鳩坑龍井茶對高脂飲食C57BL/6小鼠肝脂肪變性SREBPs通路信號的影響及腸道菌群調節作用研究
2023-08-30 14:19:36龔明秀袁懿煒張一帆葉江成郭麗李曉軍黃皓毛宇驍趙蕓趙進
茶葉科學 2023年4期
關鍵詞:小鼠
龔明秀,袁懿煒,張一帆,葉江成,郭麗,李曉軍,黃皓,毛宇驍,趙蕓,趙進*
鳩坑龍井茶對高脂飲食C57BL/6小鼠肝脂肪變性SREBPs通路信號的影響及腸道菌群調節作用研究
龔明秀1,2,袁懿煒1,2,張一帆1,2,葉江成1,2,郭麗3,李曉軍4,黃皓4,毛宇驍5,趙蕓5,趙進1,2*
1. 中國計量大學生命科學學院,食品營養與質量安全研究所,浙江 杭州 310018;2. 特色農產品品質與危害物控制技術浙江省重點實驗室,浙江 杭州 310018;3. 中國農業科學院茶葉研究所,浙江 杭州 310008;4. 浙江藝福堂茶業有限公司博士創新工作站,浙江 杭州 311500;5. 杭州市農業科學研究院,浙江 杭州 310024
探究鳩坑龍井茶水提物(LJT)對小鼠肝組織脂質代謝SREBPs通路信號影響及腸道菌群的調節作用。通過高脂飲食誘導小鼠構建非酒精性脂肪肝(NAFL)模型,并給予LJT(300?mg·kg-1)灌胃干預。定期記錄小鼠的體質量,檢測小鼠血清生化指標和葡萄糖耐受水平,觀察并分析Hematoxylin-Eosin(HE)染色、油紅O染色肝組織切片特征;應用Real-time qPCR技術檢測小鼠肝組織SREBPs通路7個基因(-、、-、-、-、、)的相對表達量,采用蛋白免疫印記技術(Western Blot)分析肝組織蛋白質表達水平,同時對小鼠腸道菌群進行高通量測序(16?S rDNA)并分析其結構。結果顯示,LJT干預后小鼠體質量、血糖AUC、血清TG、TC、LDL-C和肝臟中TG、TC水平有顯著下降,龍井組小鼠肝組織SREBP-1c、FAS、ACC-1、SCD-1和PPAR蛋白表達水平降低,-、-、、-、-、、基因的相對表達量顯著下調;16?S rDNA分析發現,小鼠腸道菌群門水平主要為Firmicutes、Bacteroidota、Desulfobacterota和Actinobacteriota4類,LJT有效延緩了高脂飲食引起的Firmicutes相對豐度升高和Bacteroidota相對豐度下降趨勢,并增加了腸道菌群的物種豐度。結果表明,LJT能夠干預小鼠肝脂肪變性SREBPs通路信號表達,改善小鼠腸道菌群紊亂,具有降脂減肥作用。
鳩坑龍井茶;非酒精性脂肪肝;SREBPs通路;腸道菌群紊亂;降脂作用
非酒精性脂肪肝病(Non-alcoholic fatty liver disease,NAFLD)是指除酒精、藥物和單基因遺傳等肝損傷原因外,導致的肝細胞內脂質積聚引起的綜合征,肝細胞內甘油三酯的積聚(即肝脂肪變性)是NAFLD的標志[1]。其發展過程包括從非酒精性脂肪肝(Non-alcoholic fatty liver,NAFL)到非酒精性脂肪性肝炎(Non-alcoholic steato-hepatitis,NASH),甚至是晚期的肝纖維化和肝硬化[2]。此癥狀與心血管疾病[3]、肥胖[4]、Ⅱ型糖尿病[5]和肝外惡性腫瘤[6]等代謝疾病密切相關。報道顯示,NAFLD成為我國發病率最高的慢性肝病,在部分地區達25%[7]。腸道菌群與NAFLD密切相關,可以通過激活固醇調節元件結合蛋白(Sterol regulatory element binding proteins,SREBPs)來影響宿主脂質代謝[8]。SREBPs是調控膽固醇、甘油三酯和脂肪酸從頭合成的關鍵核轉錄因子,通過調節SREBPs信號通路抑制肝臟中脂質沉積,緩解NAFLD癥狀。
《地理標志產品龍井茶》(GB/T 18650—2008)指出,鳩坑種是適宜加工龍井茶的茶樹良種,其芽葉肥壯,持嫩性強,內含物質豐富,具有相對豐富的香氣成分[9-10]。茶多酚具有良好的降脂作用,且降脂功效的強弱與其濃度呈正相關[11]。Li等[12]研究發現龍井茶能預防酒精引起的肝臟脂質的累積,Ma等[13]研究顯示綠茶通過調節SREBPs信號通路抑制肝臟中脂質沉積。本研究通過高脂飲食誘導C57BL/6小鼠構建非酒精性肝脂肪變性模型,研究鳩坑龍井茶水提物對小鼠肝組織脂質代謝SREBPs通路信號的影響及腸道菌群的調節作用,進而解析鳩坑龍井茶改善小鼠NAFLD功效的作用機理,旨在為促進鳩坑龍井茶產品的功效應用提供理論依據。
1 材料與方法
