森林木質藤本數量過度增長的機制與生態效應
2023-06-18 09:50:55宋述靈易伶俐張李龍陳琳余澤平宋慶妮欒豐剛方熊劉駿
廣西植物 2023年5期
宋述靈 易伶俐 張李龍 陳琳 余澤平 宋慶妮 欒豐剛 方熊 劉駿


摘 要: 木質藤本是維持森林物種多樣性的組成部分。近年來,熱帶、亞熱帶森林尤其是次生林內的木質藤本數量的過度增長給森林的恢復和健康發展造成了威脅。至今,國內尚少有研究對森林木質藤本過多的現象、增長機制與生態效應進行綜合認識。該文梳理國內外相關文獻,從木質藤本數量增長的機制與生態效應進行分析和總結,綜合相關研究認為:(1)木質藤本數量的增長與干旱化加劇、大氣CO2濃度上升、自然干擾和森林破碎化有關,在環境變化的情況下,木質藤本在形態、行為、生理等方面比樹木更具優勢,表現為更快的生長速率、更強的繁殖力和可塑性以及資源利用效率。(2)木質藤本主要通過遮蔭脅迫、資源競爭和機械壓力與損傷等方式對樹木造成影響。(3)木質藤本過度增長在個體水平上會阻礙樹木生長、生殖并引起樹木死亡,在群落水平上會改變物種組成、降低多樣性,在生態系統水平上會降低森林碳儲量,改變碳、礦質養分和水分循環過程等。因此,建議結合野外長期監測與控制實驗開展木質藤本數量動態與環境變化關系、森林干擾對木質藤本生長的影響、木質藤本對環境變化的響應及適應機理、木質藤本數量過度增長的生態效應評價研究。同時,應積極探索合理的森林木質藤本管控方法,為森林管理和恢復等方面的宏觀決策提供科學依據。
關鍵詞: 木質藤本, 干擾, 全球變化, 森林健康, 森林管理
中圖分類號: Q948? 文獻標識碼: A? 文章編號: 1000-3142(2023)05-0980-11
Abstract: Lianas are one of the components in maintaining forest diversity. The over-increase of lianas in tropical and subtropical forests, especially in secondary forests is threatening the restoration and healthy development of forests. Few domestic researches make a comprehensive evaluation on the mechanism of the over-increase of lianas and negative effects on forests. In this paper, we reviewed and summarized the related researches on mechanisms and ecological consequences of the over-increase of lianas both at home and abroad. Based on the relevant studies we hold the opinions: (1) The increase of lianas can be corelated with drought severity, elevated atmospheric CO2, increased natural disturbance (gaps) and forest fragments. Lianas possess the advantages of rapid growth, strong reproductive capacity and phenotypic plasticity, and high resource use efficiency under the altered environment. Thus, the favorable environment and inherent advantages can contribute to the increase of lianas. (2) Lianas compete with trees by means of shading stress, resource competition and mechanical loading, abrasion, and strangulation, which all may increase the mortality of trees. (3) The over-increase of lianas can negatively affect the tree growth, reproduction, and survival; at community level, lianas can alter tree community composition and decrease community diversity; at ecosystem level, lianas can ramify forest ecosystem function by decreasing carbon storage and altering carbon, nutrient, and water cycling. Therefore, we recommend that future studies should be aimed at the correlations between population dynamic changes of liana species and environmental changes, the effect of