八角楓內生真菌的分離鑒定及其發酵產物對松材線蟲的毒殺活性
2023-06-26 14:21:36錢雨可于金英賀小如雷秋艷
西北農業學報 2023年7期
錢雨可,于金英,賀小如,雷秋艷,汪 冶
(1.麗水學院,浙江麗水 323000;2.浙江理工大學,杭州 310000)
松樹萎蔫病(Pine wilt disease,PWD)是由松材線蟲(Bursaphelenehusxylophilus)引起的松樹病害,松樹感染松材線蟲之后,發病初期癥狀表現為松針的黃化和萎蔫[1],國內于1982年在南京中山陵首次發現該病[2]。在中國主要為害黑松(Pinusthunbergii)、赤松(P.densiflora)、馬尾松(P.massoniana)、海岸松(P.pinaster)、火炬松(P.taeda)、黃松(P.thunberigii×P.massoniana)等松科植物。松材線蟲與美國白蛾、松圓蚧一同列為森林三大病蟲害,屬國際重要檢疫對象,其中松材線蟲被林業部列為森林病蟲害之首。因此,尋找能夠高效殺滅松材線蟲的生物防治措施、保護松屬植物迫在眉睫。
植物內生菌是一類具有重要應用價值的微生物資源,微生物活性代謝產物可用來研發農藥、獸藥及醫藥。通過對微生物菌株發酵生產具有生物活性的代謝物,并將其用于病蟲害的控制是目前創制生物農藥的重要途徑,如來自葡萄的內生真菌(Alternariaalternata)環肽類次生代謝產物對其寄主植物的病原菌(Plasmoparaviticola)有抑制作用,但對寄主植物沒有毒害作用,目前被作為潛在的生物農藥研究開發[3]。來自銀杏的球毛殼菌(Chaetomiumglobosum)的細胞松弛素類生物堿具有抗植物病原菌作用[4-5]。中國蘊藏豐富的生物資源,利用國內植物內生菌資源篩選抗蟲(尤其是松材線蟲)活性物質,對促進新型生物農藥的開發和創制具有十分重要的意義[6-9]。植物內生真菌種類繁多,目前只有芽孢桿菌、阿姆斯特丹散囊菌和竹生炭角菌[10-11]等極少數內生生防真菌用于殺松材線蟲活性的研究,難以滿足大規模防治需求。本試驗從八角楓科八角楓屬植物八角楓(Alangiumchinense)的莖、葉組織中分離篩選具有毒殺松材線蟲作用的內生真菌,并對活性高的菌株進一步進行形態特征和分子生物學鑒定,為松材線蟲生物農藥制劑的研究和開發提供理論依據[12]。