1.1 試驗材料
2021年11月19日采摘桐廬茶園的鳩坑種茶樹鮮葉,按照龍井茶工藝加工制作龍井茶。
8周齡雄性SPF級C57BL/6小鼠購自杭州子源實驗動物科技有限公司,使用許可證號為SYXK(浙)2018-0009。
小鼠普通飼料(蛋白質20.6%、脂肪12%、碳水化合物67.4%)購自江蘇省協同醫藥生物工程有限責任公司,高脂飼料(蛋白質26.2%、脂肪34.9%、碳水化合物26.3%)購自無錫帆泊生物技術有限公司。
1.2 試驗方法
1.2.1 指標測定
水分含量參照GB 5009.3—2016直接干燥法,水浸出物含量參照GB/T 8305—2013方法,咖啡堿含量參照GB/T 8312—2013紫外分光光度法,茶多酚及兒茶素含量參照GB/T 8313—2018方法,游離氨基酸含量參照GB/T 8314—2013方法,可溶性糖含量采用苯酚-硫酸法[14],可溶性蛋白含量測定采用微孔酶標法。
1.2.2 動物實驗
動物實驗經中國計量大學動物倫理委員會批準(批件號:2022年第28號)。40只體質量為(23.00±1.00)?g的C57BL/6雄性小鼠,安置在常規環境(12?h光/暗循環)中,自由取食飲水,適應性飼養1周后進行隨機分組,分為正常組(Normal)、高脂組(Model)、陽性對照組(簡稱陽性組,Control)和鳩坑龍井茶水提物干預組(簡稱龍井組,LJT),小鼠分組方案如表1所示,小鼠每天進行灌胃。鳩坑龍井茶水提物的制備參照Li等[15]的方法并進行完善,用95?℃熱水按料液比1∶20浸提茶制品5?min,趁熱抽濾、旋蒸得濃縮濾液,然后–55?℃冷凍干燥24?h得到茶水提取物凍干粉,用于配置灌胃所需茶湯。每周測定小鼠體質量,在第9周飼養結束后解剖小鼠,觀察并記錄小鼠、肝臟和附睪脂肪形態,按照下列公式計算Lee’s指數:
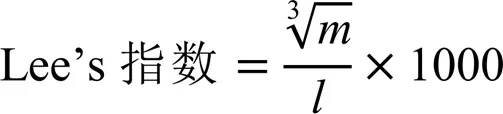
式中,為小鼠體質量,g;為小鼠體長,cm。
1.2.3 口服葡萄糖耐受量實驗(OGTT)
在小鼠飼喂第5周,禁食不禁水12?h,測定各組小鼠空腹血糖值。小鼠灌胃20%葡萄糖水溶液(2?g·kg-1),在小鼠空腹和灌胃后第15、30、60、120?min,分別用羅氏血糖儀和檢測試紙條依次測定相應時間點的小鼠血糖值,計算時間-血糖曲線下面積(AUC)。
1.2.4 小鼠血清采集
在小鼠培養第9周,禁食不禁水過夜(12?h)。禁食結束后,采用眼球取血方式采集小鼠鮮血于抗凝管中靜置2?h,然后以3?000?r·min-1冷凍離心(4?℃)10?min,收集上層血清并及時儲存于–80?℃冰柜。
1.2.5 血清和肝臟常規指標測定
血清和肝臟中高密度脂蛋白膽固醇(High-density lipoprotein,HDL-C)、低密度脂蛋白膽固醇(Low-density lipoprotein,LDL-C)、總膽固醇(Total cholesterol,TC)、總甘油三酯(Total triglyceride,TG)和總蛋白(Total protein,TP)等含量,谷草轉氨酶(Aspartate aminotransferase,AST)和谷丙轉氨酶(Alanine aminotransferase,ALT)的活性采用南京建成生物工程研究所試劑盒測定。
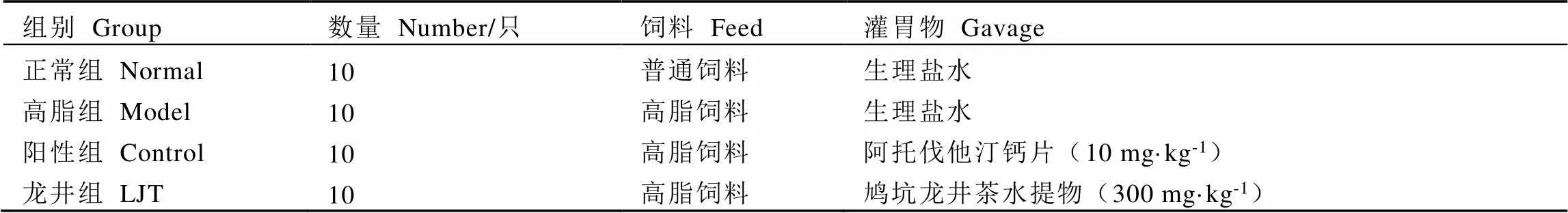
表1 小鼠分組方案
1.2.6 肝臟組織染色
采集小鼠肝組織并置于10%福爾馬林組織固定液中48?h,參考馮琳等[16]方法并對其進行油紅O染色和HE染色切片制作。
1.2.7 蛋白免疫印記
小鼠肝臟樣品液氮保存送至杭州紐貝生物有限公司,采用蛋白質印跡技術(Western blot)分析小鼠肝組織脂質代謝相關蛋白質表達情況。通過SuperSignal?West Dura Extended Duration Substrate進行顯影和定影,使用Image J 1.8.0圖像處理軟件(美國)分析條帶的光密度值,每個條帶重復3次,目的蛋白相對表達量=[目的蛋白(光密度值)/內參(光密度值)]×10n。
1.2.8 實時熒光定量PCR
1.2.9 腸道菌群檢測
采集小鼠結腸部位內容物,提取樣品總DNA后,根據保守區設計得到引物,在引物末端加上測序接頭,進行PCR擴增并對其產物進行純化、定量和均一化形成測序文庫并進行文庫質檢,質檢合格的文庫用Illumina NovaSeq 6000進行測序。測序由北京擎科生物公司完成。