forest disturbance on the growth of lianas, responses of lianas to the environmental changes and the adaption mechanisms, comprehensive evaluation on the ecological consequences of the increase of lianas by means of long-term dynamic monitoring plots and controlling experiments. It is also essential to find appropriate management and control strategies of the over-increase of lianas so as to provide reference for forest management and forest ecological restoration.
Key words: liana, disturbance, global change, forest health, forest management
木質藤本(lianas)是不能單獨直立、通過主莖或借助攀援器官進行生長的植物類群,與樹木相比具有較大的長度和胸徑比(Schnitzer & Bongers, 2002)。木質藤本是森林群落的常見組分,對群落結構和功能的維持具有一定的作用(Schnitzer & Bongers, 2002)。然而,當森林受到嚴重干擾時,木質藤本數量的過度增長給森林健康造成威脅,樹木生長受抑制、群落結構單一化、物種多樣性下降,嚴重影響森林生態系統功能(Schnitzer & Bongers, 2011)。這一問題已引起了國外學者的普遍關注。
國外學者較早關注到木質藤本數量的增長及其負面影響,在木質藤本的分布格局(Schnitzer, 2005)、數量動態(Addo-Fordjour et al., 2021)、監測方法(van der Heijden et al., 2022)、藤與樹的關系(Leonor et al., 2015)、木質藤本增長的機制及生態影響(Schnitzer & Bongers, 2011; de Azevedo Amorim et al., 2018; Marshall et al., 2020; Reis et al., 2020; Meunier et al., 2021a)等方面做了較多的研究。國內學者多關注森林木質藤本的多樣性(Hu et al., 2010;王業社等,2014)、經濟價值(季夢成等,2008)和生態功能(張朝陽等,2007)等積極作用。由于野外辨識和調查的困難,因此在很多調查與研究中往往忽略這一類特殊生活型植物。實際上,木質藤本數量過度增長的現象在我國南方很多森林,尤其是在次生林內較為普遍。例如,在一些自然保護區次生林內,木質藤本甚至形成地毯式覆蓋,嚴重阻礙了森林恢復,使保護區實行的封山育林政策變成了“封山育藤”(圖1)。
目前,國內關于木質藤本對森林影響的研究還較少。關列(1980)較早摘譯報道了人工林受藤本植物的危害及其防治,但當時并未得到較大響應;陳亞軍等(2007)綜述了木質藤本在熱帶森林的生態學功能;王伯蓀等(2009)對某種木質藤本的影響進行了具體的評述。隨后,諸多學者陸續在木質藤本對森林物種多樣性、群落結構及更新的影響(郝建輝等,2011;王忠偉等,2020)、木質藤本與樹木的關系(陸芳等,2021)、藤樹性狀比較(Chen et al., 2021)等方面進行了研究。然而,至今國內尚未見有研究對森林木質藤本過多的現象、增長機制與生態效應進行綜合認識。
為引起國內學者對森林木質藤本過度增長現象及其影響的關注,本文綜合國內外研究進展,重點分析木質藤本過度增長機制,詳細闡述木質藤本增長的潛在生態效應,并提出新的研究方向,以期為木質藤本過度增長的防控及森林管理政策的制定提供參考。
1 木質藤本數量過度增長
木質藤本具有維持森林物種多樣性(Schnitzer & Bongers, 2002)、經濟、藥用和生態功能(張朝陽等,2009;劉揚等,2021)。同時,作為層間植物,木質藤本既是很多動物的生境和通道,又是重要的食物來源(Adams et al., 2016, 2019; Odell et al., 2019)。然而,當木質藤本和樹木比例超出臨界閾值,木質藤本在森林中的積極作用將轉變為負面作用,尤其在次生林或破碎化森林中,木質藤本數量的過度增長將阻礙森林演替與更新(Marshall et al., 2020)。由于外界環境的變化導致木質藤本相對或絕對數量的增長,并給當地森林生態系統造成負面影響,本文稱為木質藤本數量過度增長,簡稱為木質藤本增長。
木質藤本增長的表現形式主要為多度(密度)、斷面積、生物量、豐度和攀爬覆蓋樹木的程度等的增加(Wright et al., 2004; Ingwell et al., 2010; Laurance et al., 2014; Wright et al., 2015)。Phillips等(2002a)首次提出了森林木質藤本增加的現象,通過綜合分析亞馬遜4個區域的47個非破碎化熱帶雨林木質藤本的數量變化,發現20年間大型木質藤本(DBH ≥10 cm)相對斷面積、相對多度和樹木死亡風險顯著增加。此后,眾多研究均得出相似的結果。例如,在尼日利亞一個經歷過火燒的低地熱帶雨林,2005—2014年間攀爬植物的種數由49種增至61種,密度由448~1 152 hm2增至1 712~4 492 hm2(Uwalaka & Muoghalu, 2017);溫帶森林木質藤本也有所增加,在1967—2007年間歐洲溫帶森林林下層木質藤本出現的頻率增加了近39%(Perring et al., 2020)。雖然也有研究結果與此不同(Smith et al., 2017; Bongers et al., 2020; Umaa et al., 2020),但多數研究認為木質藤本的數量呈遞增趨勢。在中國熱帶及亞熱帶區域的很多次生林內木質藤本長勢旺盛、分布廣泛,如熱帶森林的金鐘藤(Merremia boisiana)(王伯蓀等,2009)、亞熱帶森林的鉤藤(Uncaria rhynchophylla)、葛(Pueraria lobata)等在林內形成大面積覆蓋,嚴重影響了當地森林的健康發展(圖1)。
