、白茶等茶樣中主要非揮發性成分(包括兒茶素、氨基酸、花青素等)進行測定,并采用高效液相色譜法評估其體外血管緊張素轉換酶(Angiotensin-converting enzyme,ACE)抑制活性;此外,嘗試通過分子對接模擬試驗研究茶葉中主要化學成分和ACE之間潛在的互作機制。結果表明,經厭氧處理后的凍干樣中-氨基丁酸(-aminobutyric acid,GABA)含量增加至1.72?mg·g-1,顯著高于未厭氧處理茶樣的0.04?mg·g-1(<0.05)。厭氧處理后不同的加工工藝對茶樣中的氨基酸和多酚類化合物的組成及含量影響較大;采用白茶工藝能提高茶樣中氨基酸組分含量,而凍干工藝能最大程度保留茶樣中多酚類化合物含量。紫娟茶樹鮮葉樣品經厭氧處理后,體外ACE抑制活性顯著增強(<0.05);厭氧處理后采用冷凍干燥工藝所制樣品的ACE抑制活性最強,顯著高于采用綠茶、白茶和紅茶加工工藝制作的樣品(<0.05)。此外,分子對接模擬試驗發現,茶葉中的主要化學成分與ACE的相互作用強度與其分子結構中的親水基團和芳香環結構有關。研究結果有助于揭示厭氧處理對紫娟茶ACE抑制活性的影響,為開發具有潛在治療高血壓等特殊功能型紫娟茶產品提供理論依據。
茶;-氨基丁酸;厭氧處理;加工工藝;血管緊張素轉換酶
高血壓是心血管疾病和過早死亡的一個重要的可改變的風險因素。從1990年到2019年,全球30~79歲的高血壓人群幾乎翻了一番[1]。血管緊張素轉換酶(Angiotensin-converting enzyme,ACE)是腎素-血管緊張素系統中的一個關鍵酶,負責通過血管收縮調節血管張力,其失調可能導致高血壓和心腦血管疾病[2]。抑制ACE可以有效降低血壓,減少血栓形成和心血管疾病的風險,長期以來,抑制劑對ACE的體外抑制活性被作為篩選天然ACE抑制劑的重要因素[3]。
近年來,紫芽茶以其特殊的風味品質和保健功能引起了大家廣泛關注[4]。與傳統的綠葉茶相比,紫葉茶具有較多的生物活性成分[5],其中許多是潛在的天然ACE抑制劑,包括表沒食子兒茶素沒食子酸酯(Epigallocatechin gallate,EGCG)[6]、表沒食子兒茶素-3--(3"--甲基)沒食子酸酯[Epigallocatechin 3--(3"--methyl) gallate,EGCG3"Me][7]、原花青素[8]和花青素[9]等,它們通過體外模型被證實具有顯著的ACE抑制活性。紫娟(var.)是云南省農業科學院茶葉研究所從云南大葉群體種中選育而來,是我國最具代表性的紫芽茶品種之一。此前有報道稱,在動物研究中,紫娟茶比云南大葉種茶樹鮮葉制成的茶樣具有更強的降血壓效果[10]。值得注意的是,-氨基丁酸(-aminobutyric acid,GABA)被認為是一種重要的生物活性物質,可能在預防心血管疾病,特別是降低血壓方面發揮有益的作用[11]。厭氧處理已被證實是提高茶葉產品中GABA含量的有效方法[12-13]。因此,推測經過厭氧處理后,通過增加紫娟茶中的GABA含量,可以提高其ACE抑制活性,有望開發出一些具有治療高血壓的紫娟茶產品,滿足不同茶葉消費者對健康食品的特殊需求。
本研究以紫娟茶樹鮮葉為研究對象,對厭氧處理后制備的凍干樣、綠茶、紅茶、白茶等茶樣中主要非揮發性成分(兒茶素、氨基酸、花青素等)進行測定,并采用高效液相色譜法評估其體外ACE抑制活性[14-15];嘗試通過分子對接模擬試驗研究茶葉中主要化學成分(如兒茶素、茶黃素、花青素、GABA等)和ACE之間潛在的互作機制,旨在闡明厭氧處理后不同加工工藝對紫娟茶體外ACE抑制活性的影響,為開發具有潛在降血壓的特殊紫娟茶產品提供科學依據。
1 材料與方法
1.1 材料與試劑
1.1.1 樣品制備方法
紫娟茶樹鮮葉(采摘標準為一芽三葉)于2022年6月8日采摘自中國農業科學院茶葉研究所嵊州基地。樣品的制備流程及詳細工藝參數如圖1所示,將1?kg鮮葉攤放處理8?h后,立即在液氮中預凍處理,隨后使用真空冷凍干燥機進行冷凍干燥,標記為對照組。此外,將5?kg鮮葉分裝入塑料袋,并進行真空處理8?h。隨后各取出1?kg鮮葉分別采用真空冷凍凍干以及白茶和綠茶加工工藝制備茶樣,剩余2?kg鮮葉采用紅茶加工工藝制備紅茶樣。使用研磨機將所有樣品研磨成粉末,并保存在–20?℃冰箱待進一步分析。
1.1.2 主要試劑
兒茶素(Catechin,C)、表兒茶素(Epicatechin,EC)、兒茶素沒食子酸酯(Catechingallate,CG)、表兒茶素沒食子酸酯(Epicatechingallate,ECG)、沒食子兒茶素(Gallocatechin,GC)、表沒食子兒茶素(Epigallocatechin,EGC)、沒食子兒茶素沒食子酸酯(Gallocatechingallate,GCG)、EGCG、EGCG3''Me(98.0%)、茶黃素(Theaflavin,TF)、茶黃素-3-沒食子酸酯(Theaflavin-3-gallate,TF3G)、茶黃素-3'-沒食子酸酯(Theaflavin-3'-gallate,TF3'G)、茶黃素-3,3'-雙沒食子酸酯(Theaflavin-3,3'-digallate,TFDG)、馬尿酰-組氨酰-亮氨酸(Hippuryl-histidyl-leucine,HHL)(BR,98.0%)、馬尿酸(>98.0%)、硼酸-硼砂緩沖液(pH 8.3)購自上海源葉生物技術有限公司。天冬氨酸、蘇氨酸、絲氨酸、天冬氨酸、谷氨酸、脯氨酸、甘氨酸、丙氨酸、纈氨酸、蛋氨酸、異亮氨酸、亮氨酸、酪氨酸、苯基丙氨酸、組氨酸、色氨酸、精氨酸、胱氨酸、賴氨酸、沒食子酸(Gallic acid,GA)、GABA(純度為95.0%~99.0%)、GABA(97.0%)和ACE(≥2.0?units·mg-1)購自上海Sigma-Aldrich公司。-茶氨酸(>99.0%)和甲酸(>98.0%)購自北京百靈威科技有限公司。茚三酮(固體)和檸檬酸鋰鹽緩沖液(pH分別為2.9、4.2、8.0)購自杭州津本科學儀器有限公司。磷酸(85%~90%)、氯化鉀和三氟乙酸(≥98.0%)購自上海麥克林生化科技有限公司。乙腈和甲醇(色譜純)購自德國默克公司。純凈水購自杭州娃哈哈集團有限公司。