1 材料與方法
1.1 供試材料
1.1.1 供試植物樣本 2021年9月在浙江省麗水市白云山采集健康八角楓(Alangiumchinense)植物莖、葉組織。
1.1.2 供試線蟲 松材線蟲分離自麗水林區疫木。采用貝爾曼(Baermann)漏斗法對疫木中線蟲進行分離,待大多數線蟲懷卵后同期化處理[13],接入長滿灰葡萄孢菌的培養基,28 ℃恒溫培養2~3 d后得到處于L4期線蟲,備用。
1.1.3 培養基 PDA培養基:馬鈴薯200 g,葡萄糖20 g,瓊脂16 g,加蒸餾水至1 000 mL,pH自然,121 ℃下滅菌20 min。
PDA液體培養基:馬鈴薯200 g,葡萄糖 20 g,加蒸餾水至1 000 mL,pH自然,121 ℃下滅菌20 min。
1.1.4 主要試劑 1%DMSO、乙酸乙酯、M9緩沖液(KH2PO43 g,Na2HPO46 g、NaCl 5 g, MgSO40.12 g,加蒸餾水至1 000 mL,121 ℃下滅菌20 min)、線蟲裂解液(KOH 2 g,10% NaClO 10 mL,加蒸餾水至100 mL,現配現用),除乙酸乙酯為化學純外其他試劑均為國產分析純。
1.1.5 試驗設備 TG16G離心機(湖南凱達科學儀器有限公司)、KS 4000i Control恒溫搖床(IKA )、SG250H超聲波清洗器(上海冠特超聲儀器有限公司)、R-210旋轉薄膜蒸發儀(BUCHI)、旋渦混合器(海門市其林貝爾儀器制造有限公司)、生化培養箱(上海森信實驗儀器有限公司)、SQP電子天平(賽多利斯科學儀器(北京)有限公司)。
1.2 試驗方法
1.2.1 八角楓內生真菌分離 將八角楓植物材料用清水洗凈,莖切成1~2 cm小段、葉切成 1 cm×1 cm小塊,用酒精(1 min)和0.1%升汞(莖30 s、葉15 s)消毒后用無菌水清洗3次,貼于PDA培養基表面,28 ℃恒溫培養4~5 d,分離、純化單菌落。