2 木質藤本過度增長的機制
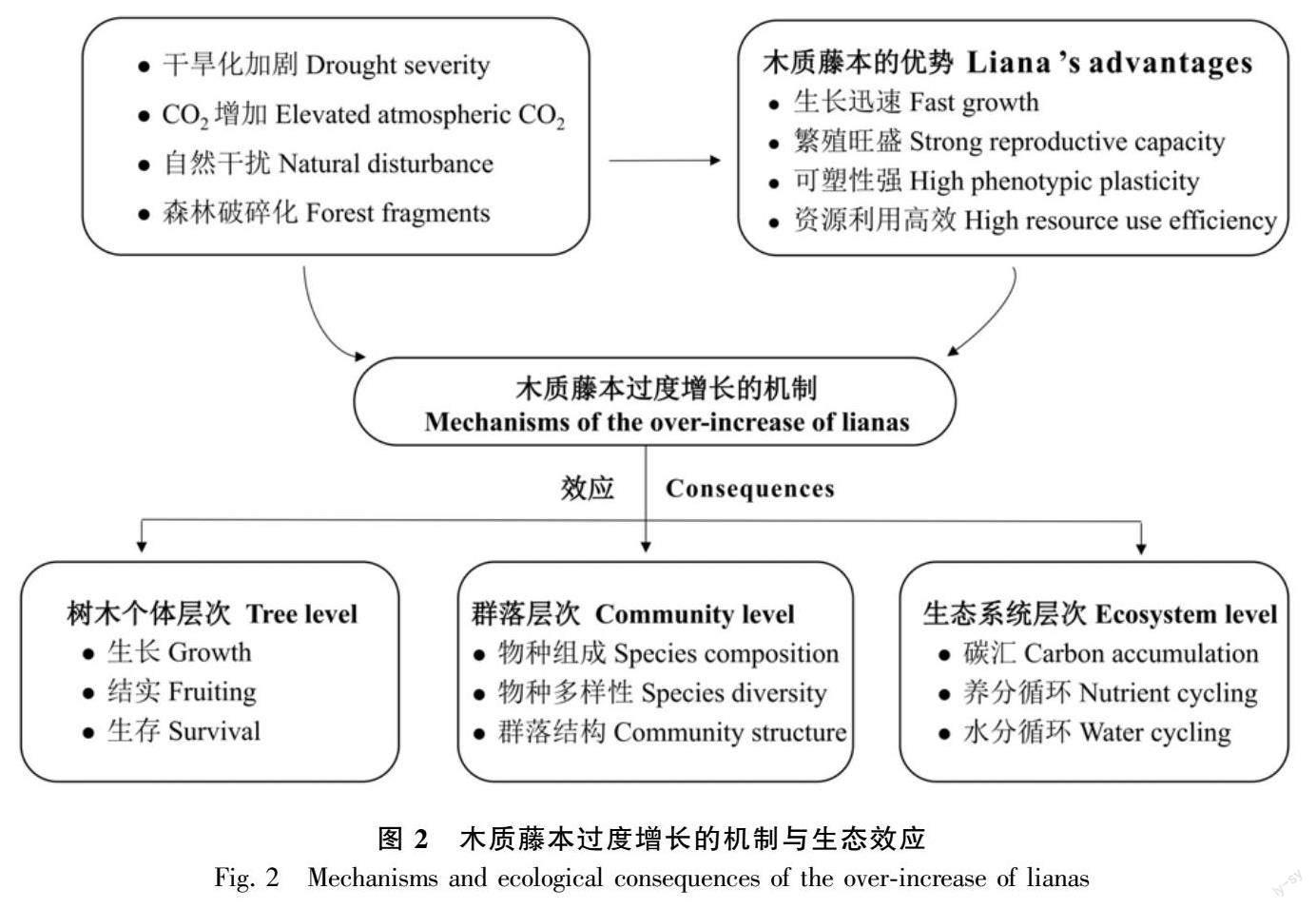
諸多研究認為木質藤本在森林群落中優勢度的增加與外界環境變化有關,主要因素為全球氣候變化、自然干擾(如林窗形成)和森林砍伐等人為干擾導致的森林破碎化、次生化(圖2)。木質藤本能因環境變化做出調整和適應,其生理、結構及行為等特征比樹木更具優勢。
2.1 干旱化加劇
有研究表明木質藤本數量動態與大氣降水量和蒸發量密切相關。木質藤本的多度、豐度和生物量均隨大氣降雨量和土壤濕度的增加而減少,隨干旱程度的加劇和干旱周期的延長而增加(Schnitzer, 2005; Swaine & Grace, 2007)。例如,在巴拿馬干旱森林內,木質藤本密度和豐度的絕對、相對值均高于濕潤森林(Parolari et al., 2020)。
在干旱的條件下,木質藤本比樹木在生長和生理等方面更具優勢。研究發現木質藤本在雨季的生長量比樹木高出2倍,而在旱季高達7倍(Schnitzer & van der Heijden, 2019),木質藤本碳累積量在旱季大于濕季(van der Heijden et al., 2019)。當降雨量減少時,木質藤本幼苗的存活量也高于樹木(Umaa et al., 2019)。此外,在干旱條件下,木質藤本水分利用效率、導水效率和資源獲取策略等均優于樹木(Cai et al., 2009; Zhu & Cao, 2009; Medina-Vega et al., 2021a, b)。例如,熱帶濕潤雨林樹木在水力安全和導水效率之間具有權衡性,而木質藤本卻沒有,較強的導水效率和抗栓塞能力表明木質藤本比樹木更耐旱(van der Sande et al., 2019)。
在干旱條件下,木質藤本各構件還表現出較強的可塑性和適應性。在旱季,木質藤本比樹木具有更強的葉片滲透調節能力,這使其在干旱加劇的環境下更具生長優勢(Maréchaux et al., 2017)。Smith-Martin等(2019)發現在旱季進行灌溉能使樹木的胸徑和生物量顯著增加,而木質藤本并未發生變化,這表明在干旱條件下木質藤本具有更強的適應力。此外,木質藤本具有發達的根系維管系統,可進行營養物質的高效運輸與分配。
木質藤本根系長而錯綜復雜且具有較強的耐脅迫能力,有些甚至能達10 m深(Restom & Nepstad, 2004),可從四周或深處利用不同來源的水分,有效減緩干旱脅迫(Chen et al., 2015)。因此,在全球干旱加劇下,木質藤本的形態、生理和行為特征的優勢可導致其相對和絕對數量的增加。
2.2 大氣CO2濃度增加
持續上升的大氣CO2濃度是木質藤本增長的潛在因素。大氣CO2濃度的增加有利于木質藤本的生長(Granados & Korner, 2002)。當CO2濃度增加時,木質藤本洋常春藤(Hedera helix)新梢長度和生物量的增長量提高了近60%(Zotz et al., 2006)。CO2增加控制實驗也發現溫帶森林木質藤本毒漆藤(Toxicodendron radicans)的生長速率、光合作用、水分利用效率顯著高于正常大氣CO2條件下的狀態(Mohan et al., 2006)。并且,木質藤本對CO2增加的響應比樹木更加迅速(Belote et al., 2004)。木質藤本具有較高的葉莖比和較低的葉片單位面積構建成本,這使其對CO2吸收和固定比樹木更具優勢(Zhu & Cao, 2010)。
2.3 自然干擾
冰雪、火災和樹木死亡等自然(林窗)干擾易促使林內木質藤本的增長。通常情況下,林窗內木質藤本多度和豐度均顯著高于非林窗地帶(Schnitzer & Carson, 2001)。一方面,林窗地帶土壤肥沃、光照較強和空余生態位較多,為木質藤本生長提供了良好的環境。另一方面,很多木質藤本在林窗內具有生長快速和繁殖力強的特征。由于減少了對自身支持結構的投資,木質藤本可將大部分營養資源用于莖的快速生長和葉的累積(Putz, 1984)。除通過種子萌發來進行植株定居外,木質藤本在林窗內也可進行無性克隆繁殖,依靠地下莖、匍匐莖不斷地萌發新枝來向新地段蔓延擴散從而促使植株數量的增加(Schnitzer et al., 2021)。此外,即便是寄主樹木倒塌死亡,很多木質藤本也能夠通過萌發新枝重新攀爬到林冠層,使得木質藤本絕對或相對數量有所增長(Rocha et al., 2020)。
2.4 森林破碎化
森林砍伐、捕獵等人為干擾使森林破碎化、次生化,會導致林緣地帶或次生林內木質藤本增長。森林破碎化后,森林環境變得干燥,土壤肥沃,光照增加,植株矮小,這些條件都有利于木質藤本的攀爬和快速生長。因此,在林緣處和次生林木質藤本的生長速率更快、植株數量更多(Ladwig & Meiners, 2010; Roeder et al., 2019),并且林緣地木質藤本的豐度和碳儲量也高于林內(Londré & Schnitzer, 2006, Magnago et al., 2017; Campbell et al., 2018)。森林破碎化也為木質藤本克隆生長創造了條件,如在拉塞爾瓦擇伐的熱帶雨林,木質藤本長距離克隆生殖對多度增加的貢獻率達到7.5%,斷面積增量高達60%(Yorke et al., 2013)。