圖1 不同工藝紫娟茶葉加工流程圖
1.2 儀器與設備
殺青機、揉捻機、烘焙提香機購于福建省安溪佳友機械有限公司,智能人工氣候室購于寧波普朗特儀器有限公司,電子天平(AB107-S型)購于瑞士Mettler Toledo公司,真空冷凍干燥機(FD5-10B型)購于美國GOLD-SIM公司,研磨機購于德國IKA公司,電熱恒溫水浴鍋(DK-S11型)購于上海森信實驗儀器有限公司,離心機(Centrifuge 5810 R型)購于德國Eppendorf公司,分光光度計(UV-3600型)、高效液相色譜系統(LC-20A型)配備WondaSil C18柱(250?mm×4.6?mm,5?μm)購于日本島津公司,超高效液相色譜系統(Acquity H-Class)配備Acquity UPLC BEH C18柱(100?mm×2.1?mm,1.7?μm)購于英國Waters公司,氨基酸分析儀(S-433D型)配備Sykam陽離子LCAK07/Li柱(4.6?mm×150?mm)購于德國Sykam公司。
1.3 方法
1.3.1 茶多酚和茶黃素分析檢測方法
茶樣中茶多酚含量的測定參考GB/T 8313—2018《茶葉中茶多酚和兒茶素類含量的檢測方法》進行;4種茶黃素類成分的含量測定參考GB/T 30483—2013《茶葉中茶黃素的測定-高效液相色譜法》進行。
1.3.2 氨基酸分析檢測方法
茶樣中氨基酸含量的測定參考Yang等[16]的方法并稍作修改,采用氨基酸分析儀進行定性定量。稱取100?mg的茶粉,加入10?mL的沸水并在100?℃下提取15?min,每間隔5?min搖勻1次。冷卻后,將混合溶液在離心機中以8?000?r·min-1離心10?min。將上清液用0.45?μm的膜過濾,并儲存在4?℃下進行氨基酸分析。氨基酸在Na+型磺酸基強酸性陽離子交換樹脂上分離。流動相由pH為2.9、4.2、8.0的檸檬酸鋰鹽緩沖液組成,流速為0.45?mL·min-1。茚三酮流速為0.25?mL·min-1,反應器溫度130?℃,進樣量為10?μL,柱溫設定為40?℃,檢測波長設定為570?nm和440?nm。采用外標法對茶樣中的氨基酸成分進行定量。
1.3.3 兒茶素、咖啡堿、GA含量分析檢測方法
茶樣中兒茶素、咖啡堿、GA含量的測定參考Yang等[16]的方法并稍作修改,采用UPLC配備Acquity UPLC BEH C18柱進行定性定量。稱取0.2?g茶粉,加入10?mL預熱的70%甲醇水溶液(∶),70?℃水浴30?min,每間隔5?min混勻1次。待冷卻后4?000?r·min-1離心5?min,用純凈水稀釋10倍后,過0.22?μm尼龍膜裝入進樣瓶待測。柱溫35?℃,進樣量5?μL,檢測波長280?nm,流速0.35?mL·min-1。溶劑A為含0.1%(∶)甲酸的水溶液,溶劑B為純甲醇。線性梯度洗脫曲線如下:0?min,3% B相;3.0?min,8% B相;7.5?min,20% B相;11.0?min,20% B相;13.0?min,60% B相;14.5?min,60% B相;15.0?min,3% B相;19.0?min,3% B相。兒茶素和咖啡堿的含量采用外標法測定。
1.3.4 花青素分析檢測方法
待測茶樣提取同1.3.1章節。茶樣中花青素總量通過pH示差法測定[17]。pH=1.0的緩沖液為0.2?mol·L-1鹽酸-氯化鉀緩沖液,pH=4.5的緩沖液為0.4?mol·L-1乙酸鈉。樣品的稀釋通過限制待測樣品在520?nm處的吸光度在0.2~0.8來確定。所有樣品分別用pH=1.0和pH=4.5緩沖液稀釋4倍。靜置50?min后用分光光度計分別在520?nm和700?nm處測量吸光度。總花青素含量(TAC;每克提取物中矢車菊素-3--葡萄糖苷當量的毫克數)按以下公式計算:
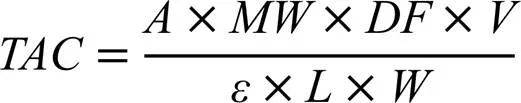
式中,為吸光度值,為稀釋系數,為最終體積(mL),為茶葉樣品的質量(g),為矢車菊素-3--葡萄糖苷的分子量(449.2?g·mol-1)。為光程(cm),為矢車菊素-3--葡萄糖苷在520?nm處的摩爾消光系數(2.69×104L·mol-1·cm-1)。比吸光度定義為:
=(1–2)–(3–4)
式中,1為樣品采用pH=1.0的緩沖液稀釋后在520?nm處測定的吸光度,2為樣品采用pH=1.0的緩沖液稀釋后在700?nm處測定的吸光度,3為樣品采用pH=4.5的緩沖液稀釋后在520?nm處測定的吸光度,4為樣品采用pH=4.5的緩沖液稀釋后在700?nm處測定的吸光度。
1.3.5 ACE抑制活性分析方法
采用HPLC方法檢測不同加工工藝茶樣的ACE抑制活性強度,參考文獻[14-15]方法并稍加修改。稱取100?mg的茶粉加入10?mL的100?℃沸水提取,沸水浴提取15?min,每間隔5?min渦旋1次。冷卻后,樣品在冷凍離心機中以8?000?r·min-1離心10?min。將上清液通過0.45?μm的膜過濾并儲存在4?℃待測。