1.3 數據分析
2 結果與分析
2.1 鳩坑龍井茶主要理化成分檢測結果
鳩坑龍井茶主要理化成分檢測結果如表3所示。鳩坑龍井茶中水浸出物含量為(31.590±0.292)%,茶多酚含量為(19.773±0.284)%,可溶性糖含量為(4.656±0.070)%,可溶性蛋白含量為(4.103±0.026)%,咖啡堿含量為(2.455±0.012)%,水分含量為(5.173±0.132)%。
2.2 鳩坑龍井茶對高脂飲食小鼠臟器、體質量和Lee’s指數的影響
鳩坑龍井茶對小鼠形體的影響如圖1所示。由圖1A可知,經過9周的高脂飲食誘導性飼養,高脂組、陽性組和龍井組小鼠的體型明顯比正常組肥大,其中龍井組小鼠體型比高脂組瘦小;正常組小鼠肝臟呈現為健康的深紅褐色,高脂組肝臟顏色偏黃,有明顯脂肪聚積現象,陽性組和龍井組小鼠肝組織形態與正常組相近;高脂組小鼠附睪脂肪體積與其他3組有明顯差異。

表2 熒光定量PCR引物序列
小鼠飼喂期間體質量和體形變化情況如圖1B和圖1C所示。高脂組小鼠體質量和Lee’s指數顯著高于正常組(<0.05),陽性組和龍井組小鼠體質量顯著低于高脂組(<0.05)。研究結果顯示,高脂飲食能夠誘導小鼠肥胖與小鼠肝組織脂肪堆積,并導致肝脂肪變性,而通過長期飲食鳩坑龍井茶能減緩或者改善小鼠肝脂肪變性癥狀。
2.3 小鼠血糖耐受水平檢測分析
小鼠血糖變化如圖2所示。正常組小鼠血糖值在口服葡萄糖溶液15?min后達到最高水平,120?min后下降至接近正常水平,并且正常組小鼠在口服糖期間AUC值顯著低于高脂組(<0.01);高脂組、陽性組和龍井組小鼠血糖在口服葡萄糖后30?min達到最高水平,120?min后高脂組血糖水平仍高于正常組,而陽性組、龍井組小鼠血糖值趨近于正常組水平,并顯著低于高脂組(<0.05)。

表3 鳩坑龍井茶中主要理化成分含量
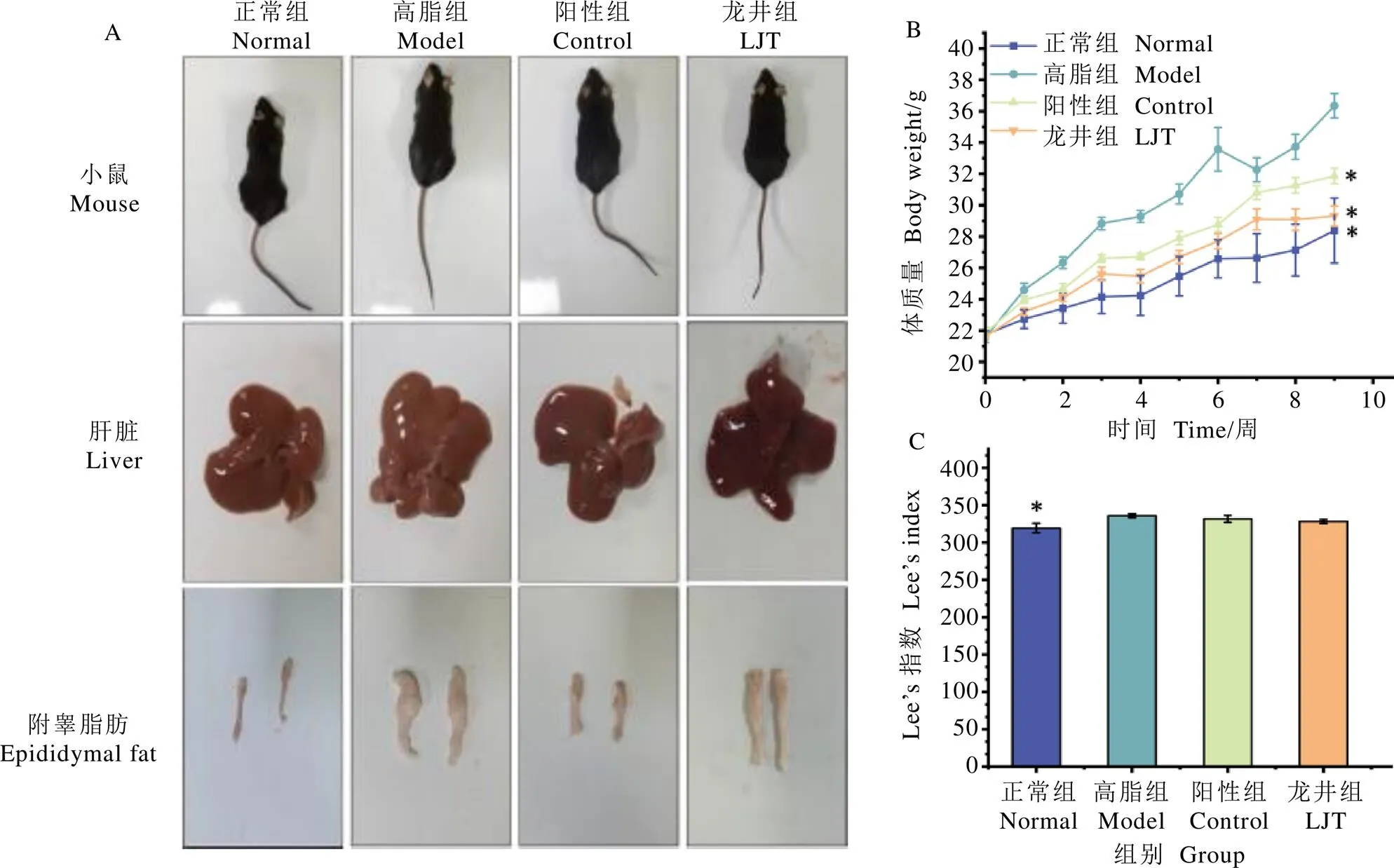
注:A為小鼠體型、肝臟和附睪脂肪,B為0~9周小鼠體質量變化,C為小鼠Lee’s指數。*表示與高脂組相比P<0.05
由圖2B可知,與高脂組小鼠相比,正常組、陽性組和龍井組的小鼠血糖AUC均顯著下降(<0.05)。檢測結果表明,高脂飲食能引起C57BL/6小鼠葡萄糖耐受能力受損,通過長期灌胃龍井茶水提物能有效改善小鼠葡萄糖耐受性。
2.4 小鼠血清生化指標檢測分析
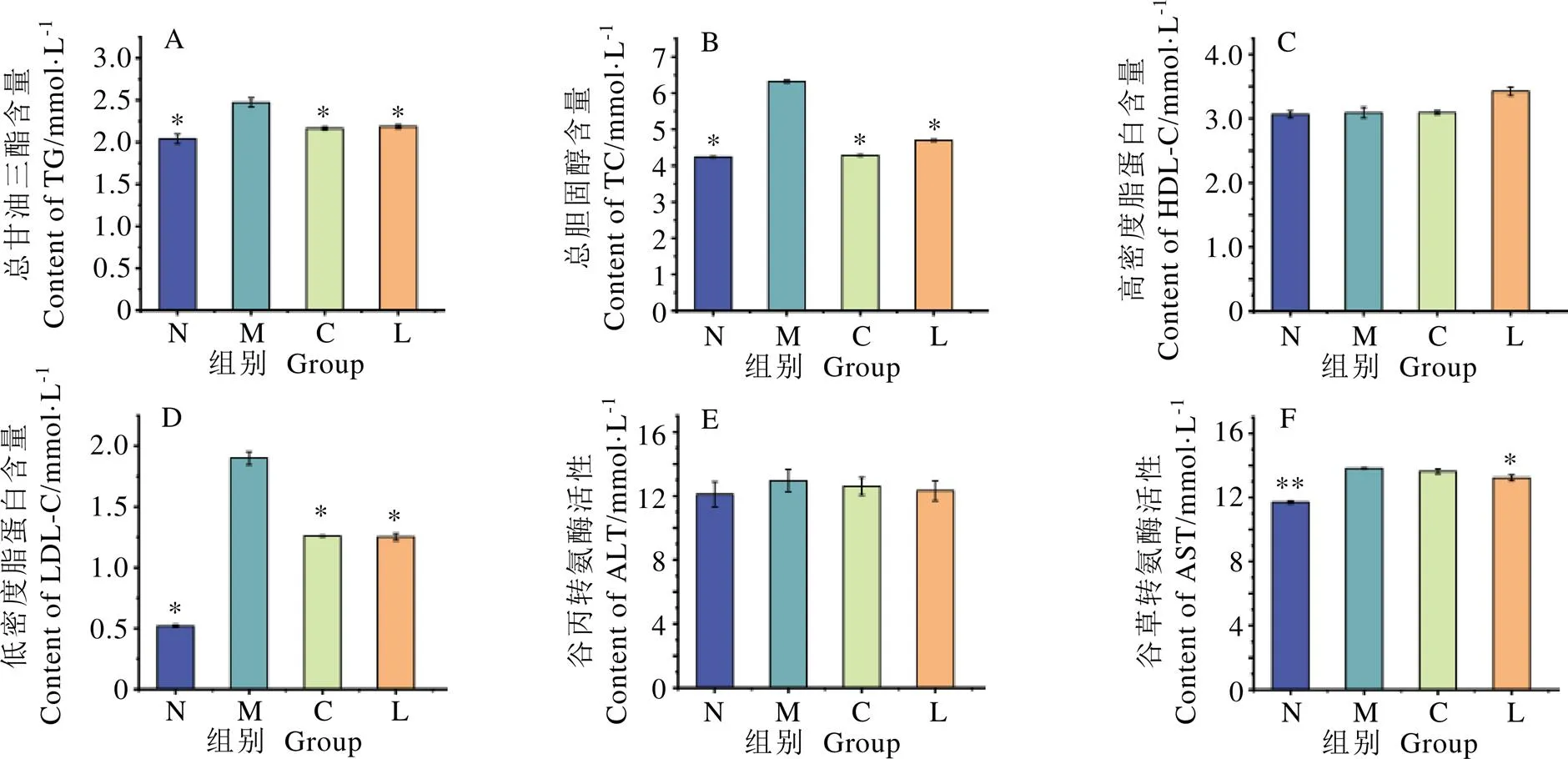
小鼠血清生化指標變化趨勢如圖3。由圖3A~D可知,與高脂組相比,正常組陽性組和龍井組小鼠的TC、TG和LDL-C值顯著降低(<0.05),各組間的HDL-C無顯著差異。小鼠血清中酶活性水平如圖3E和3F所示,高脂組小鼠AST酶活性水平顯著高于正常組(<0.01),但仍處于正常水平范圍內,龍井組小鼠血清AST酶活性水平顯著低于高脂組(<0.05);小鼠血清ALT酶活性在各組間不具有統計學差異。結果表明,鳩坑龍井茶能有效降低高脂飲食引起的小鼠血脂含量,推測長期飲用鳩坑龍井茶能夠降低血脂累積水平。
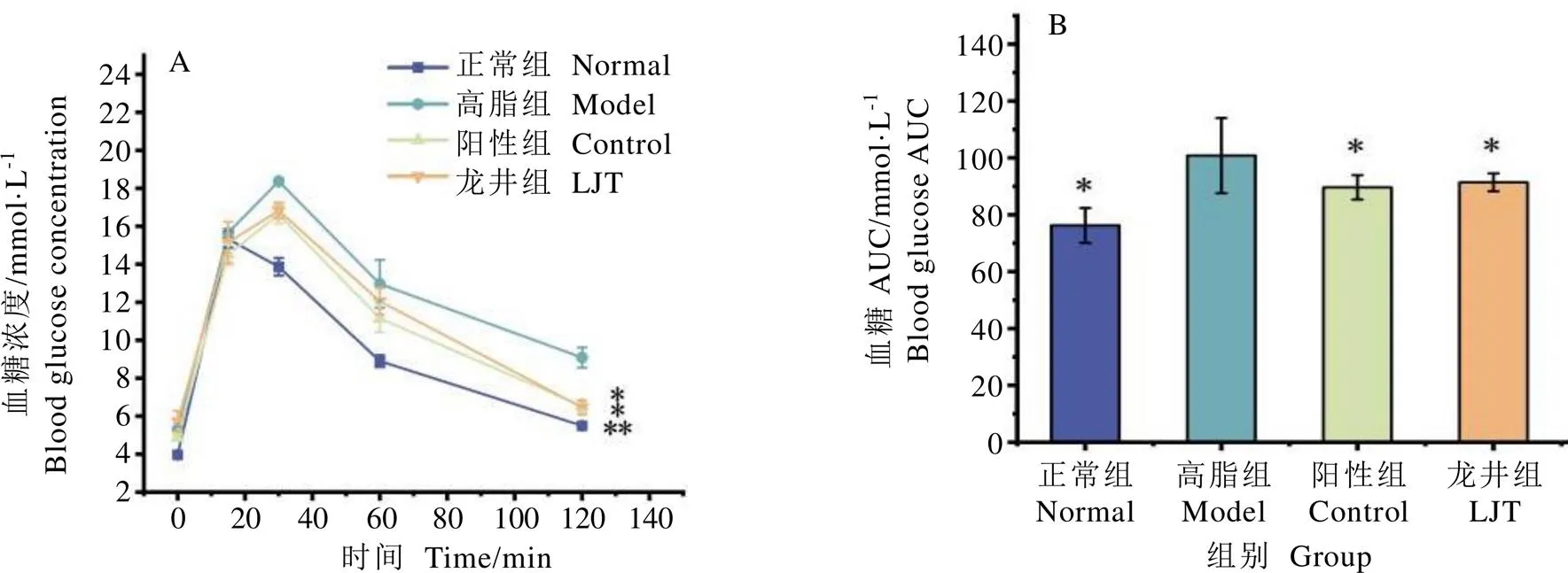
注:A為小鼠OGTT血糖變化曲線,B為小鼠OGTT曲線下面積。*表示與高脂組相比P<0.05,**表示與高脂組相比P<0.01,下同
注:N為正常組,M為高脂組,C為陽性組,L為龍井組
2.5 小鼠肝組織脂肪變性檢測分析
小鼠肝組織脂質含量如圖4。由圖4A和4B可以得出,高脂飲食能夠顯著增加小鼠肝臟TG和TC的含量(<0.05),陽性藥物和鳩坑龍井茶具有顯著降低小鼠肝臟TG和TC的作用(<0.05)。
HE染色結果顯示(圖4C),正常組的小鼠肝細胞排列緊密,肝小葉結構清晰,脂肪空泡密度小;高脂組小鼠肝細胞排列較松散,肝小葉結構不清晰,大量的脂肪空泡聚集在細胞核周圍,少量炎性細胞浸潤現象,但無氣球樣病變;陽性組和龍井組小鼠肝細胞結構中脂肪空泡減少,炎性細胞浸潤現象幾乎消失。油紅O染色結果顯示(4D),正常組小鼠肝組織中甘油三酯的含量較少,高脂組小鼠肝組織甘油三酯遍布大部分或整個細胞質,且含量顯著高于正常組(<0.05),陽性組和龍井組小鼠肝組織甘油三酯含量顯著低于高脂組(<0.05)。肝組織切片病理觀察結果表明,高脂飲食誘發小鼠肝組織脂肪堆積并發生變性,而長期飲食鳩坑龍井茶能有效緩解該癥狀。
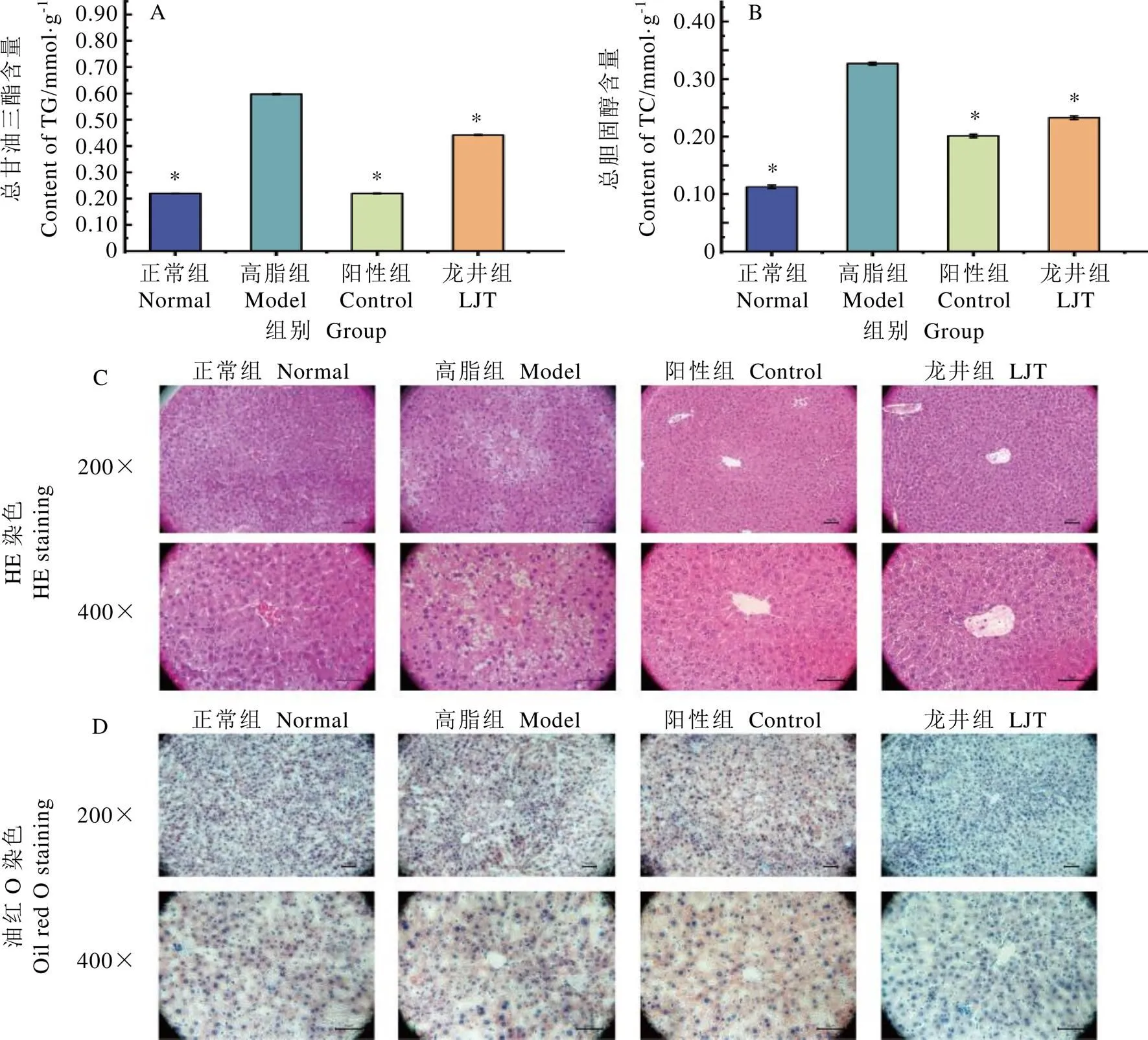
