抑制IP3R-Ca2+途徑對對乙酰氨基酚所致肝損傷及其線粒體內質網結構偶聯的影響
2023-08-02 02:58:40徐王婷宋育林
安徽醫科大學學報 2023年7期
徐王婷,宋育林
對乙酰氨基酚(acetaminophen, APAP)因具有解熱、鎮痛的作用而被廣泛應用于臨床,但是攝入過量APAP會引起嚴重的肝損傷。APAP所致肝損傷的確切機制尚不明確,可能與乙酰苯醌亞胺(N-acetyl-p-benzoquinone imine, NAPQI)形成、線粒體損傷、內質網應激等有關[1]。線粒體內質網結構偶聯(mitochondrial-associated endoplasmic reticulum membranes,MAMs)是線粒體和內質網之間存在的特殊的物理和生化連接區域,富集著多種蛋白質,如1,4,5-三磷酸肌醇受體(inositol 1,4,5-trisphate receptor, IP3R)、葡萄糖調節蛋白75(glucose regulated protein 75,GRP75)、線粒體融合蛋白2(mitofusin 2, MFN2)等,參與內質網應激、維持鈣穩態和線粒體形態、脂質合成和運輸等過程[2],并與非酒精性脂肪肝等慢性肝病有關[3]。2-氨基乙氧基二苯基硼酸鹽(2-aminoethoxydiphenyl borate,2-APB)是IP3R抑制劑[4],1,2-雙(2-氨基苯氧基)-乙烷-N, N, N′N′-四乙酸[1,2-bis(2-aminophenoxy)ethane-N,N,N′N′-tetraacetic acid,BAPTA-AM]是公認的鈣離子螯合劑[5]。為此,該研究通過觀察2-APB、BAPTA-AM對APAP所致小鼠肝損傷及其肝臟超微結構、肝組織Ca2+水平、MAMs相關蛋白線粒體融合蛋白1(mitofusin 1,MFN1)、MFN2、IP3R1、GRP75表達的影響,探討抑制IP3R-Ca2+途徑對APAP所致肝損傷及其MAMs的影響。
1 材料與方法
1.1 實驗動物40只健康的SPF級雄性C57BL/6小鼠,鼠齡7周,體質量18~22 g,購自杭州子源實驗動物科技有限公司;動物許可證號:SCXK(浙)2019-0004。
1.2 試劑與儀器APAP(純度大于99%)和BAPTA-AM(純度大于99%)購自美國MedChemExpress公司;2-APB購自美國Sigma公司(純度97%);丙氨酸氨基轉移酶(alanine aminotransferase,ALT)、天門冬氨酸氨基轉移酶(aspartate aminotransferase,AST)檢測試劑盒為羅氏診斷產品(上海) 有限公司生產;GRP75、GAPDH抗體為合肥朵能生物科技有限公司產品(貨號:AB011901、AB010301);MFN1、MFN2及IP3R1抗體購自美國Signalway Antibody公司(貨號:55628、33015、29549);山羊抗小鼠二抗及山羊抗兔二抗均購自美國Proteintech公司(貨號:SA00001-1、SA00001-2);鈣離子含量檢測試劑盒及總蛋白定量測試盒購自南京建成生物工程研究所;SDS-PAGE凝膠制備試劑盒購自北京索萊寶科技有限公司;Roche Cobas 8000全自動生化分析儀購自瑞士Roche公司;酶標儀產自美國Themo公司;透射電鏡JEM-1400flash產自日本電子株式會社;化學發光成像分析系統ZF-670產自上海嘉鵬科技有限公司。
1.3 實驗分組與處理40只小鼠隨機分為正常對照組、模型組、IP3R抑制劑(2-APB)組、鈣離子螯合劑(BAPTA-AM)組,每組10只。模型組、2-APB組、BAPTA-AM單次腹腔注射APAP(300 mg/kg)。注射APAP前30 min,2-APB組腹腔注射2-APB (20 mg/kg),BAPTA-AM組腹腔注射 BAPTA-AM(2.5 mg/kg)。腹腔注射APAP后,各組小鼠均禁食不禁水。APAP造模24 h后,稱量小鼠體質量,麻醉后摘除小鼠眼球取血,斷頭處死,取出肝臟。血標本室溫靜置2 h,3 500 r/min離心15 min,制備血清,保存于-80 ℃冰箱中。取出肝臟后,稱量肝濕重,觀察肝臟外觀情況,剪取部分肝左葉用4%甲醛緩沖液固定行肝臟病理學檢查;取相同部位約1 mm3左右的肝組織若干塊,固定于2.5%戊二醛固定液,4 ℃冷藏,用于透射電鏡觀察;余肝臟組織于液氮中保存,后轉移至-80 ℃冰箱貯存。
1.4 肝指數根據前述所稱量的小鼠處死前體質量和肝濕重計算肝指數。肝指數(%)=(肝濕重/處死前小鼠體質量) ×100%。
1.5 肝功能指標檢測全自動生化分析儀檢測小鼠血清ALT、AST水平。
1.6 肝組織病理觀察取4%甲醛緩沖液固定的肝組織,予以脫水、石蠟包埋、切片和HE染色,光鏡下觀察肝組織結構。
1.7 透射電鏡超微結構觀察2.5%戊二醛緩沖液固定的肝臟標本,按常規透射電鏡技術脫水、包埋、超薄切片、醋酸雙氧鈾-檸檬酸鉛雙染色,透射電鏡下觀察切片并采集圖像進行分析。
1.8 鈣離子含量測定取適量組織塊,經去離子水中漂洗后,按1 ∶9 [質量(g) ∶體積(ml)]比例加入去離子水,制成10%的勻漿液,以2 500 r/min離心15 min后取上清液,采用考馬斯亮藍法測定蛋白濃度,按照鈣測試盒說明書步驟用微板法檢測鈣離子含量。
1.9 Western blot檢測肝組織中MFN1、MFN2、GRP75、IP3R1蛋白表達取適量的肝組織,提取組織總蛋白,用BCA法進行蛋白定量后,進行SDS-PAGE電泳,再轉膜、封閉,將膜依次與相對應的一抗與二抗孵育。工作濃度:GAPDH 1 ∶2 000、抗MFN1抗體 1 ∶1 000、抗MFN2抗體1 ∶1 000、抗GRP75抗體1 ∶1 000、抗IP3R1抗體1 ∶1 000、二抗1 ∶5 000。最后,ECL顯影,Image J軟件分析所得圖像,計算目標蛋白與GAPDH的灰度值比值。
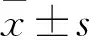
2 結果
2.1 肝指數變化模型組小鼠較正常對照組的肝濕重及肝指數均增加(P<0.01);2-APB組及BAPTA-AM組小鼠較模型組的肝濕重及肝指數均降低(P<0.01)。見表1。