以HHL作為ACE的反應底物。將0.25?units的ACE固體溶于2.5?mL的0.2?mol·L-1硼酸-硼砂緩沖溶液(0.2?mol·L-1NaCl,pH 8.3)配制成ACE溶液,稱取29.1?mg的HHL固體溶于15?mL的0.2?mol·L-1硼酸-硼砂緩沖溶液(0.2?mol·L-1NaCl,pH 8.3)配制成HHL溶液(5?mmol·L-1)。將20?μL的ACE溶液與50?μL不同質量濃度(0.5、1、2、4、8?mg·mL-1)的茶葉樣品分別混合,在37?℃水浴鍋中孵育10?min,隨后加入100?μL的HHL(5?mmol·L-1),漩渦搖勻后立即在37?℃下酶促反應60?min,最后加入300?μL的1.0?mol·L-1鹽酸終止反應。體系溶液采用0.45?μm水系濾膜過濾后進行HPLC檢測分析馬尿酸的含量。檢測波長為228?nm,柱溫為35?℃,進樣量為10?μL,流速為1?mL·min-1,流動相A為超純水(含0.1%三氟乙酸),B相為純乙腈,洗脫條件為75%的A相和25%的B相等度洗脫。為了避免樣品顏色可能產生的影響,以加入樣品但不加ACE溶液作為樣品空白組(表1)。
ACE抑制活性根據以下公式計算:
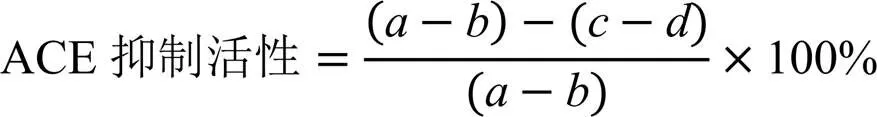
式中,為對照組馬尿酸的峰面積,為空白組馬尿酸的峰面積;為樣品組馬尿酸的峰面積;為樣品空白組馬尿酸的峰面積。
1.3.6 分子對接分析方法
為了在分子水平上模擬化合物和ACE之間的相互作用,使用AutodockVina 1.2.3(Scripps Research Institute,La Jolla,CA,USA)進行分子對接模擬[18]。從蛋白質數據庫(http://www.rcsb.org)獲得人類ACE-利辛普利復合物(蛋白質ID:1O86)的三維晶體結構,作為對接過程中的受體。茶黃素和兒茶素等對接配體的三維結構從美國國家醫學圖書館(https://pubchem.ncbi.nlm.nih.gov)獲得,使用ChemBio3DUltra 14.0軟件(Cambridge Soft,Cambridge,MA,USA)進行能量最小化。在對接之前,除去1O86中的所有雜分子和水,但保留鋅原子和甘氨酸,并添加極性氫。以鋅原子坐標為中心(x:43.821,y:38.240,z:46.712),對接網格大小為22.5?? × 22.5?? × 22.5??,網格間距為0.375??。配體和ACE模型之間的分子對接采用AutodockVina 1.2.3軟件進行。使用拉馬克遺傳算法,運行64次。
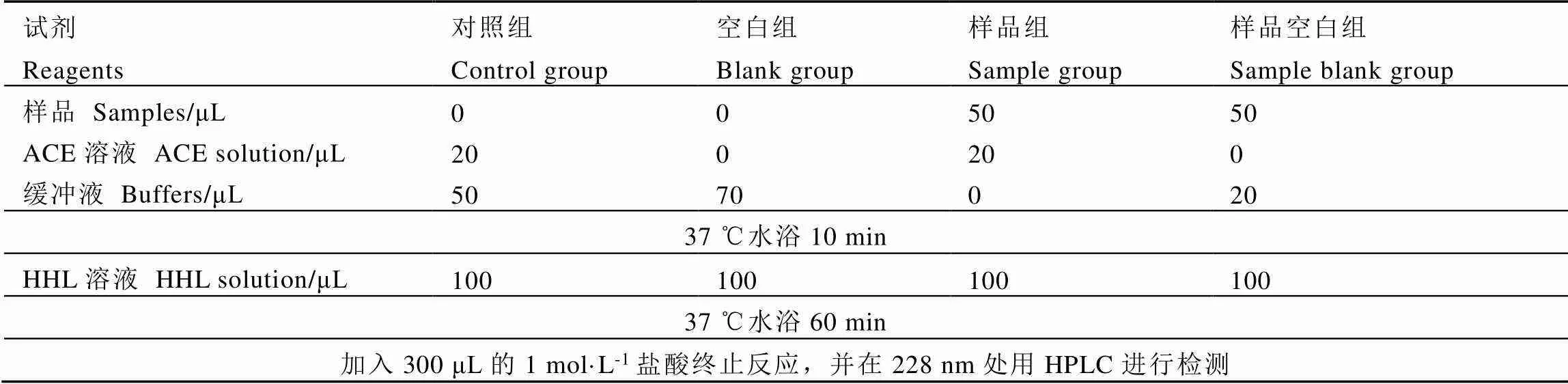
表1 體外ACE抑制率測定方法
通過比較化合物的對接姿勢中的結合能值,根據活性口袋中的最低結合能確定最佳構象,所有的對接都重復3次。配體和ACE模型之間的結合力類型通過在線網站(https://plip-tool.biotec.tu-dresden.de/plip-web/plip/index)分析。采用PyMOL 2.5.4軟件(DeLano Scientific LLC,CA,USA)分析蛋白質-配體之間的相互作用并可視化作圖。
1.4 數據處理
采用GraphPad Prism 9軟件(GraphPad Software,San Diego,USA)進行單因素方差分析(ANOVA)并計算IC50值,IC50值被定義為抑制一半ACE活性所需的抑制劑的濃度,通過ACE抑制率和抑制劑濃度的非線性曲線擬合計算。使用TBtools軟件進行熱圖分析。
2 結果與分析
2.1 厭氧處理后茶葉中氨基酸組成及含量分析
紫娟茶樹鮮葉經厭氧處理后采用不同加工工藝制備的茶葉樣品中氨基酸組成及含量如圖2所示。經厭氧處理后的凍干樣中GABA含量顯著增加至1.72?mg·g-1(<0.05),而未厭氧的常規凍干茶樣中僅為0.04?mg·g-1。不同加工工藝制備的綠茶、白茶和紅茶樣品中,綠茶樣的GABA含量最高(1.68?mg·g-1),其次為白茶樣(1.58?mg·g-1),紅茶樣最低(1.23?mg·g-1)。據報道,采用中茶108茶樹品種鮮葉經厭氧處理制備的綠茶和白茶的GABA含量分別為2.21、1.08?mg·g-1,而制備的紅茶的GABA含量最低,僅為0.86?mg·g-1[18],這與本研究結果基本一致,表明茶鮮葉厭氧處理后采用綠茶加工工藝,有望獲得富含GABA的特殊功能型茶產品。此外,茶葉中其他一些氨基酸組分也發生了顯著變化,例如谷氨酸含量在常規凍干樣(對照)中含量最高(0.83?mg·g-1),而在厭氧凍干樣中顯著下降至0.06?mg·g-1(<0.05),與厭氧凍干樣相比,厭氧處理后制備的綠茶、紅茶和白茶樣中含量均顯著增加,尤其是白茶樣中,其含量增加至0.52?mg·g-1(<0.05)。結果表明在厭氧處理結束后,谷氨酸等作為GABA合成前體的氨基酸能夠通過其他途徑進行補充,在天冬氨酸和谷氨酰胺也觀察到相似的變化規律。不同茶樣中氨基酸總量也存在顯著差異,常規凍干茶樣的顯著高于厭氧凍干茶樣(<0.05),鮮葉經厭氧處理后加工成成品茶,白茶中的最高(18.47?mg·g-1),而紅茶中的最低(11.46?mg·g-1),其原因是萎凋過程中蛋白質水解形成大量游離氨基酸[19],而紅茶在發酵過程中氨基酸成分轉化為揮發性成分導致含量顯著降低[20]。
2.2 厭氧處理后茶葉中多酚類及咖啡堿含量分析