圖4 小鼠肝組織脂質含量
2.6 鳩坑龍井茶對肝組織脂質代謝通路信號的影響
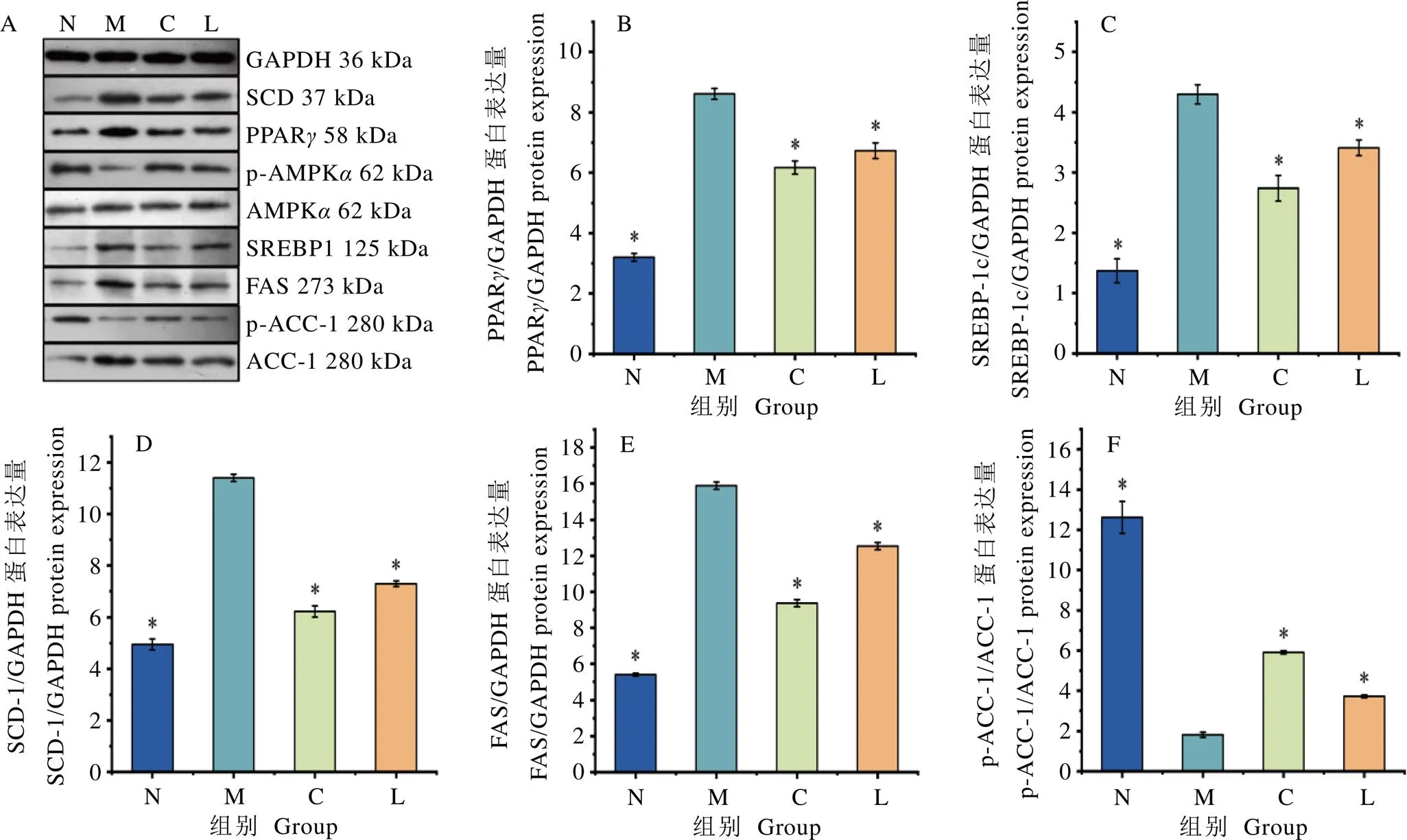
小鼠肝臟蛋白免疫印跡結果如圖5A所示。分析圖5A和圖5B可知,與高脂組相比,正常組、陽性組和龍井組PPAR相對表達量顯著降低(<0.05);由圖5A、圖5C~F可知,正常組、陽性組和龍井組中SREBP-1c、SCD-1、FAS蛋白表達量顯著低于高脂組(<0.05)。與高脂組相比,陽性組、龍井組p-ACC-1/ACC-1值顯著升高,結果表明,陽性藥物與鳩坑龍井茶都具有影響高脂飲食小鼠肝組織脂質合成代謝過程的效應。
由圖6A可知,正常組、陽性組和龍井組相對表達量顯著低于高脂組(<0.05),說明高脂飲食促使小鼠肝組織相對表達量增加,促進脂質合成代謝。由圖6B可知,高脂組小鼠肝組織中-、、-和-基因相對表達量顯著高于正常組(<0.05);與高脂組相比,經陽性藥物和鳩坑龍井茶干預后,小鼠肝組織-、、-和-基因表達水平顯著下降(<0.05)。圖6C顯示,高脂組小鼠肝組織中-和基因表達水平顯著高于其他組(<0.05),說明高脂組中膽固醇合成顯著增加,鳩坑龍井茶干預能抑制高脂小鼠肝組織膽固醇合成過程。
2.7 小鼠腸道菌群檢測分析
各組小鼠腸道菌群分類操作單元(Operational taxonomic units,OTUs)韋恩圖如圖7所示。檢測分析獲得正常組小鼠腸道菌群有457種OTUs,高脂組、陽性組和龍井組分別獲得327、319、389種OTUs,分別有201、194、229種OTUs與正常組相同,表明龍井組小鼠腸道菌群構成與正常組最為相似。
注:N為正常組,M為高脂組,C為陽性組,L為龍井組
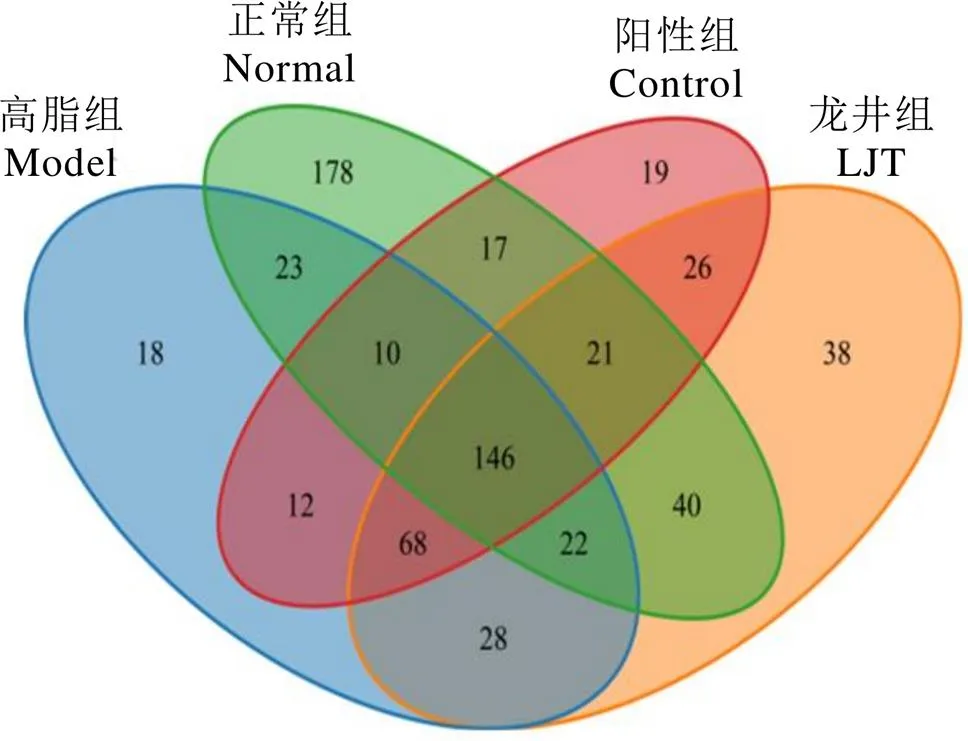
圖7 OTUs韋恩圖
2.7.1 腸道菌群的多樣性分析
由稀釋曲線(圖8A)可知,樣品測序量充分,測序深度足以覆蓋大多數菌群,可進行數據分析。菌群物種的多樣性和豐度由多樣性分析體現(圖8B~D),由Chao1指數圖可見,與正常組相比,高脂組小鼠腸道菌群豐度極顯著降低(<0.01),陽性組和龍井組顯著降低(<0.05);由香農指數和辛普森指數所示,正常組菌群多樣性良好,高脂飲食導致腸道菌群多樣性顯著降低;與高脂組相比,龍井組香農指數顯著增高(<0.05),而正常組、龍井組之間辛普森指數無顯著差異。
多樣性主要通過PCA和PCoA體現出來(圖8E和8F),由PCA圖(PC1和PC2分別占小鼠腸道菌群結構總體差異的58.69%和9.84%)結果可知,正常組與高脂組、陽性組、龍井組腸道菌群組成結構兩兩之間存在差異,但是鳩坑龍井茶干預后小鼠腸道菌群的組成結構趨近于正常組,說明鳩坑龍井茶具有改善高脂飲食導致的小鼠腸道菌群多樣性紊亂效應,并且促使小鼠腸道菌群結構趨向于正常組小鼠腸道菌群生態。
2.7.2 腸道菌群組成分析
腸道菌群各水平上分布如圖9所示。在門水平上主要檢測到Firmicutes、Bacteroidota、Desulfobacterota和Actinobacteriota等4種菌群(圖9A)。與正常組相比,高脂組小鼠腸道菌群中Desulfobacterota的相對豐度增加,并且Firmicutes/Bacteroidota比值(F/B值)提高;經龍井茶干預后,小鼠腸道菌群中Firmicutes相對豐度降低,Bacteroidota相對豐度增加,F/B值降低趨勢明顯,顯示小鼠腸道菌群組成有改善效果。
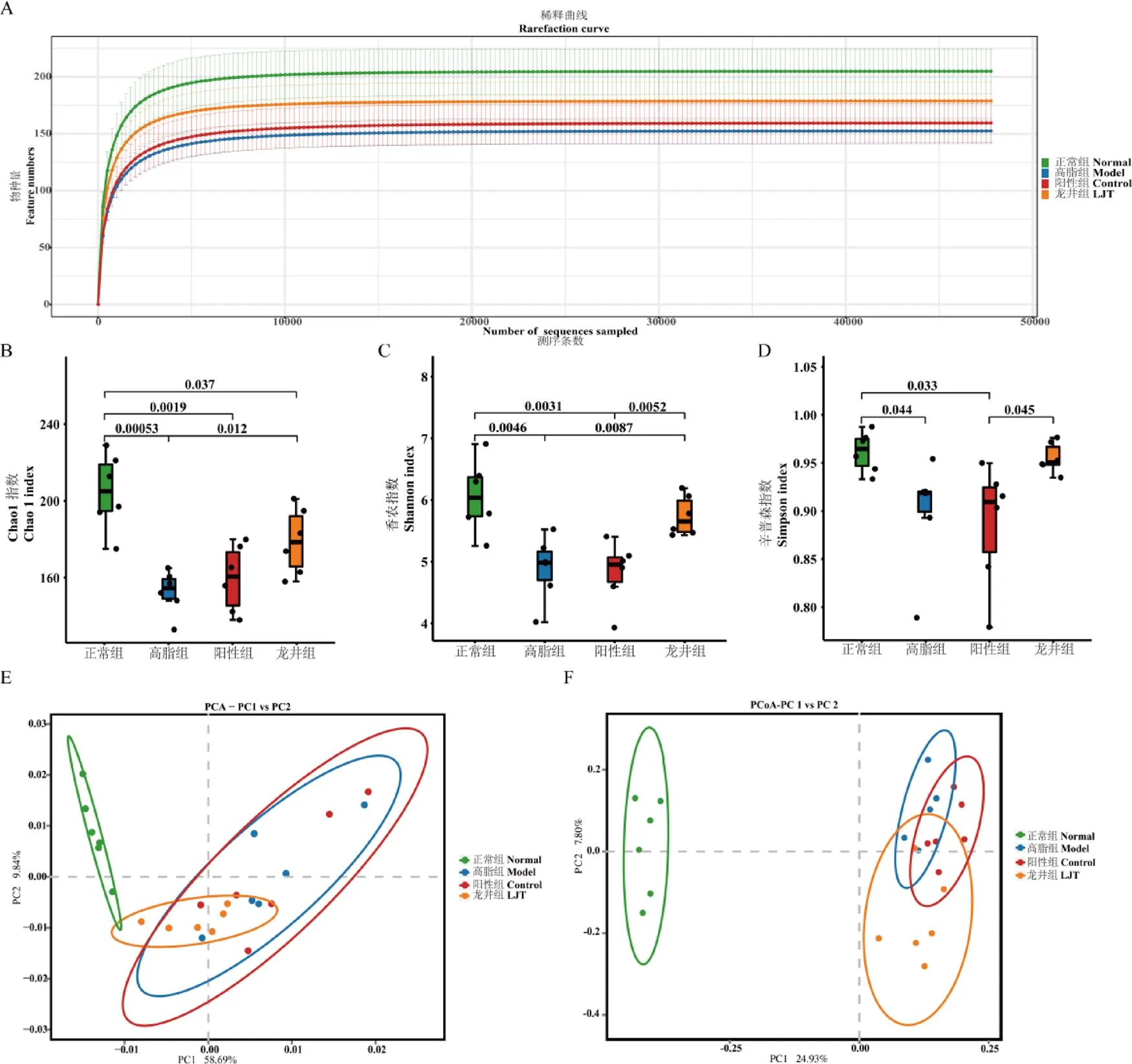
注:A為稀釋曲線,B為Chao1指數,C為香農指數,D為辛普森指數,E為PCA圖,F為PCoA圖
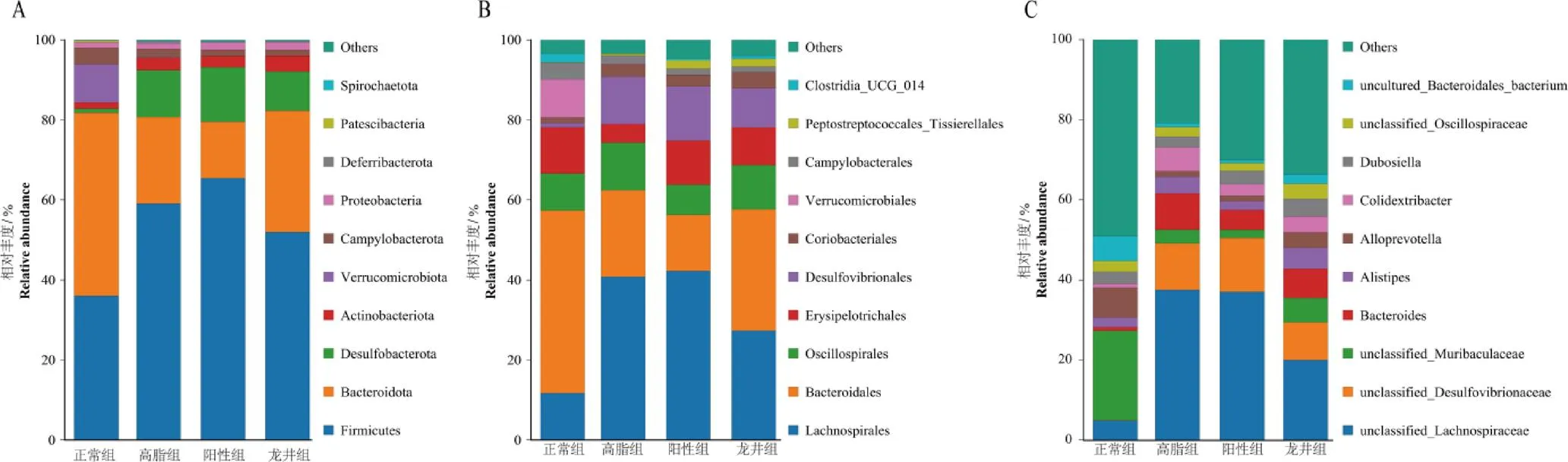
注:A為門水平上菌群分布,B為目水平菌群分布,C為屬水平菌群分布