表1 各組小鼠肝指數變化
2.2 肝功能變化與正常對照組相比,模型組小鼠ALT、AST水平增加,差異有統計學意義(P<0.01)。與模型組相比,2-APB組及BAPTA-AM組小鼠ALT、AST水平降低,差異有統計學意義(P<0.01),見表2。

表2 各組小鼠血清ALT、AST變化
2.3 肝組織病理學改變正常對照組小鼠肝小葉結構完整,肝細胞形態正常。模型組小鼠的肝臟可見肝小葉結構紊亂,小葉中央靜脈周圍可見大片肝細胞壞死,并見肝竇充血及炎細胞浸潤。相比于模型組,2-APB組及BAPTA-AM組小鼠肝小葉結構尚完整,可見肝細胞變性,肝細胞壞死面積均明顯減少。見圖1。

圖1 各組小鼠肝組織病理學變化 HE×200
2.4 肝細胞超微結構變化透射電鏡下,正常對照組小鼠線粒體未腫脹,線粒體嵴完整;模型組可見線粒體腫脹,線粒體嵴結構模糊、斷裂、消失,內質網腫脹斷裂,MAMs數量增加;2-APB組及BAPTA-AM組線粒體腫脹程度明顯減輕,部分線粒體出現嵴斷裂或溶解消失,內質網結構基本完整,MAMs數量減少。見圖2。

圖2 各組小鼠肝臟樣品電鏡檢查結果 ×20 000
2.5 肝組織鈣離子含量的變化正常對照組、模型組、2-APB組和BAPTA-AM組的小鼠肝組織鈣含量分別為(0.031±0.011)、(0.066±0.029)、(0.036±0.011)和(0.040±0.016)mmol/g prot,4組間方差分析結果:F=7.476,P=0.001。與正常對照組比較,模型組小鼠肝組織勻漿中鈣離子含量增加,差異有統計學意義(P<0.01);與模型組比較,2-APB組及BAPTA-AM組小鼠肝組織勻漿中鈣離子含量均降低,差異有統計學意義(P<0.01)。
2.6 肝組織MFN1、MFN2、GRP75、IP3R1蛋白表達情況與正常對照組比較,模型組小鼠肝臟的MFN1、MFN2蛋白表達均減少,而IP3R1、GRP75蛋白的表達增加(P<0.01)。與模型組比較,2-APB組及BAPTA-AM組MFN1、MFN2蛋白表達均增加,而IP3R1、GRP75蛋白的表達減少(P<0.01)。見圖3。