不同加工工藝對紫娟茶樣中多酚類和咖啡堿含量的影響如表2所示。采用凍干處理能最大限度保留紫娟茶樣中類黃酮成分,而加工成綠茶、紅茶和白茶產品后,類黃酮成分含量呈現出不同程度下降。茶鮮葉經8?h厭氧處理后,凍干樣的EC、ECG及GCG含量均顯著高于常規凍干茶樣(<0.05)。通常,采用綠茶和白茶加工工藝能較好地保留茶鮮葉中茶多酚成分,而紅茶因發酵促進了茶多酚氧化形成聚合物如茶黃素類(主要包括TF、TF3G、TF3'G、TFDG 4種)和茶紅素類等,從而導致其含量顯著下降[19]。然而,相較于厭氧處理凍干茶樣,綠茶樣和白茶樣的絕大部分兒茶素組分以及茶多酚含量均顯著下降。在厭氧處理后,綠茶攤放和白茶萎凋過程觀察到部分葉片黃變或紅梗,結合綠茶樣和白茶樣中的茶黃素總量較高(分別為7.01?mg·g-1和6.67?mg·g-1),均顯著高于凍干樣中的茶黃素總量,推斷可能是由于茶鮮葉直接放入真空袋進行抽真空處理導致部分葉片出現機械損傷,因此在攤放和萎凋過程中葉片中部分茶多酚成分發生了酶促氧化生成茶黃素和茶紅素等茶色素類成分。其中,相較于厭氧凍干樣,綠茶和白茶的兒茶素總量分別下降了91.46?mg·g-1和98.14?mg·g-1,然而,茶黃素總量僅分別增加了4.83?mg·g-1和4.49?mg·g-1。值得注意的是,EGCG3''Me是紫娟茶樣的特殊功能成分,在厭氧凍干樣中含量為13.05?mg·g-1,在紅茶樣中僅為0.29?mg·g-1,這與以往的研究結果相似[21]。此外,3種成品茶中GA含量均顯著高于凍干茶樣,可能是由于在加工過程中,部分酯型兒茶素會水解成非酯型兒茶素并釋放GA,導致GA含量顯著增加[20]。此外,凍干處理能較好地保留紫娟茶樹鮮葉中花青素成分,厭氧凍干樣中其總量為0.95?mg·g-1,略高于常規凍干樣(0.91?mg·g-1),兩者差異不顯著,而與凍干茶樣相比,成品茶中花青素總量均顯著降低(<0.05),尤其是紅茶樣中最低,為0.10?mg·g-1。據報道[22-23],與紫娟茶樹鮮葉相比,加工成綠茶和紅茶后花青素含量分別下降了60.78%和92.90%,這與本研究結果相似,可能是由于加工過程中的高溫和發酵條件導致花青素成分降解。由此可見,為了更好地保持厭氧處理后茶樣中類黃酮成分,尤其是EGCG3''Me和花青素等特殊成分,應采用真空冷凍干燥進行加工。此外,常規凍干和厭氧凍干茶樣中咖啡堿含量分別為37.67?mg·g-1和37.55?mg·g-1,兩者之間無顯著差異,表明厭氧處理對茶樣中咖啡堿含量無顯著影響。相較于凍干茶樣,白茶樣中咖啡堿含量顯著增加至38.97?mg·g-1,而紅茶樣中則顯著下降至35.33?mg·g-1(<0.05),均存在顯著性差異。
現有的小區治理結構中存在著業主自治、居民自治和政府管理三種形式,但這些治理形式的邊界如何確定,不同物業類型的小區治理結構又有著何種差異,是困擾當前小區內部治理的關鍵性難題。
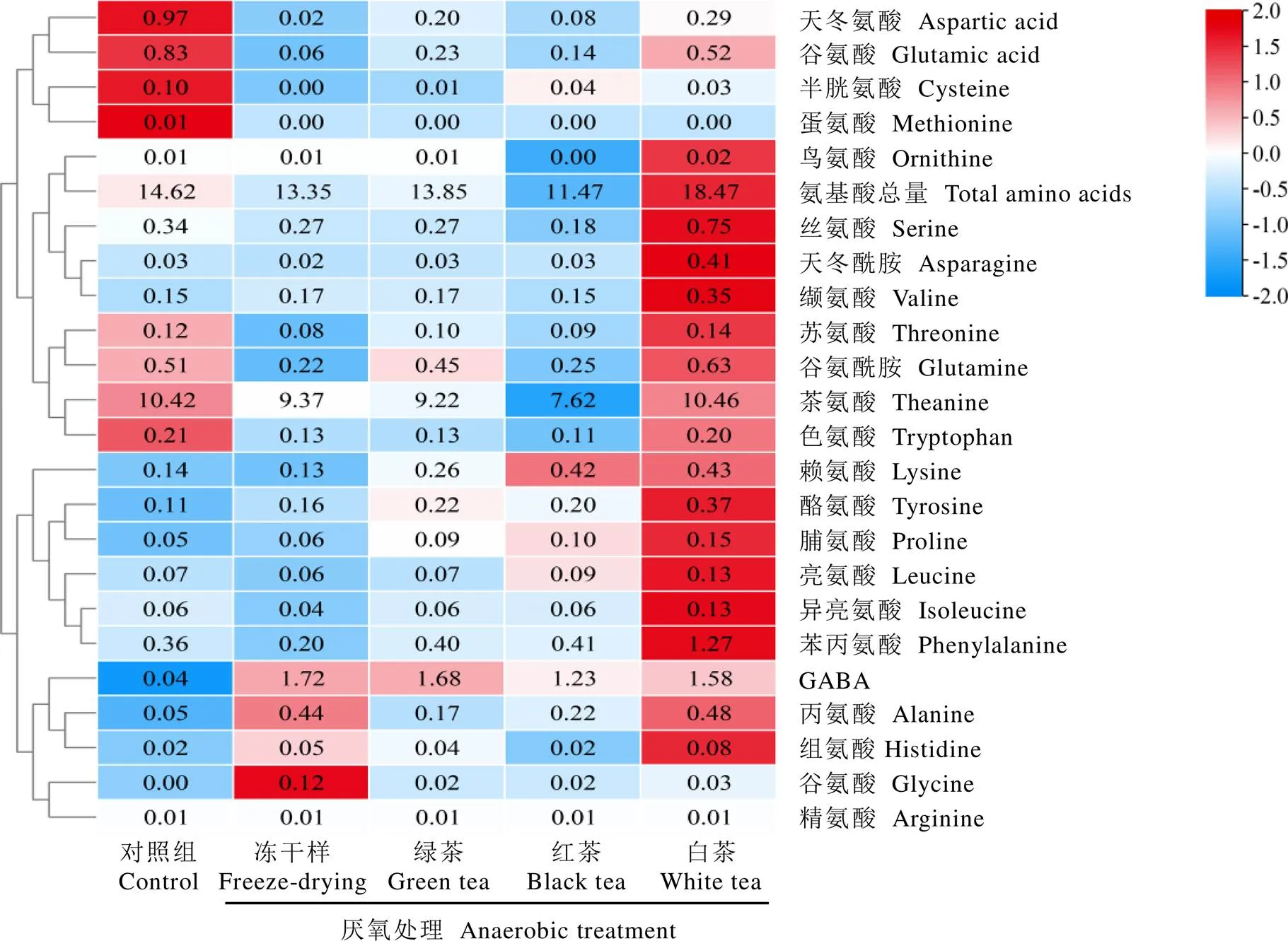
注:熱圖中的數值為茶樣中相應氨基酸的平均含量,單位為mg·g-1。對每行數值進行歸一化處理,熱圖的圖例通過顏色和數值的對應關系,將不同數值范圍映射到不同的顏色

表2 不同加工工藝紫娟茶樣中多酚類和咖啡堿含量
注:數據以平均值±標準差(SD)表示。同一行中的不同字母表示通過Tukey檢驗,各茶類之間的差異顯著(?0.05)
Note: The data are expressed as the mean ± standard deviation (SD). The different letters in the same row indicate that the difference between the teas is significant through Tukey test (?0.05)
2.3 不同類型茶葉對ACE的抑制活性分析
如圖3A所示,通過HPLC系統對HA和HHL混合標準品進行檢測,兩者能很好地分離。HA進樣質量濃度為0.012?5~0.400?0?mg·mL-1,HA峰面積(mAU·min)與濃度(mg·mL-1)的線性關系良好,回歸方程為=39?561?808+197?970,2=0.997?8(圖3B)。表明在該濃度范圍內,HA濃度與峰面積具有較好的線性關系,故樣品中的HA峰面積能反映樣品中HA的含量,進而反映ACE抑制活性。
IC50值被定義為抑制一半ACE活性所需抑制劑的濃度,數值越小表明抑制劑對ACE抑制活性越強。如圖4所示,常規凍干和厭氧凍干茶葉樣品的IC50值分別為3.247?mg·mL-1和2.608?mg·mL-1。厭氧凍干茶葉樣品的IC50值顯著低于常規凍干茶葉樣品的IC50值(<0.05),說明厭氧處理可以提高紫娟茶樹鮮葉樣品的ACE抑制活性。結合非揮發性化學成分的分析,推斷樣品的ACE抑制活性增強可能與厭氧處理后樣品中的GCG、ECG、EC和GABA等成分增加有關。此外,將該批次厭氧處理后的紫娟茶樹鮮葉按照綠茶、紅茶和白茶加工工藝制備成茶產品后,相較于厭氧凍干樣,其ACE抑制活性的IC50值(分別為3.603?mg·mL-1、5.420?mg·mL-1及3.457?mg·mL-1)均顯著增加(<0.05)。其中綠茶樣和白茶樣之間沒有顯著差異,但兩者均顯著低于紅茶樣。由此可知,茶鮮葉經厭氧處理后采用綠茶和白茶的加工工藝能夠更好地保持其對ACE的抑制活性。最近的研究表明,厭氧處理可以增加桑樹葉片中的GABA含量,并能提高桑葉提取物的體外ACE抑制活性[24],這支持了GABA可能具有作為ACE抑制劑的潛力。結合不同茶樣中氨基酸含量的分析結果(圖2),綠茶樣和白茶樣中的GABA含量均高于紅茶樣,因此,GABA可能是厭氧處理后茶樣中的重要抑制劑成分。常規凍干和厭氧白茶對ACE抑制活性無顯著差異,而常規凍干樣中GABA含量顯著低于厭氧白茶,表明茶樣中除了GABA還存在其他對ACE具有強抑制活性的成分。Dong等[6]發現,綠茶中的茶多酚能直接鈍化體外ACE的活性,顯示出較強的體外ACE抑制活性。楊高中等[18]研究報道,中茶108茶樹品種的鮮葉經厭氧處理后加工而成的不同類型茶葉樣品對ACE的抑制活性不同,其中,綠茶對ACE的抑制率較高,白茶次之,紅茶的抑制活性最弱,這與本研究結果基本一致。在本研究中,不同加工工藝制成的茶樣中的茶多酚含量差異顯著,其中,凍干樣中茶多酚含量顯著高于綠茶樣和白茶樣,紅茶樣中茶多酚含量最低,這與各個茶樣的ACE抑制活性強弱相對應,表明茶樣中的茶多酚可能是重要的ACE抑制劑。