在目水平上主要檢測到Lachnospirales、Bacteroidales、Oscillospirales、Erysipelotrichales、Desulfovibrionales、Coriobacteriales等菌群(圖9B)。與正常組相比,高脂組小鼠腸道菌群中Lachnospirales、Desulfovibrionales、Oscillospirales、Coriobacteriales、Peptostreptococcales_Tissierellales的相對豐度升高,而Bacteroidales、Erysipelotr-ichales、Verrucomicrobiales、Campylobacterales和Clostridia_UCG_014的相對豐度降低;陽性組小鼠腸道菌群中Lachnospirales、Peptostre-ptococcales_Tissierellales、Erysipelotrichales和Clostridia_UCG_014相對豐度進一步升高,Bacteroidales、Oscillospirales和Campylobac-terales相對豐度降低;龍井組小鼠腸道菌群中Lachnospirales、Desulfovibrionales和Campylobacterales相對豐度降低,同時Bacteroidales、Erysipelotrichales、Coriobacteriales、Peptostreptococcales_Tissierellales和Clostridia_UCG_014相對豐度升高。
小鼠的腸道菌群在屬水平的分布如圖9C所示。與正常組相比,高脂組小鼠腸道菌群中、、、和的相對豐度上調,同時、和相對豐度下調;陽性組小鼠腸道菌群經陽性藥物干預后,、、和相對豐度下降,而相對豐度上升;龍井組小鼠腸道菌群經鳩坑龍井茶干預后,、、和相對豐度下降,-、、、和相對豐度上升。
2.7.3 LEfSe分析
經LEfSe多級物種差異線性判別分析(LDA≥4),各組小鼠體內菌群變化結果如圖10所示。正常組小鼠腸道中主要標志性菌屬為、、、、、和;高脂組小鼠主要標志性菌屬為和;陽性組小鼠主要標志性菌屬為、、和;龍井組標志性菌屬主要為。
2.7.4 KEGG通路功能性預測分析
由圖11A所示,與正常組相比,高脂組下調花生四烯酸代謝、磷酸鹽和次磷酸鹽代謝、核黃素代謝、丙氨酸代謝、-亞麻酸代謝、維他命B6代謝、煙酸和煙酰胺代謝、亞油酸代謝、-谷氨酰胺和-谷氨酸代謝、入脂代謝、磷酸肌醇代謝和甘氨酸,絲氨酸和蘇氨酸代謝,上調甲烷代謝、C5-支鏈二元酸代謝、2-氧羧酸代謝、卟啉與葉綠素代謝和半胱氨酸、蛋氨酸代謝。由圖11B所示,與高脂組相比,龍井組下調次生代謝的生物合成、C5-支鏈二元酸代謝、淀粉和蔗糖代謝、氰基氨基酸代謝和氮代謝,推測鳩坑龍井茶具有調節C5-支鏈二元酸代謝作用。
2.7.5 小鼠腸道菌群豐度與生化指標相關性分析
腸道菌群豐度與小鼠生化指標相關性結果如圖12A所示。小鼠體質量與相對豐度負相關;TG含量與和相對豐度負相關,與相對豐度正相關;LDL-C含量與、、和相對豐度負相關,與和相對豐度正相關;AST活性水平與、、和-相對豐度負相關,與和相對豐度正相關。其中,與小鼠體質量及血清指標相關性最高的腸道菌種為和。
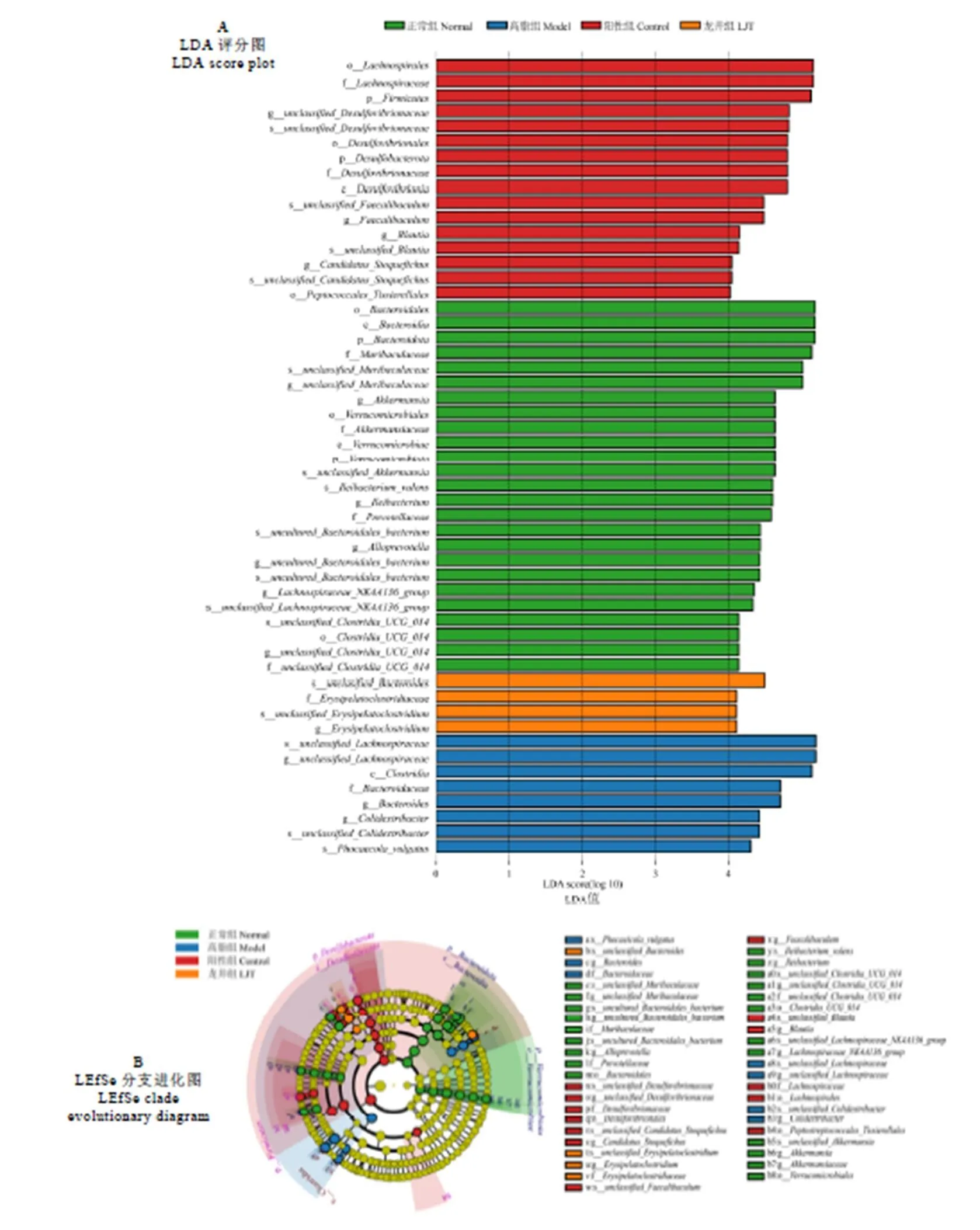
圖10 LEfSe分析
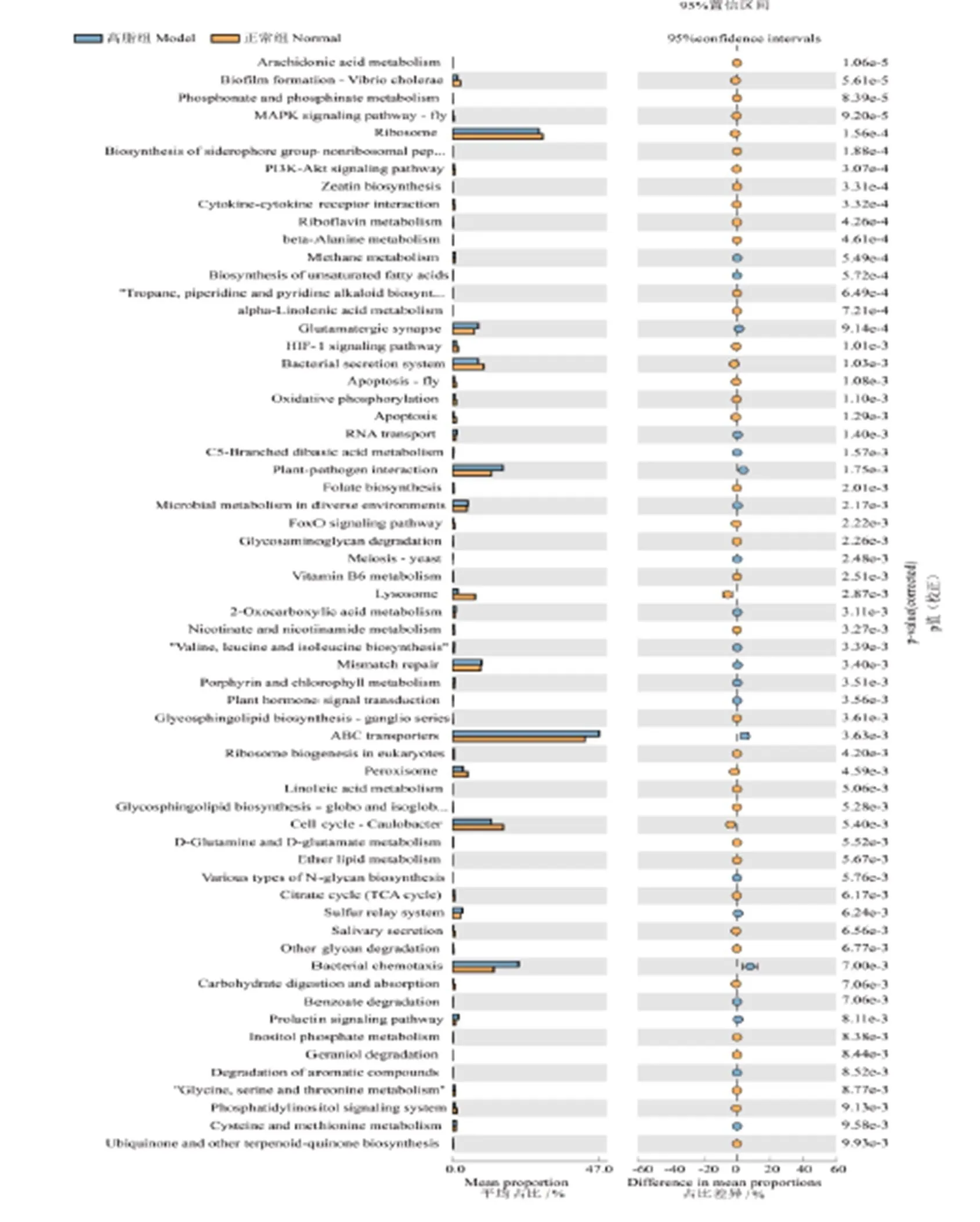
注:A為正常組與高脂組KEGG對比(由于數據過多,僅展示人類疾病分類且P<0.01),B為高脂組與龍井組KEGG對比
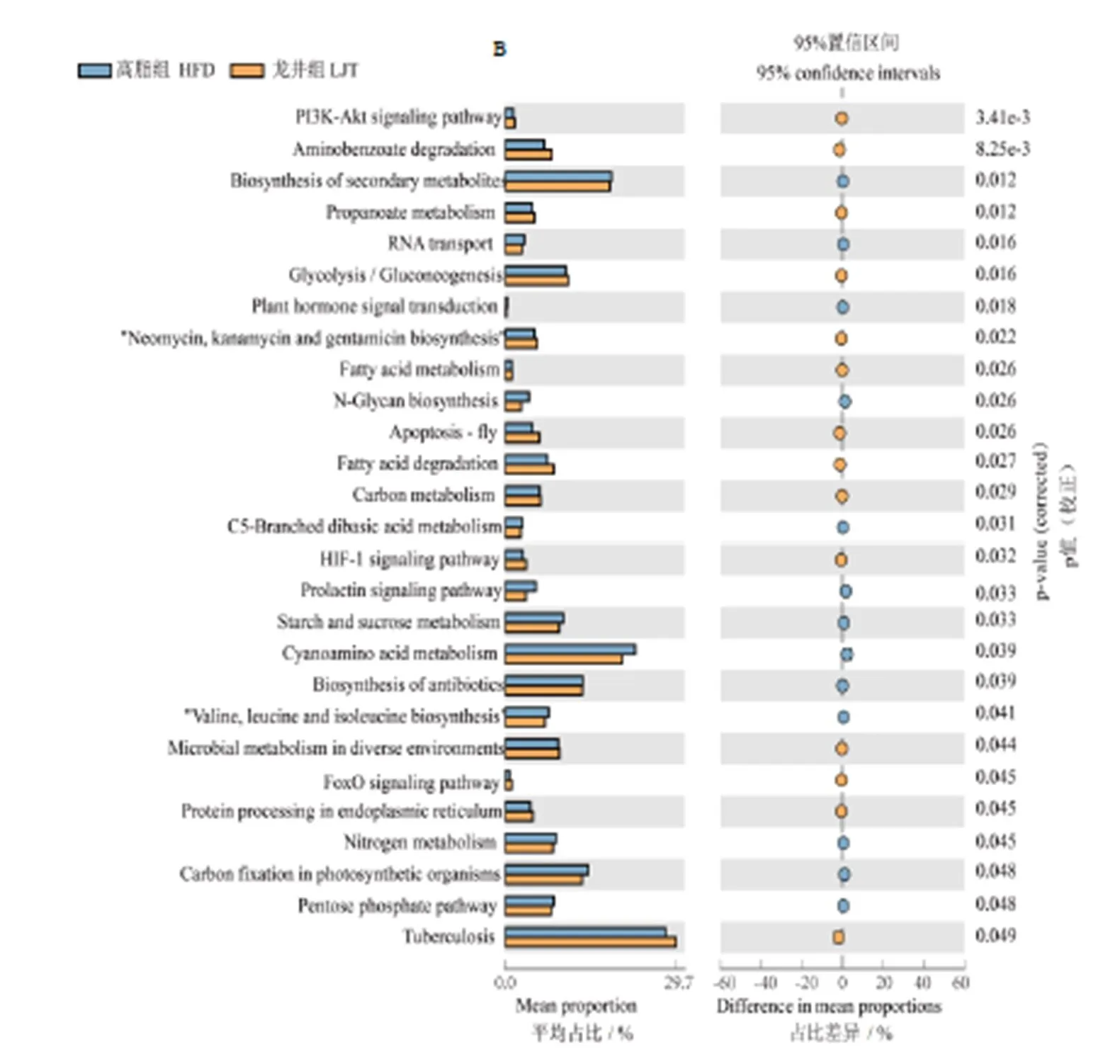
續圖11
Continued Fig.11
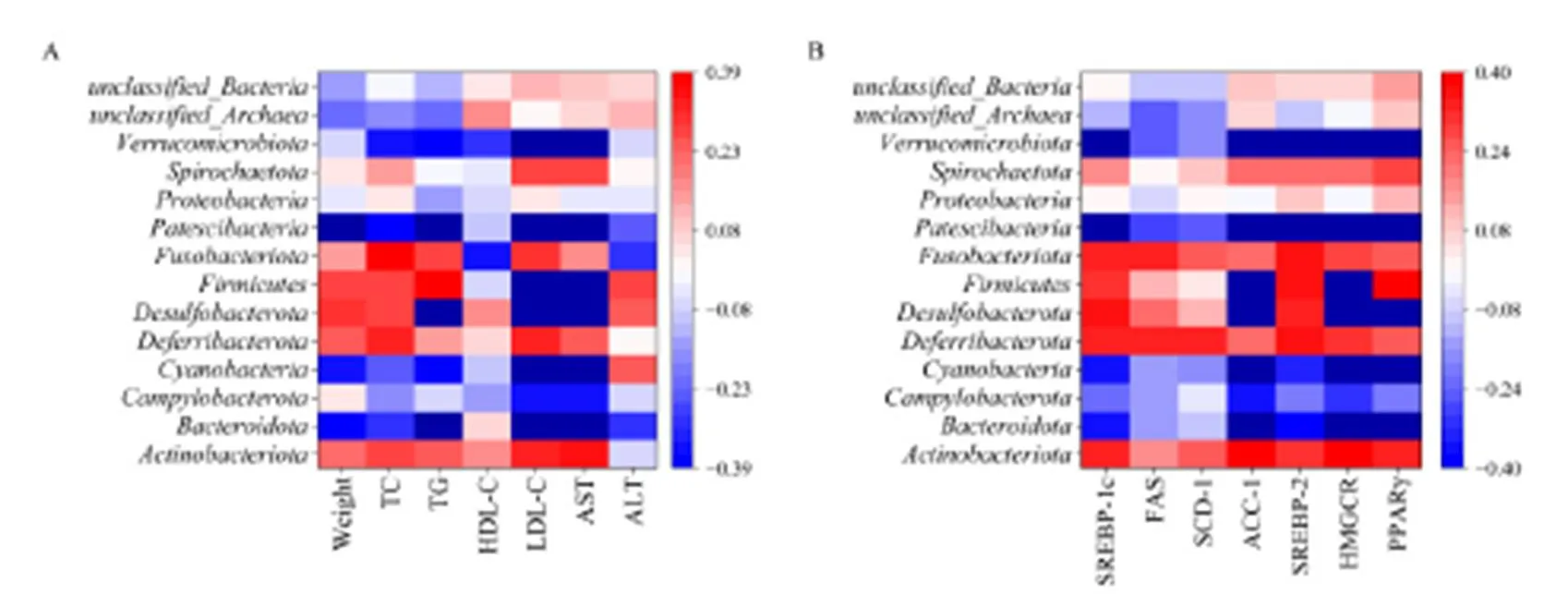
注:A為小鼠體質量、血清生化指標與腸道菌群豐度相關性,B為小鼠肝脂質代謝基因相對表達量與腸道菌群豐度相關性
腸道菌群豐度與小鼠肝脂質代謝通路基因相對含量相關性分析如圖12B所示。SREBP-1基因相對含量與Patescibacteria、Verrucomicrobiota和unclassified_Archaea相對豐度呈負相關,ACC-1基因相對含量與Bacteroidota、Cyanobacteria、Patescibacteria和Verrucomicrobiota相對豐度呈負相關,與Desulfobacterota和Firmicutes呈正相關;SREBP-2基因相對含量與Patescibacteria和Verrucomicrobiota相對豐度呈負相關;HMGCR基因相對含量與Bacteroidota、Cyanobacteria、Patescibacteria和Verrucomicrobiota相對豐度呈正相關;與Desulfobacterota和Firmicutes相對豐度呈正相關;PPAR基因相對含量與Bacteroidota、Campylobacterota、Cyanobacteria、Patescibacteria和Verrucomicrobiota相對豐度呈負相關,與Desulfobacterota相對豐度呈正相關。其中,小鼠肝脂質代謝基因相對含量與Verrucomicrobiota、Bacteroidota和Cyanobac-teria腸道菌種豐度相關性最高。