1.2.2 發酵液及胞外代謝產物制備 待純化真菌長滿整個平板以后,切一小塊(1 cm ×1 cm)長有菌絲的培養基置于PDA液體培養基中, 180 r·min-1、28 ℃培養72 h,發酵液用4層紗布過濾,取150 mL發酵液,待用。剩余發酵液加入等體積乙酸乙酯萃取胞外代謝產物,上層有機相經旋蒸濃縮,溶質用1% DMSO稀釋,待用;下層水相經旋蒸濃縮,溶質用無菌水稀釋,待用。將以上3部分稀釋溶液分別用針筒過濾器(孔徑 0.22 μm)過濾除菌,置4 ℃冰箱,備用。
1.2.3 樣品制備 將各菌株發酵液稀釋0、2、4、8倍,各取500 μL于24孔板中,平行3組。
各菌株有機相用1% DMSO稀釋至濃度為0.062 5 mg·mL-1、0.125 mg·mL-1、0.25 mg·mL-1、0.5 mg·mL-1,1 mg·mL-1,各取500 μL于24孔板中,平行3組,為樣品組;陰性對照組用1% DMSO處理。水相用無菌水稀釋至濃度為0.062 5 mg·mL-1、0.125 mg·mL-1、0.25 mg·mL-1、0.5 mg·mL-1, 1 mg·mL-1,各取500 μL于24孔板中,平行3組。
1.2.4 殺松材線蟲內生真菌篩選 采用浸漬法[14]測定各菌株代謝產物對松材線蟲的毒殺作用。樣品組和陰性對照組分別加入500 μL濃度為100條·mL-1的松材線蟲,于28 ℃條件下放置12 h、24 h、36 h和48 h,觀察線蟲的存活和死亡數量,并計算死亡率與校正死亡率[15-16]。
死亡率=死蟲數/供試蟲數×100%
校正死亡率=(處理組死亡率-對照組死亡率)/(1-對照組死亡率)×100%
1.3 統計分析
采用Excel 2016軟件整理試驗數據,運用GraphPad Prism 8軟件進行數據處理與單因素方差分析(One-way ANOVA),運用MEGA-X軟件進行ITS序列比對與系統進化樹的構建。
2 結果與分析
2.1 八角楓內生真菌形態學特征