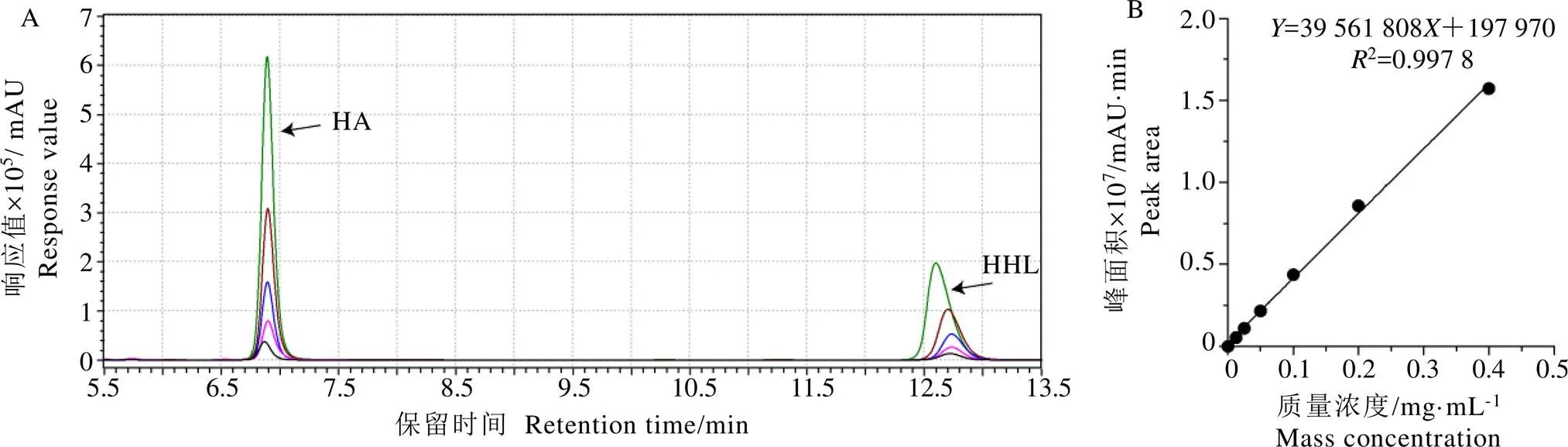
圖3 HHL和HA的色譜分離圖(A)以及HA的標準曲線圖(B)
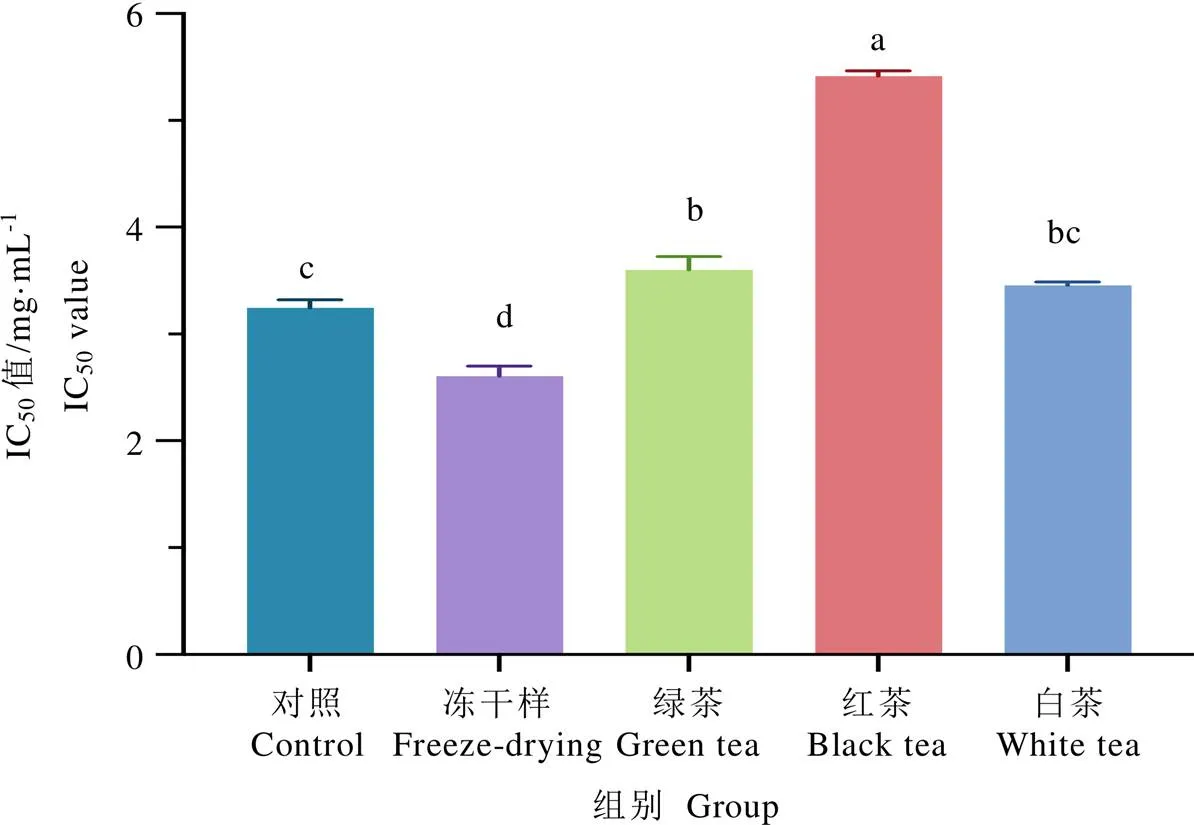
圖4 不同茶葉樣品的體外ACE抑制活性
茶葉中主要化學成分與其體外ACE抑制活性的IC50值的皮爾遜相關性分析見表3。相關性系數為正值時,表明該化合物含量越高,樣品體外ACE抑制活性的IC50值越大(即體外ACE抑制活性越弱);而相關性系數為負值時,表明該化合物含量越高,樣品體外ACE抑制活性的IC50值越小(即體外ACE抑制活性越強),可以推斷該化學成分可能是茶樣中主要的抑制劑成分。由表3可知,不同茶樣的體外ACE抑制活性的IC50值與茶樣中的花青素總量以及ECG的相關性系數均為–0.97,達到極顯著負相關水平(<0.01);與茶樣中的EGCG3"Me、茶多酚、GCG、兒茶素總量、EGCG等化學成分的相關系數在–0.95~–0.92,達到顯著負相關水平(<0.05)。因此,推斷這些成分可能是茶樣中發揮體外ACE抑制活性的主要化學成分。值得注意的是,GABA與茶樣的體外ACE抑制活性的IC50值的相關系數為–0.04,表明GABA對茶樣的體外ACE抑制活性貢獻不大。此外,茶黃素類成分和GA成分與茶樣的體外ACE抑制活性的IC50值均呈正相關,相關系數在0.61~0.84,但均未達到顯著水平(>0.05)。
注:采用皮爾遜相關性進行分析,相關性值用符號*表示,其中*表示<0.05,**表示<0.01
Note: The Pearson correlation was used for the analysis, and the correlationvalues are indicated by the symbol *, * indicates<0.05 and ** indicates<0.01
2.4 分子對接分析
茶葉中的主要化學成分與ACE模型分子對接的結合親和力結果如表4所示。結合親和力是指在結合過程中釋放的能量,其絕對值越大,結合越容易發生。因此,可以根據結合親和力的大小預測化合物對ACE的抑制活性強度[25]。分子對接結果顯示,茶黃素類化合物與ACE分子對接的結合親和力絕對值最大(10.49~11.37?kcal·mol-1),顯著高于其他成分(<0.05),表明茶黃素類可能是紅茶中發揮ACE抑制活性的重要成分。此外,飛燕草素-3--半乳糖苷和矢車菊素-3--半乳糖苷是紫娟茶樹鮮葉中含量最豐富的兩種花青素組分[5],它們與ACE分子對接的結合親和力絕對值分別為9.46、8.91?kcal·mol-1。EGCG3"Me也是紫娟茶樹鮮葉中的特殊功能成分,其與ACE的結合親和力絕對值為9.43?kcal·mol-1,高于未甲基化的EGCG(9.34?kcal·mol-1)和非酯型兒茶素EC(8.29?kcal·mol-1)。相比之下,GABA與ACE的結合親和力絕對值為3.91?kcal·mol-1,顯著小于其他化學成分(<0.05),表明GABA對體外ACE抑制活性可能較弱。然而,分子對接結果僅從理論層面揭示這些成分對ACE抑制活性的強弱,后續還要開展ACE抑制活性試驗進行驗證。
分子對接模擬表明,茶黃素類成分對ACE抑制活性可能強于花青素和兒茶素類成分。然而,2.3章節的相關性分析結果則表明茶黃素類與茶樣的體外ACE抑制活性的IC50值均呈相關(即表明茶黃素類并不是茶樣中抑制體外ACE活性的主要成分),兩者結果不一致。相較于厭氧凍干茶樣,盡管茶黃素類成分的含量在3種成品茶樣(綠茶、紅茶和白茶)中均顯著積累,但增加量相對較低(4.49~6.13?mg·g-1);然而3種成品茶樣中兒茶素總量急劇下降,這可能是導致成品茶樣的ACE抑制活性低于厭氧凍干茶樣的主要原因。因此,盡管茶黃素類成分對ACE抑制活性可能強于花青素和兒茶素類成分,但由于其在茶葉中的含量相對較低,其可能并非是茶葉中主要發揮體外ACE抑制活性的成分。值得注意的是,以往的研究報道GABA在體內具有降低動物和人類高血壓的健康功效,GABA一方面可以通過抑制ACE活性和減少血管緊張素Ⅱ的合成來降低血壓,另一方面可以作用于中樞神經系統,通過調節突觸前GABAB受體抑制去甲腎上腺素釋放或通過GABAA受體抑制節后交感心神經,從而引起血管擴張[11,26]。此外,GABA具有更高的口服生物利用度和穿越血腦屏障能力,這可能有助于GABA在體內發揮更有效的降壓活性[11]。然而,茶黃素類、兒茶素類(尤其是酯型兒茶素)和花青素類成分的生物利用度相對較低[27],因此,有必要對這些成分在體內的降血壓作用機制進一步研究。