3 討論
本研究以C57BL/6小鼠為研究對象,經9周高脂飲食誘導模型組小鼠血脂升高、肝組織脂肪變性、糖耐受能力下降和腸道菌群紊亂,與相關研究報道結果具有相似性[17-20],進一步佐證本研究中NAFL小鼠造模成功的科學性和可信度。與高脂組相比,經龍井茶水提物干預,小鼠的體質量、血脂水平、肝組織脂肪變性癥狀得到緩解,小鼠腸道菌群多樣性和豐度明顯增加并趨于正常組腸道菌群生態。
在脂肪代謝過程中,PPAR是啟動脂肪細胞分化的起點[21]。PPAR在脂肪分化方面具有決定性作用[22]。SREBP-1c可靶向促進脂質合成基因-、、-的表達,推動脂肪酸的合成,在TG和TC合成中發揮重要作用[23],其異常高表達會導致肝TG積累和脂代謝紊亂。通過龍井茶水提物干預,促使高脂飲食誘導的小鼠肝組織脂肪合成關鍵酶AMPK活性增強,調控脂質代謝通路信號p-ACC-1/ACC-1比例升高,降低ACC-1蛋白質表達水平,從而抑制脂肪酸合成,減少肝組織脂質蓄積[24]。本研究結果顯示,鳩坑龍井茶水提物具有抑制脂質代謝通路SREBP-1c及其下游脂聯素基因、和-的相對表達量,阻斷肝組織脂肪持續堆積,延緩肝組織脂肪變性,進而緩解NAFLD的進展。
SREBPs存在3種形式的亞型,除SREBP-1c外還存在SREBP-1a和SREBP-2[25]。SREBP-2是體內參與調節細胞膽固醇代謝合成相關酶類的基因調控因子,可直接作用于TC合成途徑的限速酶HMGCR,從而直接調節膽固醇的合成,同時還反饋調節減少內源性膽固醇的生物合成[26]。本研究結果顯示,鳩坑龍井茶水提物具有降低小鼠肝組織-和基因相對表達量,減少肝臟內膽固醇從頭合成。
人體腸道中寄生的細菌數量有近1014,調控人體對攝入飲食的消化吸收,是決定攝入能量轉化率的重要組分。本研究結果顯示,鳩坑龍井茶水提物能夠改善NAFLD小鼠的腸道菌群結構,使其接近正常小鼠腸道菌群的生態。腸道菌群在門水平中的主要菌為Bacteroidota、Firmicutes、Actinobacteriota和Proteobacteria,占腸道菌群的98%,腸道和肝臟通過門靜脈直接關聯,通過“腸-肝軸”影響肝臟脂質代謝[27],腸道菌群失衡是促進NAFLD的重要因素[28]。已有研究報道,高脂飲食C57BL/6小鼠與正常飲食小鼠相比,腸道菌群中厚壁菌門的種類和數量增加,而擬桿菌門的種類和數量下降[29],F/B值升高[30],F/B值常被用來作為腸道菌群有序性的評判標準。本研究結果證實了高脂飲食能夠引起小鼠腸道菌群中F/B值升高,而龍井組小鼠腸道菌群F/B值下降,表明鳩坑龍井水提物具有調節和改善小鼠腸道菌群紊亂的作用效應。
Guo等[31]研究發現,在結腸中產生H2S,抑制結腸上皮細胞呼吸作用[32],抑制丁酸鹽等短鏈脂肪酸(SCFAs)的生成。本研究發現,在高脂組小鼠腸道菌群中相對豐度上升,而在龍井組相對豐度下降。Carter等[33]研究發現,和能促進丁酸鹽的產生,相對豐度與丙酸含量有很強的相關性[34],丁酸可刺激食欲抑制激素分泌,促脂質代謝從而減緩NAFLD[35-36]。本研究分析發現,鳩坑龍井茶水提物干預后,小鼠腸道菌群中和Erysipelotrichales相對豐度增加,表明鳩坑龍井茶水提物具有促進腸道菌群代謝產生友好型短鏈脂肪酸的效應,進而緩解小鼠NAFLD癥狀。
[1] Hardy T, Oakley F, Anstee Q M, et al. Nonalcoholic fatty liver disease: pathogenesis and disease spectrum [J]. Annual Review of Pathology, 2016, 11: 451-496.
[2] Paternostro R, Trauner M. Current treatment of non-alcoholic fatty liver disease [J]. Journal of Internal Medicine, 2022, 292(2): 190-204.