從八角楓莖、葉組織中共分離得到11株內生真菌,編號:BZ-1~BZ-11,其中菌株BZ-8具有較好的殺線蟲活性,其菌落形態特征如圖1。
菌株BZ-8:菌落圓形,正面深褐色,孢子顆粒狀,背面白至淡黃色。發酵液菌絲體球形,表面輻射狀。
2.2 八角楓內生真菌發酵液的殺線蟲活性測定結果
由表1可知,八角楓內生真菌BZ-8發酵液具有明顯的殺松材線蟲活性,且經過稀釋后其活性仍穩定在80%以上。為確定菌株BZ-8發酵液中活性成分性質,分別對有機相和水相的殺蟲活性作進一步測定與分析。

表1 BZ-8發酵液殺松材線蟲毒力測定Table 1 Nematicidal activity of fermentation broth from BZ-8 against PWN
由圖2可知,菌株BZ-8發酵液有機相對松材線蟲具有顯著毒殺作用,濃度為0.25 mg·mL-1樣品經12 h處理后,平均校正死亡率達32.28%;濃度為0.5mg·mL-1樣品經12 h處理后其平均校正死亡率達77.63%;濃度為1.0 mg·mL-1樣品經48 h處理后其平均校正死亡率達 99.15%,具極顯著殺蟲活性。有機相經24 h和48 h處理后的IC50值分別為0.29 mg·mL-1、0.21 mg·mL-1(表2)。
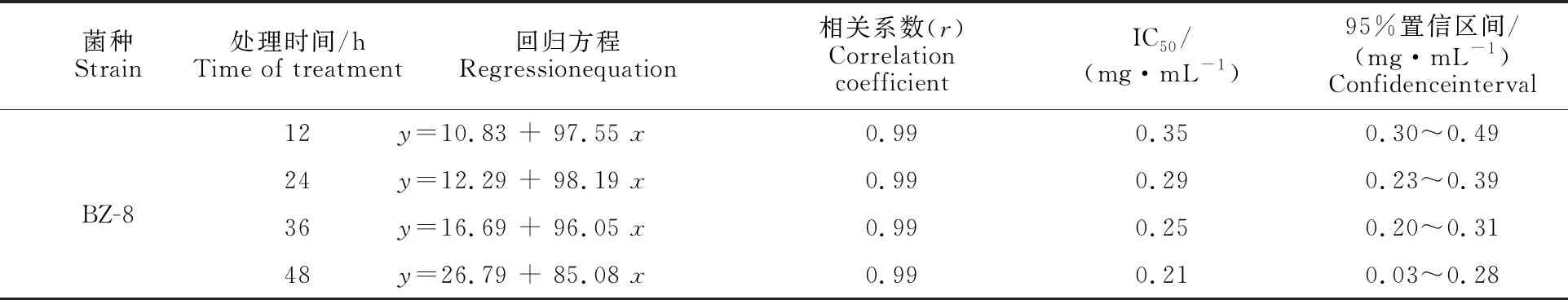
表2 BZ-8胞外發酵產物(有機相)殺松材線蟲毒力測定Table 2 Nematicidal activity of extracellular fermentation products (oganic phase) from BZ-8 against PWN
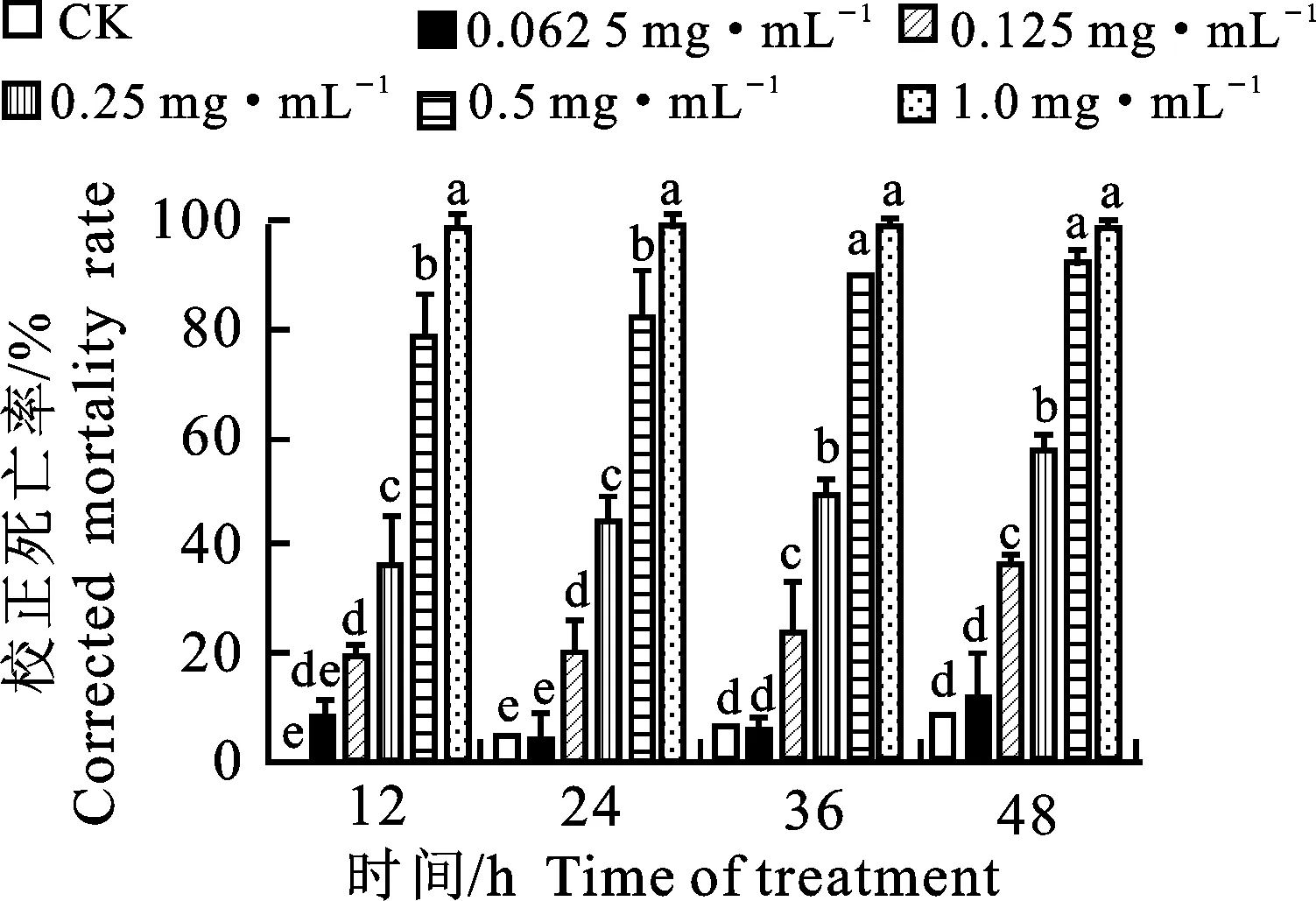
圖中每個時間處理下小寫英文字母不同者表示經Duncan’s法檢驗差異顯著(P<0.05)
由圖3可知,菌株BZ-8水相濃度為0.5 mg·mL-1時,經12 h處理后其殺線蟲活性即可達到95%以上,平均線蟲死亡率為98.92%;濃度為0.062 5 mg·mL-1的水相經12 h處理后其殺蟲率達到60%以上,經48 h處理后平均線蟲死亡率也可達86.34%。由此可見菌株BZ-8發酵液水相在低濃度下仍可保持其良好的殺蟲活性,且經24 h和48 h處理后的IC50值分別為0.04 mg·mL-1、0.02 mg·mL-1(表3),優于有機相的殺蟲活性。