圖3 各組小鼠肝組織MFN1、MFN2、IP3R1、GRP75蛋白表達情況
3 討論
APAP過量是急性肝衰竭的主要原因之一。肝細胞壞死是APAP所致肝損傷的早期表現[1]。既往研究[6]表明,單次腹腔注射APAP(300 mg/kg)24 h后小鼠血清ALT、AST可維持在較高水平,肝臟主要表現為小葉中心性壞死。本研究中,模型組小鼠較正常對照組的肝指數及肝濕重增加,ALT、AST水平升高,肝臟可見小葉中心性壞死,提示APAP致小鼠肝損傷模型復制成功。
有研究[7]顯示,APAP肝損傷時線粒體和細胞核內鈣水平升高,肝細胞內鈣超載可能與瞬時受體電位M2(transient receptor potential melastatin 2,TRPM2)激活有關。MAMs作為線粒體與內質網之間的特殊結構,通過自身數量和(或)所富集的蛋白質的變化,發揮不同的生物學功能,如調控內質網中的Ca2+向線粒體轉運,調控線粒體形態、內質網應激等[2-3]。IP3R是位于內質網膜上的Ca2+釋放通道,電壓依賴性陰離子通道1(voltage-dependent anion channel 1,VDAC1)是位于線粒體外膜的Ca2+通道,而GRP75作為分子伴侶連接IP3R和VDAC1[8]。IP3R與GRP75 結合介導內質網釋放Ca2+,Ca2+通過位于MAMs的復合物 IP3R-GRP75-VDAC1進入線粒體[2-3,8]。干擾GRP75可影響GRP75-IP3R的相互作用從而影響MAMs[9]。IP3R有3個亞型,分別是IP3R1、IP3R2、IP3R3,非酒精性脂肪性肝炎患者肝臟IP3R1表達上調[3]。在線粒體融合過程,MFN1或MFN2形成同型或異型復合物,促進線粒體融合、MAMs穩定和Ca2+的轉運[2]。本研究顯示,與正常對照組相比,模型組小鼠肝細胞線粒體嵴斷裂、消失,內質網腫脹斷裂;MAMs數量增加;肝臟組織中Ca2+含量較正常對照組增加;MFN1、MFN2蛋白的表達水平降低(P<0.01),IP3R1蛋白及GRP75蛋白的表達水平增加,提示IP3R-Ca2+途徑的增強、MAMs及其相關蛋白表達異常參與了APAP所致的肝損傷過程。
2-APB具有多種生物活性,通過抑制IP3R和鈣庫操縱的鈣離子內流(store-operated calcium entry,SOCE)通道降低細胞內Ca2+水平和細胞器之間Ca2+的移動[10]。2-APB可以減輕APAP所致肝損傷[4]。BAPTA-AM可通過減輕細胞內Ca2+超載起到保護細胞作用[5]。本研究顯示,與模型組比較,2-APB組及BAPTA-AM組小鼠血清ALT、AST水平下降,光鏡下肝臟病理變化明顯改善。2-APB的作用與文獻[4]報道相似。表明2-APB及BAPTA-AM均具有保護APAP所致肝損傷的作用,提示抑制IP3R-Ca2+途徑可以減輕APAP所致的肝損傷。其機制可能有以下2種:① 降低肝臟Ca2+含量過載改善線粒體、內質網損傷:鈣離子超載可以影響線粒體的結構和功能[11],并與APAP所致的肝損傷有關[7]。本研究顯示,與模型組比較,2-APB組及BAPTA-AM組小鼠肝組織勻漿中Ca2+含量減少、肝臟線粒體及內質網病理變化減輕。提示2-APB及BAPTA-AM可通過降低肝臟Ca2+超載、改善線粒體及內質網損傷保護APAP所致的肝損傷。② 影響MAMs結構及其相關蛋白的表達:MAMs通過其自身結構和所募集蛋白的變化參與鈣平衡、線粒體功能、內質網應激和細胞死亡等的調控[ 2-3]。抑制GRP75的表達可減低以VDAC1/ITPR1(IP3R1)標志的HuH7細胞MAMs數量;增強GRP75表達則作用相反[12]。文獻報道,APAP中毒可導致小鼠肝臟線粒體MFN2蛋白水平下降[13];酒精性肝損傷大鼠肝臟存在線粒體功能異常和MFN2蛋白表達下調[14]。本研究顯示,與模型組比較,2-APB組及BAPTA-AM組小鼠MAMs數量減少,MFN1、MFN2蛋白表達水平上調,IP3R1及GRP75蛋白表達水平降低。提示2-APB和BAPTA-AM可以通過改變MAMs數量、抑制IP3R1及GRP75和上調MFN1、MFN2蛋白表達減輕APAP所致的肝損傷,確切機制有待進一步探討。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
學苑創造·A版(2020年9期)2020-10-13 09:41:02
數學物理學報(2020年2期)2020-06-02 11:29:24
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