[3] Brouwers M C G J, Simons N, Stehouwer C D A, et al. Non-alcoholic fatty liver disease and cardiovascular disease: assessing the evidence for causality [J]. Diabetologia, 2020, 63(2): 253-260.
[4] Carmiel-Haggai M, Cederbaum A I, Nieto N. A high-fat diet leads to the progression of non-alcoholic fatty liver disease in obese rats [J]. The FASEB Journal, 2005, 19(1): 136-138.
[5] El-Koofy N M, Anwar G M, El-Raziky M S, et al. The association of metabolic syndrome, insulin resistance and non-alcoholic fatty liver disease in overweight/obese children [J]. Saudi Journal of Gastroenterology, 2012, 18(1): 44-49.
[6] Powell E E, Wong V W, Rinella M. Non-alcoholic fatty liver disease [J]. Lancet, 2021, 397(10290): 2212-2224.
[7] Bashiardes S, Shapiro H, Rozin S, et al. Non-alcoholic fatty liver and the gut microbiota [J]. Molecular Metabolism, 2016, 5(9): 782-794.
[8] Charroux B, Royet J. Gut-derived peptidoglycan remotely inhibits bacteria dependent activation of SREBP byadipocytes [J]. Plos Genetics, 2022, 18(3): e1010098. doi: 10.1371/journal.pgen.1010098.
[9] 朱蔭, 邵晨陽, 張悅, 等. 不同茶樹品種龍井茶香氣成分差異分析[J]. 食品工業科技, 2018, 39(23): 241-246, 254. Zhu Y, Shao C Y, Zhang Y, et al. Comparison of differences in aroma constituents of Longjing tea produced from different tea germplasms [J]. Science and Technology of Food Industry, 2018, 39(23): 241-246, 254.
[10] 孫達, 龔恕, 崔宏春, 等. 不同品種茶樹春秋季鮮葉超微綠茶粉適制性研究[J]. 浙江農業學報, 2021, 33(3): 437-446. Sun D, Gong S, Cui H C, et al. Suitablity of fresh spring and autumn leaves from different tea cultivars for ultramicro green tea powder production [J]. Acta Agriculturae Zhejiangensis, 2021, 33(3): 437-446.
[11] 王素敏, 徐歡歡, 黃業偉, 等. 茶多酚的降脂作用及其機制研究進展[J]. 食品研究與開發, 2016, 37(10): 219-224. Wang S M, Xu H H, Huang Y W, et al. Review on the effect of tea polyphenols in hypolipidemic and its medchanism [J]. Food Research and Development, 2016, 37(10): 219-224.