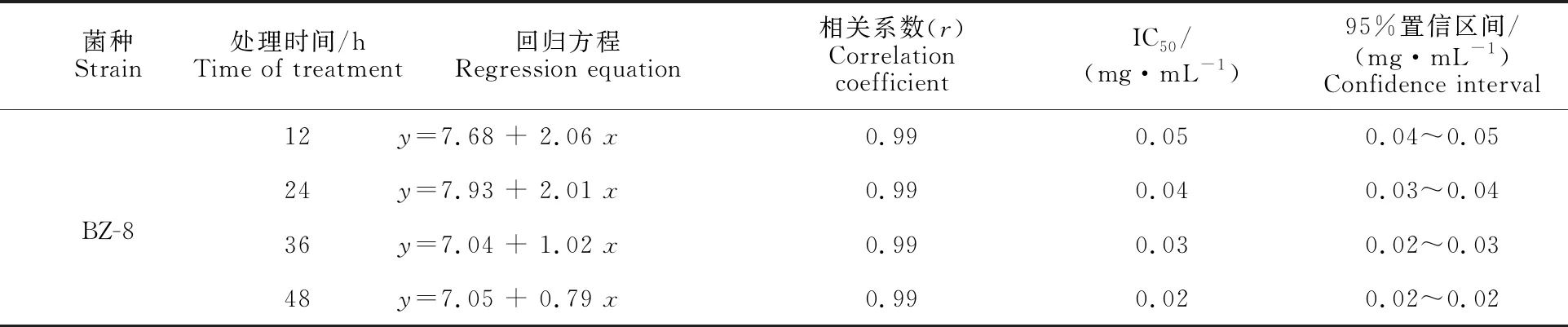
表3 BZ-8胞外發酵產物(水相)殺松材線蟲毒力測定Table 3 Nematicidal activity of extracellular fermentation products (aqueous phase) from BZ-8 against PWN
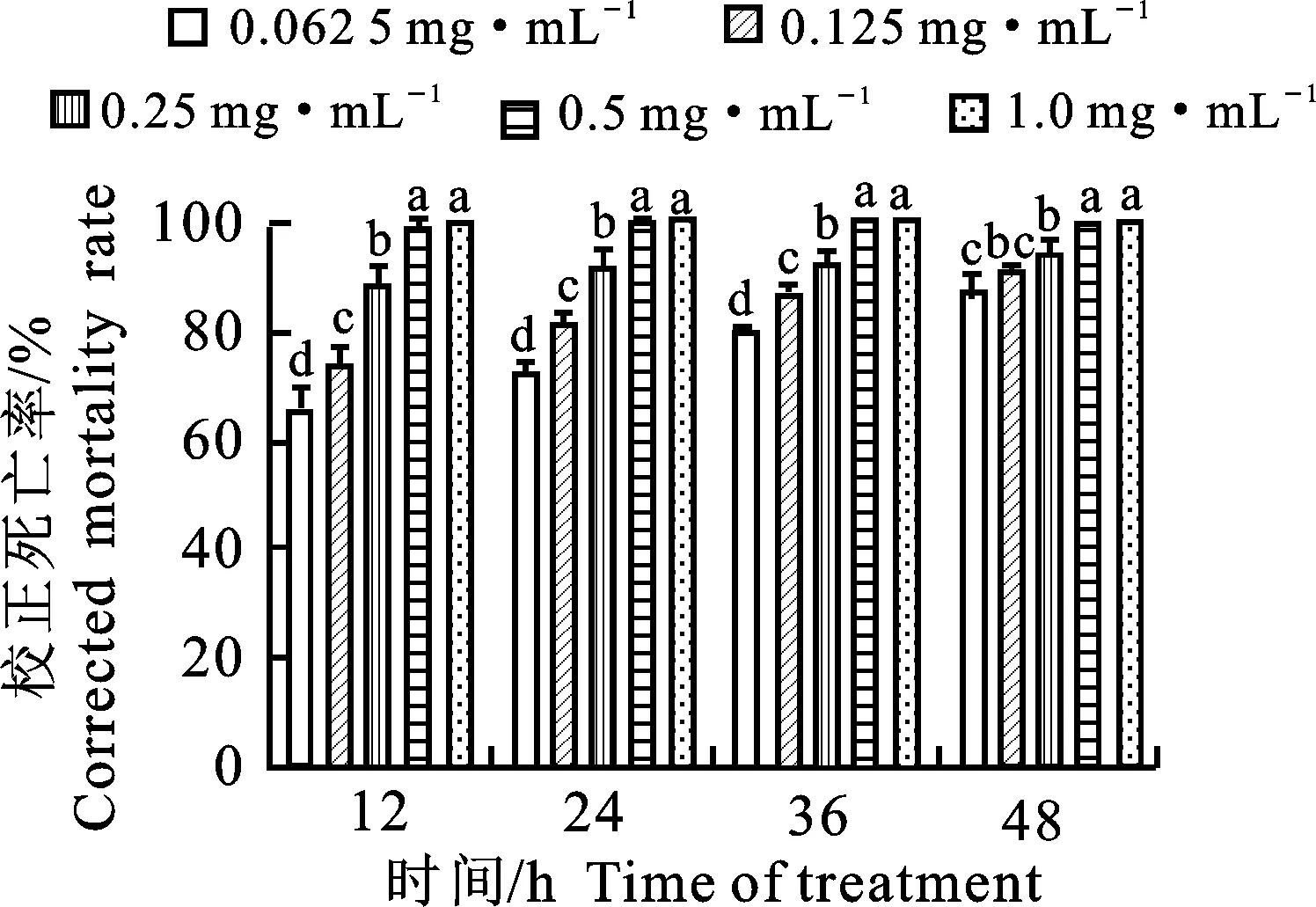
圖中每個時間處理下小寫英文字母不同者表示經Duncan’s法檢驗差異顯著(P<0.05)
2.3 內生真菌的分子生物學鑒定
ITS1引物序列:5′TCCGTAGGTGAACCTGCGG3′;ITS4引物序列:5′TCCTCCGCTTATTGATATGC 3′。PCR體系(50 μL):2×TaqMaster Mix 25 μL,ITS1引物和ITS4引物各 1 μL,gDNA 1 μL,ddH2O 22 μL。
反應程序:預變性95 ℃ 5 min;變性95 ℃ 15 s,退火50 ℃ 20 s,延伸72 ℃ 40 s,循環40次。PCR產物送去上海元莘生物醫藥科技有限公司測序。
2.3.1 ITS序列結果 以TS1和TS4為引物對菌株BZ-8的ITS序列進行擴增,擴增結果見 圖4,擴增產物片段約500 bp[17]。
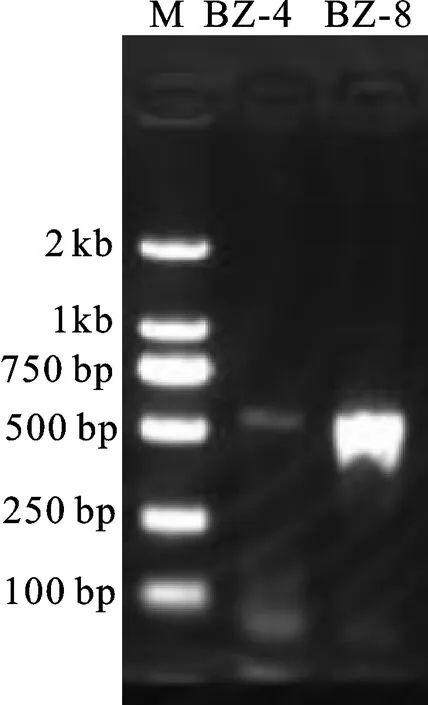
圖4 PCR產物凝膠電泳圖Fig.4 PAGE gels of PCR products
2.3.2 系統進化樹的構建 為確定菌株BZ-8的種類,下載了更多相關序列以進一步確定其種屬關系。圖5表明,菌株BZ8與日本曲霉Aspergillusjaponicus關系非常緊密,各自聚于同一分支[18],自展支持率為97%,結合其性狀特征,鑒定菌株BZ-8為日本曲霉Aspergillusjaponicas。