3 木質藤本與樹木的競爭方式
3.1 遮蔭脅迫
木質藤本借助莖、枝、葉等器官攀爬到樹木的樹冠,大面積遮蔭導致樹木可獲得光源減少,光合作用受阻,進而影響樹木的生長和發育,嚴重時可致樹木死亡(圖3)。木質藤本在林冠層形成的覆蓋會增加林分郁閉度,從而阻礙林下幼苗更新。在巴拿馬熱帶雨林,木質藤本對樹冠的嚴重覆蓋使樹木死亡率增加了100%(Ingwell et al., 2010);木質藤本使林窗內樹木更新量下降了46%(Schnitzer & Carson, 2010)。木質藤本葉的物候期更長,很多種甚至全年常綠(Putz & Windsor, 1987),常年的遮蔭易使林下幼苗得不到充足的光照而死亡。
3.2 資源競爭
資源的爭奪是木質藤本與樹木的競爭方式,二者對光照、水分和養分等資源存在激烈的競爭(Meunier et al., 2021b)。在清除木質藤本后,樹木的樹干液流速率增加了60%(Leonor et al., 2015)。在礦質養分與水分競爭中,藤與樹的競爭甚至比樹與樹的競爭更加強烈。Tobin等(2012)研究發現樹木的樹干液流速率在清除藤本后增加了約8%,而對樹木的清除并未做出響應。此外,木質藤本具有更高的資源吸收、運輸和利用效率,對資源的爭奪更具競爭力,木質藤本葉單位質量碳同化速率、光能吸收和利用效果均高于樹木(Cai et al., 2009; Asner & Martin, 2012),且木質藤本根系發達(Collins et al., 2016)。例如,當土壤養分增加時,木質藤本葉面積比和光合能力顯著高于樹木(Cai et al., 2008; Pasquini et al., 2015)。
3.3 機械壓力與損傷
木質藤本攀爬會給樹木帶來機械壓力與損傷(Vleut & Pérez-Salicrup, 2005)。木質藤本在樹冠形成大面積覆蓋,給樹冠帶來巨大的載重,使樹梢或樹枝極易折斷,新梢不易萌生。一些木質藤本對樹干的纏繞可導致樹干扭曲、凹陷,增加樹木折斷和病蟲害風險,影響樹木水分和營養物質運輸,最終導致樹木死亡(圖3)。
4 木質藤本過度增長的生態效應
4.1 阻礙樹木生長發育
從個體水平來看,木質藤本增長會從以下三個方面阻礙樹木生長發育。(1)阻礙樹木的正常生長。Finlayson等(2022)研究表明木質藤本會使樹木的生長量降低156%;宋述靈(2019)發現杉木被木質藤本攀爬后連年徑向生長和胸斷面積生長分別降低29%和40%。(2)增加樹木的死亡率。在亞馬遜低地雨林,大樹被木質藤本攀爬后死亡率增加了2倍多(Phillips et al., 2005)。(3)降低樹木的結實率和結實量。在熱帶雨林,樹木結實率與木質藤本的蓋度成顯著負相關(Nabe-Nielsen et al., 2009),木質藤本使堅果類樹種果實產量下降了200%(Kainer et al., 2014)。
木質藤本對幼苗和幼樹的生長也會有負面影響。例如,木質藤本會使樹木幼苗的葉面積減少5倍(Toledo-Aceves & Swaine, 2008);Martínez-Izquierdo等(2016)將14種樹木幼苗分別種于清除和未清除木質藤本的樣地,發現木質藤本使幼苗存活率和株高生長量分別降低了75%和300%;Schnitzer等(2005)將3個不同耐蔭程度的幼樹種植于木質藤本密集的低地雨林,發現2 a后木質藤本使樹木生物量分配格局和樹形均發生變化。由此可見,木質藤本對樹木的影響貫穿了樹木整個生活史。
4.2 改變群落組成與結構
從群落水平來看,木質藤本過度增長會從以下兩個方面改變群落組成與結構。(1)改變群落物種組成。木質藤本對不同樹種影響的差異性易使群落樹種組成發生改變(Visser et al., 2018; Reis et al., 2020)。例如,在加納熱帶常綠落葉混交林內木質藤本使先鋒樹種Nauclea diderichii和Khaya anthotheca的生物量分別下降了32%和50%,而對非先鋒耐蔭樹種Garcinia kola并無影響(Toledo-Aceves & Swaine, 2008)。(2)阻礙群落更新、降低物種多樣性和改變群落結構。木質藤本會降低群落樹種的生殖力,使樹木開花結實的植株數、開花量和結果量顯著下降(García León et al., 2018);Schnitzer和Carson(2010)發現木質藤本使林窗內樹木的更新和多樣性分別下降了46%和65%。木質藤本對不同胸徑樹木影響的差異性也會導致群落結構發生變化(Estrada-Villegas et al., 2020)。
4.3 影響生態系統功能
從生態系統層面來看,木質藤本過度增長會影響森林生態系統碳匯功能。在巴拿馬熱帶雨林,由于受木質藤本的干擾,5 a間樹木生物量的增長量降低22%,據模型預測,30 a后樹木生物量可降低32%,60 a后將達到47%(Lai et al., 2017)。藤本清除實驗驗證了上述模型的合理性,在巴拿馬60 a的熱帶雨林內,木質藤本使地上生物量的年凈累積量下降了76%(van der Heijden et al., 2015)。其他連續監測、模型預測輔助研究和木質藤本清除方式實驗均得出相似結果(di Porcia e Brugnera et al., 2019; van der Heijden et al., 2019; Estrada-Villegas et al., 2020; Meunier et al., 2021a)。雖然木質藤本生物量有所增長,但其增量遠無法補償樹木生物量的損失量,對樹木生物量損失量的彌補不足30%(Schnitzer et al., 2016; Lai et al., 2017)。