TF3G、飛燕草素-3--半乳糖苷、EGCG、EC、GA和GABA等6個代表性化合物與ACE蛋白受體的分子對接的結合袋和相互作用分析如圖5所示。通過分子對接可以預測分子中的特定功能區域與目標蛋白質中結合位點之間的互作關系,對篩選天然生物活性抑制劑具有參考價值[28]。ACE的主要活性部位由3個活性口袋(S1、S2和S1')和Zn2+組成。S1口袋包含Ala354、Glu384和Tyr523,S2口袋包含Gln281、His353、Lys511、His513和Tyr520,而S1'口袋只包含Glu162[29-30]。分子對接結果表明,6種化合物都可以進入ACE的疏水口袋,形成穩定的疏水鍵。Shukor等[31]研究發現,一些官能團,如羥基和羧基,可能作為氫鍵受體或供體促進配體化合物與ACE之間的相互作用。然而,配體化合物與ACE之間相互作用的強弱也受到化合物空間結構等因素的影響。其中,TF3G主要與ACE的Ala354、Arg522、His353等殘基形成12個氫鍵,鍵距為2.3??~3.2??;還與Zn2+螯合形成1個鍵距為2.7??的氫鍵。飛燕草素-3--半乳糖苷與ACE的Tyr523、Glu384、Glu403和Glu411等殘基形成9個氫鍵,鍵距為2.8??~3.2??。此外,EGCG主要與ACE的Glu384、Glu411、Arg522和Tyr523等殘基形成8個氫鍵,鍵距為2.7??~3.1??。與EGCG相比,EC由于缺乏沒食子酰基,與ACE殘基His387、Glu411、Try523等形成6個氫鍵。酯型兒茶素水解后形成的GA則可以與ACE殘基Glu411和Gly404形成6個氫鍵,而且GA的苯環還與His387的咪唑環形成1個π堆積,鍵距為3.9??。GABA可與Tyr523和Glu411的ACE殘基形成3個氫鍵,且GABA可以深入隧道,直接與Zn2+螯合(鍵距為2.4??)。分子對接結果表明,6種成分主要與ACE的S1和S2口袋相互作用,而TF3G和GABA還能與ACE的Zn2+相互作用。茶黃素類與ACE的結合能力強于花青素類以及兒茶素類成分,可能與其分子結構中包含更多的親水基團和芳香環結構有關,使其能夠與ACE中的多個結構域發生相互作用。GABA與ACE的結合能力弱,可能是由于其分子量相對較小、結構較為簡單,導致其與ACE之間的相互作用較弱。
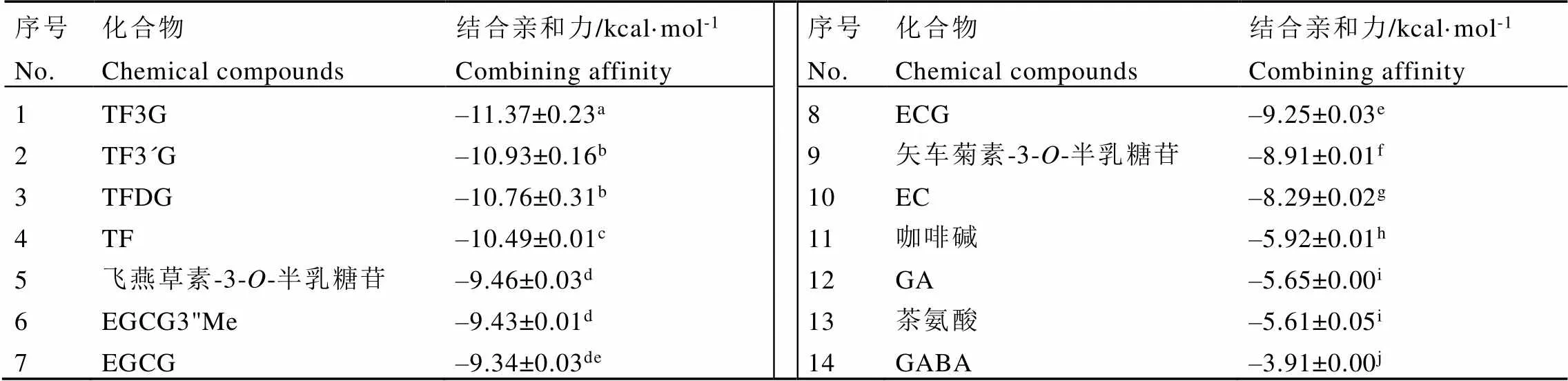
表4 茶葉中主要化學成分與ACE的分子對接的結合親和力
注:數據以平均值±標準差(SD)表示。同一列中的不同字母表示經Tukey多重比較檢驗,各化合物之間存在顯著差異(?0.05)
Note: The data are expressed as the mean ± standard deviation (SD). The different letters in the same column indicate a significant difference between the compounds by Tukey's multiple comparisons test (?0.05)

注:A為茶黃素-3-沒食子酸酯,B為飛燕草素-3-O-半乳糖苷,C為表沒食子兒茶素沒食子酸酯,D為表兒茶素,E為沒食子酸,F為γ-氨基丁酸。綠色虛線代表氫鍵,黑色虛線代表π堆疊
3 結論
本研究表明,厭氧處理能顯著提高紫娟茶樹鮮葉中GABA含量,而厭氧處理后不同的加工工藝不僅對茶樣中的氨基酸組成及含量影響較大,還顯著影響茶葉中多酚類化合物的組成和含量;采用凍干工藝能最大程度保留茶樣中多酚類化合物含量,其次為綠茶和白茶工藝,紅茶工藝保留茶樣中多酚類化合物含量最低。此外,厭氧處理能顯著提高紫娟茶的體外ACE抑制活性。紫娟茶樹鮮葉經厭氧處理后,采用凍干工藝制備的樣品體外ACE抑制活性最強,其次是采用綠茶和白茶工藝制備的樣品,而采用紅茶工藝制備的樣品ACE抑制活性最弱。相關性分析表明,茶葉中的茶多酚,尤其是兒茶素類成分可能是主要的體外ACE抑制活性成分,而與厭氧處理后GABA的增加無關。分子對接結果表明,茶黃素類成分與ACE的相互作用強于花青素類和兒茶素類成分,GABA最弱,這可能與其分子結構包含更多的親水基團和芳香環結構有關。后續有必要開展茶黃素類、兒茶素類和GABA等功能成分的體內試驗以明確其降血壓作用。研究結果有助于揭示厭氧處理對紫娟茶的體外ACE抑制活性的影響,為開發具有潛在治療高血壓的特殊功能型紫娟茶產品提供理論依據。
[1] Zhou B, Carrillo-larco R M, Danaei G, et al. Worldwide trends in hypertension prevalence and progress in treatment and control from 1990 to 2019: a pooled analysis of 1201 population-representative studies with 104 million participants [J]. The Lancet, 2021, 398(10304): 957-980.
[2] Li X C, Zhang J, Zhuo J L. The vasoprotective axes of the renin-angiotensin system: physiological relevance and therapeutic implications in cardiovascular, hypertensive and kidney diseases [J]. Pharmacological Research, 2017, 125: 21-38.
[3] Wu J, Liao W, Udenigwe C C. Revisiting the mechanisms of ACE inhibitory peptides from food proteins [J]. Trends in Food Science & Technology, 2017, 69: 214-219.