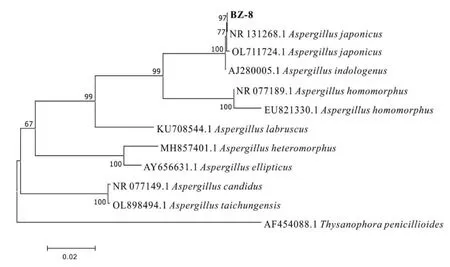
圖5 菌株BZ-8系統進化樹Fig.5 Phylogenetic tree of strain BZ-8
3 討 論
八角楓(A.chinense)作為民族用藥含有酚苷、蒽醌及其苷類、生物堿類等有效成分[19],具有抗菌、抗炎、肌松、心血管系統等生理活性,可用于治療風濕關節疼痛、心力衰竭、跌打損傷等病癥[20],是目前風濕定片、風濕定膠囊、金骨蓮膠囊等制劑的主藥[21]。另外,八角楓也是一種有毒植物[22],八角楓生物堿可阻斷神經肌肉接點的傳導,引起平滑肌松弛,具有呼吸抑制、呼吸肌麻痹和中樞抑制作用[23]。一些八角楓屬植物具有殺蟲作用[24]。
本試驗通過對八角楓枝葉內生真菌的分離純化,得到11株內生真菌,其中一株內生菌(BZ-8)胞外發酵產物具有良好殺線蟲活性,通過分子生物學鑒定為日本曲霉Aspergillusjaponicus。A.japonicus發酵液有機相經24 h處理后,其IC50為0.29 mg·mL-1,殺蟲活性優于甲維鹽而次于阿維菌素;A.japonicus發酵液水相經 24 h處理后其IC50為0.04 mg·mL-1,說明其殺蟲活性成分存在于水相中,為水溶性物質,且活性明顯優于阿維菌素[25]。
何瓊等[26]研究發現,日本曲霉(A.japonicus)發酵液經24 h處理后對2齡南方根結線蟲幼蟲的校正死亡率達到100%,且具有優良穩定性。吳海燕[27]發現A.japonicus發酵濾液5倍稀釋液的殺線蟲率可于6 h內達到100%,且近50%的線蟲蟲體被降解,8 h近90%的線蟲被降解。證實該菌株次生代謝產物具有明顯的殺線蟲活性,可用于線蟲的生物防治,具有作為一種新型殺線劑的開發潛力。
本試驗針對八角楓內生真菌日本曲霉A.japonicus的胞外發酵液毒殺松材線蟲作用進行初步研究,還需要對胞外發酵液的有機相和水相殺蟲活性部位進一步分離純化,以發現活性先導化合物。并對殺蟲活性成分的毒殺松材線蟲的作用機制進行深入研究。