木質藤本過度增長可改變森林生態系統水分動態。雖然木質藤本的胸斷面積僅占熱帶森林的5%,但葉蒸騰量占森林總量的12%(Restom & Nepstad, 2001)。此外,木質藤本在水分吸收、運輸和利用等過程比樹木具有優勢(Andrade et al., 2005; Cai et al., 2009; Zhu & Cao, 2010; Chen et al., 2015)。木質藤本也會影響樹木的水分平衡。例如,木質藤本使樹木樹干液流速率下降近一半(Campanello et al., 2016)。
木質藤本過度增長亦能改變森林生態系統的礦質養分動態。較高的葉產量占比(Schnitzer & Bongers, 2002; Kusumoto & Enoki, 2008)和葉片性質與樹木的差異(Cai & Bongers, 2007)能改變森林養分動態和土壤理化性質。熱帶森林木質藤本葉生物量占地上葉總生物量比值高達36%(Tang et al.,2012),凋落物產量和葉面積占比均達到40%(Schnitzer & Bongers, 2002; Tang et al., 2012)。木質藤本的養分含量(如氮、磷)顯著高于樹葉(Asner & Martin, 2012)。因此,木質藤本的過度增長將可能加速森林葉凋落物的分解和養分釋放、改變土壤的養分狀態與呼吸速率、增加土壤養分異質性和養分流失的風險(Putz, 1984; Kusumoto & Enoki, 2008; Liu et al., 2017)。
5 研究展望
雖然越來越多的學者參與到木質藤本對森林影響的研究,但是現階段對木質藤本增長的機制和評價認識還有諸多不足,很多問題尚未解決。結合上述內容,本文認為今后研究可從以下方面開展。
5.1 重視森林動態監測樣地木質藤本定位研究
長期定位監測研究是深入認識種群與森林群落特征變化的重要手段。總體上,木質藤本的定位監測研究起步較晚、研究區域有限、研究尺度較小。在森林動態監測樣地網絡中很多研究單位都未將木質藤本納入監測體系(Schnitzer & Bongers, 2011),現有主要研究地為中美洲和南美洲的巴西、巴拿馬和墨西哥等地的熱帶雨林,非洲少量分布,其他洲較少參與,亞熱帶、溫帶森林關于木質藤本的研究十分缺乏(Schnitzer et al., 2015)。因此,今后森林動態監測樣地的相關研究應將木質藤本納入其中,尤其是要增加亞洲、非洲的研究點,從全球尺度上認識和解析木質藤本的分布格局、動態變化等問題。另外,鑒于木質藤本野外識別和數據采集難度較大,建議采用遙感信息技術開展相關研究(Waite et al., 2019; Chandler et al., 2021)。
5.2 加強環境變化與木質藤本數量動態關系研究
現有研究從氣候變化和干擾的角度對木質藤本的增長原因進行了初步探討,但研究都較為分散、獨立,環境變化導致木質藤本增長的發生機制尚不清楚。建議開展以下研究:(1)各因子的單獨和協同作用對木質藤本動態變化的影響研究,尤其是大時空尺度的大氣CO2變化與木質藤本數量變化的關系研究(Schnitzer & Bongers, 2011);(2)森林干擾使木質藤本增長的發生機制研究,應將干擾狀況、生態因子與木質藤本生物學、生理特性相結合分析木質藤本更新和多樣性維持機制。同時,應加強不同森林干擾類型對木質藤本數量變化影響的比較研究;(3)木質藤本對環境變化的響應研究,對變化環境的適應和調整是木質藤本是否具有競爭優勢的關鍵,應從生理、行為、功能性狀等方面探討木質藤本對環境變化適應機理(Liu et al., 2021),結合控制實驗模擬多種環境因子(如光照、CO2、水分)對木質藤本生長、生理及行為等特征的影響。
5.3 構建木質藤本過度增長的生態效應評價體系
評價木質藤本對森林的影響是制定管理政策的基礎。木質藤本過度增長在個體、種群、群落和生態系統層面均具有一定的負面影響,而相關研究仍然存在不足,對一些領域的深入認識較為缺乏。(1)大時空尺度研究較少。木質藤本的影響與森林類型、年齡等密切相關(Lai et al., 2017; Estrada-Villegas et al., 2021),現有研究多位于熱帶森林,其他區域開展的相關研究不足且長周期的研究較為缺乏。今后應加強多區域、多種森林類型木質藤本的影響評價研究,從大時空尺度認識木質藤本如何影響森林恢復。(2)對木質藤本增加的生態效應評價不夠全面,主要集中在木質藤本對樹木個體的影響,評價指標較為單一,多為樹木胸徑生長,而木質藤本對森林生物量、生態系統物質循環和生產力等方面的影響研究十分缺乏,建議借助模型構建木質藤本對生態系統的影響的全面認識(di Porcia e Brugnera et al., 2019; Meunier et al., 2022)。在我國南方諸多自然保護區次生林內,木質藤本過度增長嚴重阻礙著森林的恢復,建議開展木質藤本過度增長對自然保護區生態系統功能的潛在影響研究。(3)木質藤本與樹木的關系研究通常將木質藤本和樹木作為統一的整體,然而,不同樹種受木質藤本的影響(Visser et al., 2018)、不同木質藤本對樹木施加的壓力都具有種間差異(Muller-Landau et al., 2018),因此難以準確量化出木質藤本對森林的影響。此外,樹木被木質藤本干擾后的響應、環境變化如何影響木質藤本與樹木的關系等方面的研究仍較為缺乏。(4)木質藤本與森林動物的相互關系缺乏關注。動物是森林生態系統的重要組成部分,雖然木質藤本是森林動物的重要食物來源(Odell et al., 2019),可增加動物的多樣性(Schnitzer et al., 2020),但它對樹木的負面影響會降低動物的食物來源從而影響動物群落(Adams et al., 2016; García León et al., 2018)。因此,木質藤本對動物凈影響力有多大;木質藤本的增加分別對哪些種、哪些類型的動物有何影響;動物對木質藤本的取食又如何影響木質藤本數量變化,以及二者間的相互作用關系是否因森林類型、所處區域而具有差異。今后研究都需對這些問題進行系統、定量的評價(Schnitzer, 2018; Coverdale et al., 2021)。