[4] Yang G Z, Meng Q, Shi J, et al. Special tea products featuring functional components: health benefits and processing strategies [J]. Comprehensive Reviews in Food Science and Food Safety, 2023, 22(3): 1686-1721.
[5] Shi J, Simal-gandara J, Mei J, et al. Insight into the pigmented anthocyanins and the major potential co-pigmented flavonoids in purple-coloured leaf teas [J]. Food Chemistry, 2021, 363: 130278. doi: 10.1016/j.foodchem.2021.130278.
[6] Dong J, Xu X, Liang Y, et al. Inhibition of angiotensin converting enzyme (ACE) activity by polyphenols from tea () and links to processing method [J]. Food & Function, 2011, 2(6): 310. doi: 10.1039/c1fo10023h.
[7] Kurita I, Maeda-yamamoto M, Tachibana H, et al. Antihypertensive effect of Benifuuki tea containing-methylated EGCG [J]. Journal of Agricultural and Food Chemistry, 2010, 58(3): 1903-1908.
[8] Fan W D, Zong H R, Zhao T, et al. Bioactivities and mechanisms of dietary proanthocyanidins on blood pressure lowering: a critical review ofand clinical studies [J]. Critical Reviews in Food Science and Nutrition, 2022: 1-17. doi: 10.1080/10408398.2022.2132375.
[9] Ockermann P, Headley L, Lizio R, et al. A review of the properties of anthocyanins and their influence on factors affecting cardiometabolic and cognitive health [J]. Nutrients, 2021, 13(8): 2831. doi: 10.3390/nu13082831.
[10] 包云秀, 夏麗飛, 李友勇, 等. 茶樹新品種‘紫娟’[J]. 園藝學報, 2008, 35(6): 934. Bao Y X, Xia L F, Li Y Y, et al. A new tea tree cultivar ‘Zjuan’ [J]. Acta Horticulturae Sinica, 2008, 35(6): 934.
[11] Hinton T, Johnston G A R. GABA-enriched teas as neuro-nutraceuticals [J]. Neurochemistry International, 2020, 141: 104895. doi: 10.1016/j.neuint.2020.104895.
[12] Dai W D, Xie D C, Lin Z, et al. A nontargeted and targeted metabolomics study on the dynamic changes in metabolite levels during the anaerobic treatment of-aminobutyric acid (GABA) tea [J]. LWT, 2020, 126: 109313. doi: 10.1016/j.lwt.2020.109313.
[13] Chen Q, Zhang Y M, Tao M M, et al. Comparative metabolic responses and adaptive strategies of tea leaves () to N2and CO2anaerobic treatment by a nontargeted metabolomics approach [J]. Journal of Agricultural and Food Chemistry, 2018, 66(36): 9565-9572.
[14] Yin Z T, Yan R Y, Jiang Y S, et al. Identification of peptides in Qingke baijiu and evaluation of its angiotensin converting enzyme (ACE) inhibitory activity and stability [J]. Food Chemistry, 2022, 395: 133551. doi: 10.1016/j.foodchem.2022.133551.