[12] Li B Y, Li H Y, Zhou D D, et al. Effects of different green tea extracts on chronic alcohol induced-fatty liver disease by ameliorating oxidative stress and inflammation in mice [J]. Oxid Med Cell Longev, 2021, 2021: 5188205. doi: 10.1155/2021/5188205.
[13] Ma H, Zhang B, Hu Y, et al. The novel intervention effect of cold green tea beverage on high-fat diet induced obesity in mice [J]. Journal of Functional Foods, 2020, 75: 104279. doi: 10.1016/j.jff.2020.104279.
[14] 王忠民, 吳謀成, 李小定, 等. 葡萄多糖的提取及含量測定[J]. 新疆農業大學學報, 2002, 25(2): 57-58. Wang Z M, Wu M C, Li X D, et al. Extraction and contents, measurement of the VLP [J]. Journal of Xinjiang Agricultural University, 2002, 25(2): 57-58.
[15] Li M, Xu J, Zhang Y, et al. Comparative analysis of fecal metabolite profiles in HFD-induced obese mice after oral administration of Huangjinya green tea extract [J]. Food and Chemical Toxicology, 2020, 145: 111744. doi: 10.1016/j.fct.2020.111744.
[16] 馮琳, 龔自明, 劉盼盼, 等. 青磚毛茶對高脂飲食小鼠腸道微生物的影響[J]. 中國食品學報, 2021, 21(7): 87-96. Feng L, Gong Z M, Liu P P, et al. Effects of Qingzhuan Maocha on gut microbiota in high-fat diet fed mice [J]. Journal of Chinese Institute of Food Science and Technology, 2021, 21(7): 87-96.
[17] Ma H, Zhang B, Hu Y, et al. Correlation analysis of intestinal redox state with the gut microbiota reveals the positive intervention of tea polyphenols on hyperlipidemia in high fat diet fed mice [J]. Journal of Agricultural and Food Chemistry, 2019, 67(26): 7325-7335.
[18] Velázquez K T, Enos R T, Bader J E, et al. Prolonged high-fat-diet feeding promotes non-alcoholic fatty liver disease and alters gut microbiota in mice [J]. World Journal of Hepatology, 2019, 11(8): 619-637.
[19] Aydos L R, Do Amaral L A, De Souza R S, et al. Nonalcoholic fatty liver disease induced by high-fat diet in C57BL/6 models [J]. Nutrients, 2019, 11(12): 3067. doi: 10.3390/nu11123067.
[20] Yin J, Li Y, Han H, et al. Melatonin reprogramming of gut microbiota improves lipid dysmetabolism in high-fat diet-fed mice [J]. Journal of Pineal Research, 2018, 65(4): e12524. doi: 10.1111/jpi.12524.
[21] 黃玉晶. 鄰苯二甲酸酯母體暴露對早產和胎兒發育的影響及過氧化物酶體增殖物激活受體在其中的作用研究[D]. 重慶: 第三軍醫大學, 2014. Huang Y J. The role of peroxisome proliferator activated receptor in the effects of gestation exposure to phthalates on preterm delivery and fetal development [D]. Chongqing: Army Medical University, 2014.
[22] 劉亞茹, 苗志國, 高明磊, 等. PPAR在動物脂肪發育中的研究進展[J]. 黑龍江畜牧獸醫, 2019(1): 32-35. Liu Y R, Miao Z G, Gao M L, et al. Research advance on PPARin animal adipose tissue [J]. Heilongjiang Animal Science and Veterinary Medicine, 2019(1): 32-35.
[23] Shi W, Hou T, Guo D, et al. Evaluation of hypolipidemic peptide (Val-Phe-Val-Arg-Asn) virtual screened from chickpea peptides by pharmacophore model in high-fat diet-induced obese rat [J]. Journal of Functional Foods, 2019, 54: 136-145.
[24] Geng T T, Liu Y, Xu Y T, et al. H19 lncRNA promotes skeletal muscle insulin sensitivity in part by targeting AMPK [J]. Diabetes, 2018, 67(11) : 2183-2198.
[25] Loregger A, Raaben M, Nieuwenhuis J, et al. Haploid genetic screens identify SPRING/C12ORF49 as a determinant of SREBP signaling and cholesterol metabolism [J]. Nature Communications, 2020, 11: 1128. doi: 10.1038/s41467-020-14811-1.
[26] Wei S, Espenshade P J. Expanding roles for SREBP in metabolism [J]. Cell Metabolism, 2012, 16(4): 414-419.
[27] Milosevic I, Vujovic A, Barac A, et al. Gut-liver axis, gut microbiota, and its modulation in the management of liver diseases: a review of the literature [J]. International Journal of Molecular Sciences, 2019, 20(2): 395. doi: 10.3390/ijms20020395.
[28] Soderborg T K, Clark S E, Mulligan C E, et al. The gut microbiota in infants of obese mothers increases inflammation and susceptibility to NAFLD [J]. Nature Communications, 2018, 9: 4462. doi: 10.1038/s41467-018-06929-0.
[29] Ley R E, B?ckhed F, Turnbaugh P, et al. Obesity alters gut microbial ecology [J]. PNAS, 2005, 102(31): 11070-11075.
[30] Do M H, Lee H B, Oh M J, et al. Polysaccharide fraction from greens ofalleviates high fat diet-induced obesity [J]. Food Chemistry, 2021, 33:128395. doi: 10.1016/j.foodchem.2020.128395.
[31] Guo J L, Han X, Zhan J C, et al. Vanillin alleviates high fat diet-induced obesity and improves the gut microbiota composition [J]. Frontiers in Microbiology, 2018, 9: 2733. doi: 10.3389/fmicb.2018.02733.
[32] Beaumont M, Andriamihaja M, Lan A, et al. Detrimental effects for colonocytes of an increased exposure to luminal hydrogen sulfide: the adaptive response [J]. Free Radical Biology and Medicine, 2016, 93: 155-164.
[33] Carter J K, Bhattacharya D, Borgerding J N, et al. Modeling dysbiosis of human NASH in mice: loss of gut microbiome diversity and overgrowth of[J]. Plos One, 2021, 16(1): e0244763.
[34] Smith B J, Miller R A, Ericsson A C, et al. Changes in the gut microbiome and fermentation products concurrent with enhanced longevity in acarbose-treated mice [J]. BMC Microbiology, 2019, 19: 130. doi: 10.1186/s12866-019-1494-7.
[35] Musso G, Gambino R, Cassader M. Obesity, diabetes, and gut microbiota: the hygiene hypothesis expanded? [J]. Diabetes Care, 2010, 33(10): 2277-2284.
[36] Den Besten G, Bleeker A, Gerding A, et al. Short-chain fatty acids protect against high-fat diet-induced obesity via a PPAR-dependent switch from lipogenesis to fat oxidation [J]. Diabetes, 2015, 64(7): 2398-2408.
Effect of JiukengLongjing Tea on SREBPs Signaling Pathway and Gut Microbiota Regulation in High-fat Diet C57BL/6 Mice with Hepatic Steatosis
GONG Mingxiu1,2, YUAN Yiwei1,2, ZHANG Yifan1,2, YE Jiangcheng1,2, GUO Li3, LI Xiaojun4, HUANG Hao4, MAO Yuxiao5, ZHAO Yun5, ZHAO Jin1,2*
1. Institute of Food Nutrition and Quality Safety, College of Life Sciences, China Jiliang University, Hangzhou 310018, China; 2. Key Laboratory of Pecialty Agri-product Quality and Hazard Controlling Technology of Zhejiang Province, Hangzhou 310018, China; 3. TeaResearch Institute, Chinese Academy of Agricultural Sciences, Hangzhou 310008, China; 4. Doctor Innovation Workstation of Zhejiang Yifutang Tea Industry Co., Ltd., Hangzhou 311500, China; 5. Hangzhou Academy of Agricultural Sciences, Hangzhou 310024, China
To investigate the effect of Jiukeng Longjing tea water extract (LJT) on liver steatosis and the regulation of gut microbiota in C57BL/6 mice fed with high-fat diet, a non-alcoholic fatty liver model was established in mice induced by a high-fat diet, and LJT (300?mg·kg-1) was gavaged for intervention. The body weight of mice was recorded regularly, and serum biochemical indicators such as AST, ALT, TC, TG, LDL-C, HDL-C, and glucose tolerance levels were measured. The characteristics of HE staining and oil red O staining liver tissue sections were observed and analyzed. Real-time qPCR technology was used to detect the expressions of seven genes including,,,,,, andin mouse liver tissues. The relative expressions of proteins related to lipid metabolism were studied by western blot. At the same time, the gut microbiota of mice was sequenced by high-throughput sequencing (16?S rDNA) and its structure was analyzed. The results show that the body weight, blood glucose AUC, serum TG, TC, LDL-C, and liver TG, TC levels significantly decreased under LJT intervention. Western blot shows that LJT intervention reduced the expressions of SREBP-1c, FAS, ACC-1, SCD-1, and PPARin liver tissue of mice. LJT also significantly downregulated the relative expressions of,,,,,andin liver tissue. The 16?S rDNA detection reveals that the levels of gut microbiota were mainly classified into four categories: Firmicutes, Bacteroidota, Desulfobacterota, and Actinobaciota. LJT could effectively alleviate the trend of increasing the relative abundance of Firmicutes and decreasing the relative abundance of Bacteroidota caused by high-fat diet, and increase the species abundance of gut microbiota. Therefore, LJT could interfere with the signal expression of SREBPs pathway in mouse liver steatosis, and improve the disturbance of gut microbiota in mice, thereby achieve the effect of reducing fat and weight loss.
Jiukeng Longjing tea, non-alcoholic fatty liver, SREBPs path, gut microbiota dysbiosis, lipid-lowering effect
S571.1,R151.3
A
1000-369X(2023)04-576-17
2023-01-16
2023-04-12
浙江省重點研發計劃項目(2020C02045)、杭州市農業與社會發展科研項目(202203A06、202203A11、202203B09)、開化茶產業提升浙江省團隊科技特派員項目、杭州市科技特派員項目(20221122I80)、衢州市重點科技攻關項目科技強農專項(2023k098)
龔明秀,女,碩士研究生,主要從事藥食同源植物營養與功效評價研究。*通信作者:zhaojin@cjlu.edu.cn
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