5.4 綜合木質藤本的保護和藤本過度增長的管控
木質藤本過度增長會造成諸多負面影響,而它們亦是森林物種多樣性組成部分,并且部分還屬于珍稀瀕危物種,一些還具有一定的經濟和觀賞價值。由于有的木質藤本與樹木具有互利共生的關系,因而在某些情況下這些木質藤本有助于森林的恢復。大型木質藤本的數量也是評價原始森林健康的指標。因此,今后應立足森林恢復和保護,開展木質藤本保護與管控權衡的研究。在生態環境良好的區域應對木質藤本加以保護,而受影響嚴重的森林應對木質藤本進行適當清除。然而,木質藤本管控的新技術探索將是一大挑戰,當前對木質藤本管理方式呈一刀切的特點,費力耗時、成本高且成效低(Pérez-Salicrup et al., 2001)。在哪里清除、如何清除、如何評價清除效應等問題都需深入研究,從而探索出合理的木質藤本管控方法,為森林管理和生態恢復等方面的宏觀決策提供科學依據。
參考文獻:
ADAMS BJ, SCHNITZER SA, YANOVIAK SP, 2016. Trees as islands: canopy ant species richness increases with the size of liana-free trees in a neotropical forest [J]. Ecography, 40(9): 1067-1075.
ADAMS BJ, SCHNITZER SA, YANOVIAK SP, 2019. Connectivity explains local ant community structure in a neotropical forest canopy: a large-scale experimental approach [J]. Ecology, 100(6): e02673.
ADDO-FORDJOUR PL, AGYEI LA, OFOSU-BAMFO B, et al., 2021. Temporal dynamics of liana communities in moist semi-deciduous forest stands with different management histories in Ghana [J]. For Ecol Manage, 489: 119042.
ANDRADE JL, MEINZER FC, GOLDSTEIN G, et al., 2005. Water uptake and transport in lianas and co-occurring trees of a seasonally dry tropical forest [J]. Trees, 19(3): 282-289.
ASNER GP, MARTIN RE, 2012. Contrasting leaf chemical traits in tropical lianas and trees: implications for future forest composition [J]. Ecol Lett, 15(9): 1001-1007.
BELOTE RT, WELTZIN JF, NORBY RJ, 2004. Response of an understory plant community to elevated [CO2] depends on differential responses of dominant invasive species and is mediated by soil water availability [J]. New Phytol, 161(3): 827-835.
BONGERS F, EWANGO CEN, VAN DER SANDE MT, et al., 2020. Liana species decline in Congo basin contrasts with global patterns [J]. Ecology, 101(5): e03004.
CAI ZQ, BONGERS F, 2007. Contrasting nitrogen and phosphorus resorption efficiencies in trees and lianas from a tropical montane rain forest in Xishuangbanna, south-west China [J]. J Trop Ecol, 23(1): 115-118.
CAI ZQ, POORTER L, HAN Q, et al., 2008. Effects of light and nutrients on seedlings of tropical Bauhinia lianas and trees [J]. Tree Physiol, 28(8): 1277-1285.