[15] Wei D, Fan W L, Xu Y. Identification of water-soluble peptides in distilled spent grain and its angiotensin converting enzyme (ACE) inhibitory activity based on UPLC-Q-TOF-MS and proteomics analysis [J]. Food Chemistry, 2021, 353: 129521. doi: 10.1016/j.foodchem.2021.129521.
[16] Yang C, Hu Z Y, Lu M L, et al. Application of metabolomics profiling in the analysis of metabolites and taste quality in different subtypes of white tea [J]. Food Research International, 2018, 106: 909-919.
[17] Da Silva T B V, Castilho P A, De Sá-nakanishi A B, et al. The inhibitory action of purple tea onstarch digestion compared to otherteas [J]. Food Research International, 2021, 150: 110781. doi: 10.1016/j.foodres.2021.110781.
[18] 楊高中, 彭群華, 張悅, 等. 厭氧處理對不同類型茶葉的氨基酸組成及生物活性的影響[J]. 茶葉科學, 2022, 42(2): 222-232. Yang G Z, Peng Q H, Zhang Y, et al. Effects of anaerobic treatment on amino acid composition and biological activities of different type teas [J]. Journal of Tea Science, 2022, 42(2): 222-232.
[19] Wang Y, Kan Z, Thompson H J, et al. Impact of six typical processing methods on the chemical composition of tea leaves using a singlecultivar, Longjing 43 [J]. Journal of Agricultural and Food Chemistry, 2019, 67(19): 5423-5436.
[20] Yu F, Chen C, Chen S N, et al. Dynamic changes and mechanisms of organic acids during black tea manufacturing process [J]. Food Control, 2022, 132: 108535. doi: 10.1016/j.foodcont.2021.108535.
[21] 呂海鵬, 楊停, 梁名志, 等. “紫娟”茶中的EGCG3"Me成分研究[J]. 現代食品科技, 2014, 30(9): 286-289, 296. Lü H P, Yang T, Liang M Z, et al. Study of EGCG3"Me content in Zijuan tea [J]. Modern Food Science and Technology, 2014, 30(9): 286-289, 296.
[22] 時鴻迪, 王邦政, 李乾, 等. 不同加工工藝下‘紫娟’茶品質的比較[J]. 中國農學通報, 2020, 36(34): 142-147. Shi H D, Wang B Z, Li Q, et al. The quality of ‘Zijuan’ tea under different processing techniques: a comparative analysis [J]. Chinese Agricultural Science Bulletin, 2020, 36(34): 142-147.
[23] Lü H P, Dai W D, Tan J F, et al. Identification of the anthocyanins from the purple leaf coloured tea cultivar Zijuan (var.) and characterization of their antioxidant activities [J]. Journal of Functional Foods, 2015, 17: 449-458.
[24] Tu J, Liu G H, Jin Y C, et al. Enrichment of-aminobutyric acid in mulberry leaves and the inhibitory effects of the water extract on ACE and-glucosidase activity [J]. Industrial Crops and Products, 2022, 177: 114485. doi: 10.1016/j.indcrop.2021.114485.
[25] Wu Y, Han Z S, Wen M C, et al. Screening of-glucosidase inhibitors in large-leaf yellow tea by offline bioassay coupled with liquid chromatography tandem mass spectrometry [J]. Food Science and Human Wellness, 2022, 11(3): 627-634.
[26] Diana M, Quílez J, Rafecas M. Gamma-aminobutyric acid as a bioactive compound in foods: a review [J]. Journal of Functional Foods, 2014, 10: 407-420.
[27] Liu Z, Bruins M E, Ni L, et al. Green and black tea phenolics: bioavailability, transformation by colonic microbiota, and modulation of colonic microbiota [J]. Journal of Agricultural and Food Chemistry, 2018, 66(32): 8469-8477.
[28] Wang X M, Chen H X, Fu X G, et al. A novel antioxidant and ACE inhibitory peptide from rice bran protein: biochemical characterization and molecular docking study [J]. LWT, 2017, 75: 93-99. doi: 10.1016/j.lwt.2016.08.047.
[29] Shih Y H, Chen F A, Wang L F, et al. Discovery and study of novel antihypertensive peptides derived from cassia obtusifoliaseeds [J]. Journal of Agricultural and Food Chemistry, 2019, 67(28): 7810-7820.
[30] Mirzaei M, Mirdamadi S, Ehsani M R, et al. Production of antioxidant and ACE-inhibitory peptides fromprotein hydrolysates: purification and molecular docking [J]. Journal of Food and Drug Analysis, 2018, 26(2): 696-705.
[31] Shukor N A, Van Camp J, Gonzales G B, et al. Angiotensin-converting enzyme inhibitory effects by plant phenolic compounds: a study of structure activity relationships [J]. Journal of Agricultural and Food Chemistry, 2013, 61(48): 11832-11839. doi:10.1021/jf404641v.
Changes in Chemical Composition of ZijuanTea under Anaerobic Treatment Conditions and Their Effects onACEActivity
YANG Gaozhong1,2, SHI Jiang1, ZHANG Yue1, PENG Qunhua1, LIN Zhi1*, Lü Haipeng1*
1. Tea Research Institute of the Chinese Academy of Agricultural Science/Key Laboratory of Biology, Genetics and Breeding of SpecialEconomic Animals and Plants, Ministry of Agriculture and Rural Affairs, Hangzhou 310008, China; 2. Graduate School of Chinese Academy of Agricultural Sciences, Beijing 100081, China
In this study, fresh leaves of Zijuan tea were treated with anaerobic treatment and used to make freeze-dried, green, black and white tea samples. The main non-volatile components (such as catechins, amino acids, anthocyanins) in these samples were determined and theirangiotensin-converting enzyme (ACE) inhibitory activities were evaluated by high performance liquid chromatography. In addition, molecular docking simulation experiments were conducted to investigate the potential interaction mechanisms between the major chemical components in tea and ACE. The results show that the GABA content in the freeze-dried samples after anaerobic treatment increased to 1.72?mg·g-1, which was significantly higher than that in the tea samples without anaerobic treatment (0.04?mg·g-1,<0.05). The composition and contents of amino acids and polyphenolic compounds in the tea samples were significantly affected by the processing methods after anaerobic treatment. The white tea process could increase the contents of amino acid components in the tea samples, while the freeze-drying process could maximize the retention of polyphenolic compounds in the tea samples. TheACE inhibitory activity of Zijuan tea fresh leaf samples was significantly enhanced after anaerobic treatment (<0.05); the strongest ACE activity inhibition was observed in samples prepared by freeze-drying process after anaerobic treatment, which was significantly higher than that of samples prepared by green, white, and black tea processing methods (<0.05). In addition, molecular docking simulation experiments reveal that the strength of the interaction between the main chemical components in tea and ACE was related to the hydrophilic groups and aromatic ring structures in their molecular structures. The results of the study helped to reveal the effect of anaerobic treatment on the ACE inhibitory activity of Zijuan tea and provided a theoretical basis for the development of special functional Zijuan tea products with potential treatment of hypertension.
tea,-aminobutyric acid, anaerobic treatment, processing technology, angiotensin-converting enzyme
S571.1
A
1000-369X(2023)04-553-14
2023-04-17
2023-05-14
國家茶葉產業技術體系(CARS-19)、中國農業科學院科技創新工程項目(CAAS-ASTIP-2014-TRICAAS)
楊高中,男,碩士研究生,主要從事茶葉品質化學與營養健康方面研究。*通信作者:linz@tricaas.com;lvhaipeng@tricaas.com