CAI ZQ, SCHNITZER SA, BONGERS F, 2009. Seasonal differences in leaf-level physiology give lianas a competitive advantage over trees in a tropical seasonal forest [J]. Oecologia, 161(1): 25-33.
CAMPBELL MJ, EDWARDS W, MAGRACH A, et al., 2018. Edge disturbance drives liana abundance increase and alteration of liana-host tree interactions in tropical forest fragments [J]. Ecol Evol, 8(8): 4237-4251.
CHANDLER CJ, VAN DER HEIJDEN GMF, BOYD DS, et al., 2021. Remote sensing liana infestation in an aseasonal tropical forest: addressing mismatch in spatial units of analyses [J]. Remote Sens Ecol Con, 7(3): 397-410.
CHEN YJ, CHEN JW, CAI ZQ, 2007. Lianas and their functions in tropical forests [J]. Chin Bull Bot, 24(2): 240-249. [陳亞軍, 陳軍文, 蔡志全, 2007. 木質藤本及其在熱帶森林中的生態學功能 [J]. 植物學通報, 24(2): 240-249.]
CHEN YJ, CAO KF, SCHNITZER SA, et al., 2015. Water-use advantage for lianas over trees in tropical seasonal forests [J]. New Phytol, 205(1): 128-136.
CHEN YJ, CHOAT B, STERCK F, et al., 2021. Hydraulic prediction of drought-induced plant dieback and top-kill depends on leaf habit and growth form [J]. Ecol Lett, 24(11): 2350-2363.
COLLINS CG, WRIGHT SJ, WURZBURGER N, 2016. Root and leaf traits reflect distinct resource acquisition strategies in tropical lianas and trees [J]. Oecologia, 180(4): 1037-1047.
COVERDALE TC, OCONNELL RD, HUTCHINSON MC, et al., 2021. Large herbivores suppress liana infestation in an African savanna [J]. Proc Natl Acad Sci USA, 118(41): e2101676118.
DE AZEVEDO AMORIM T, NUNES-FREITAS AF, ROSADO BHP, 2018. Revisiting the hypothesis for increasing liana abundance in seasonal forests: a theoretical review [J]. Plant Soil, 430(1): 1-6.
DI PORCIA E BRUGNERA M, MEUNIER F, LONGO M, et al., 2019. Modeling the impact of liana infestation on the demography and carbon cycle of tropical forests [J]. Glob Change Biol, 25(11): 3767-3780.
ESTRADA-VILLEGAS S, HALL JS, VAN BREUGEL M, et al., 2020. Lianas reduce biomass accumulation in early successional tropical forests [J]. Ecology, 101(5): e02989.
ESTRADA-VILLEGAS S, HALL JS, VAN BREUGEL M, et al., 2021. Lianas do not reduce tree biomass accumulation in young successional tropical dry forests [J]. Oecologia, 195(4): 1019-1029.
FINLAYSON C, ROOPSIND A, GRISCOM BW, et al., 2022. Removing climbers more than doubles tree growth and biomass in degraded tropical forests [J]. Ecol Evol, 12(3): e8758.
GARCA LEN MM, MARTNEZ IZQUIERDO L, MELLO FNA, et al., 2018. Lianas reduce community-level canopy tree reproduction in a Panamanian forest [J]. J Ecol, 106(2): 737-745.
GRANADOS J, KORNER C, 2002. In deep shade, elevated CO2 increases the vigor of tropical climbing plants [J]. Glob Change Biol, 8(11): 1109-1117.
GUAN L, 1980. Damage of vines in forest plantations and of their control and management [J]. J Jilin For Sci Technol, (4): 120-121. [關列, 1980. 人工林受藤本植物的危害及其防治 [J]. 吉林林業科技, (4): 120-121.]
GOLDSTEIN G, SANTIAGO LS, 2016. Tropical tree physiology [M]// CAMPANELLO PI, MANZAN E, VILLAGRA M, et al. Carbon allocation and water relations of lianas versus trees. Switzerland: Springer: 103-124.
HAO JH, TAO JP, ZHAO K, et al., 2011. The effects of lianas on community structure and regeneration of Vatica astrotricha plantations in Bawangling Nature Reserve, Hainan Province, China [J]. J SW Univ (Nat Sci Ed), 33(2): 34-39. [郝建輝, 陶建平, 趙科, 等, 2011. 木質藤本對人工林群落結構及更新的影響 [J]. 西南大學學報(自然科學版), 33(2): 34-39.]
HU L, LI MG, LI Z, 2010. Geographical and environmental gradients of lianas and vines in China [J]. Global Ecol Biogeogr, 19(4): 554-561.
INGWELL LL, WRIGHT SJ, BECKLUND KK, et al., 2010. The impact of lianas on 10 years of tree growth and mortality on Barro Colorado Island, Panama [J]. J Ecol, 98(4): 879-887.
JI MC, ZHENG G, YU SG, 2008. Resources and landscape application of wild vine in Longwangshan National Nature Reserve, Zhejiang Province [J]. Sci Silv Sin, 44(9): 165-169. [季夢成, 鄭鋼, 俞曙光, 2008. 浙江龍王山自然保護區藤本植物資源及園林應用 [J]. 林業科學, 44(9): 165-169.]
KAINER KA, WADT LHO, STAUDHAMMER CL, 2014. Testing a silvicultural recommendation: Brazil nut responses 10 years after liana cutting [J]. J Appl Ecol, 51(3): 655-663.
KUSUMOTO B, ENOKI T, 2008. Contribution of a liana species, Mucuna macrocarpa Wall., to litterfall production and nitrogen input in a subtropical evergreen broad-leaved forest [J]. J For Res, 13(1): 35-42.
LADWIG LM, MEINERS SJ, 2010. Spatiotemporal dynamics of lianas during 50 years of succession to temperate forest [J]. Ecology, 91(3): 671-680.
LAI HR, HALL JS, TURNER BL, et al., 2017. Liana effects on biomass dynamics strengthen during secondary forest succession [J]. Ecology, 98(4): 1062-1070.
LAURANCE WF, ANDRADE AS, AINHOA M, et al., 2014. Long-term changes in liana abundance and forest dynamics in undisturbed Amazonian forests [J]. Ecology, 95(6): 1604-1611.
LEONOR AC, SCHNITZER SA, REID JP, et al., 2015. Liana competition with tropical trees varies seasonally but not with tree species identity [J]. Ecology, 96(1): 39-45.
LIU Q, STERCK FJ, ZHANG JL, et al., 2021. Traits, strategies, and niches of liana species in a tropical seasonal rainforest [J]. Oecologia, 196(2): 499-514.
LIU Y, ZHANG N, LUO J, et al., 2021. Chemical constituents of stems and leaves from Uncaria rhynchophylla in Guizhou and their antitumor activities [J]. Guihaia, 41(7): 1061-1069. [劉揚, 張妮, 羅俊, 等, 2021. 黔產鉤藤莖、葉化學成分及抗腫瘤活性研究 [J]. 廣西植物, 41(7): 1061-1069.]
LIU YC, SHANG Q, ZHANG B, et al., 2017. Effects of understory liana Trachelospermum jasminoides on distributions of litterfall and soil organic carbon in an oak forest in central China [J]. Sustainability, 9(6): 1019.
LONDR RA, SCHNITZER SA, 2006. The distribution of lianas and their change in abundance in temperate forests over the past 45 years [J]. Ecology, 87(12): 2973-2978.
LU F, LI XK, WANG B, et al., 2021. Spatial pattern of lianas of Litsea dilleniifolia community and its relationship with main tree species in Nonggang, Guangxi [J]. Acta Ecol Sin, 41(15): 6191-6202. [陸芳, 李先琨, 王斌, 等, 2021. 廣西弄崗五椏果葉木姜子群落木質藤本空間格局及其與樹種間的關系 [J]. 生態學報, 41(15): 6191-6202.]
MAGNAGO LFS, MAGRACH A, BARLOW J, et al., 2017. Do fragment size and edge effects predict carbon stocks in trees and lianas in tropical forests? [J]. Funct Ecol, 31(2): 542-552.
MARCHAUX I, BARTLETT MK, IRIBAR A, et al., 2017. Stronger seasonal adjustment in leaf turgor loss point in lianas than trees in an Amazonian forest [J]. Biol Lett, 13(1): 20160819.
MARSHALL AR, PLATTS PJ, CHAZDON RL, et al., 2020. Conceptualising the global forest response to liana proliferation [J]. Front For Glob Change, 3: 35.
MARTNEZ-IZQUIERDO L, GARCA MM, POWERS JS, et al., 2016. Lianas suppress seedling growth and survival of 14 tree species in a Panamanian tropical forest [J]. Ecology, 97(1): 215-224.
MEDINA-VEGA JA, BONGERS F, POORTER L, et al., 2021a. Lianas have more acquisitive traits than trees in a dry but not in a wet forest [J]. J Ecol, 109(6): 2367-2384.
MEDINA-VEGA JA, BONGERS F, SCHNITZER SA, et al., 2021b. Lianas explore the forest canopy more effectively than trees under drier conditions [J]. Funct Ecol, 35(2): 318-329.
MEUNIER F, VAN DER HEIJDEN GMF, SCHNITZER SA, et al., 2021a. Lianas significantly reduce aboveground and belowground carbon storage: a virtual removal experiment [J]. Front For Glob Change, 4: 663291.
MEUNIER F, VERBEECK H, COWDERY B, et al., 2021b. Unraveling the relative role of light and water competition between lianas and trees in tropical forests: a vegetation model analysis [J]. J Ecol, 109(1): 519-540.
MEUNIER F, VISSER MD, SHIKLOMANOV A, et al., 2022. Liana optical traits increase tropical forest albedo and reduce ecosystem productivity [J]. Glob Change Biol, 28(1): 227-244.
MOHAN JE, ZISKA LH, SCHLESINGER WH, et al., 2006. Biomass and toxicity responses of poison ivy (Toxicodendron radicans) to elevated atmospheric CO2 [J]. Proc Natl Acad Sci USA, 103(24): 9086-9089.
MULLER-LANDAU HC, VISSER MD, GODOY O, 2018. How do lianas and vines influence competitive differences and niche differences among tree species? Concepts and a case study in a tropical forest [J]. J Ecol, 107(3): 1469-1481.
NABE-NIELSEN J, KOLLMANN J, PEA-CLAROS M, 2009. Effects of liana load, tree diameter and distances between conspecifics on seed production in tropical timber trees [J]. For Ecol Manage, 257(3): 987-993.
ODELL EH, STORK NE, KITCHING RL, 2019. Lianas as a food resource for herbivorous insects: a comparison with trees [J]. Biol Rev, 94(4): 1416-1429.
PAROLARI AJ, PAUL K, GRIFFING A, et al., 2020. Liana abundance and diversity increase with rainfall seasonality along a precipitation gradient in Panama [J]. Ecography, 43(1): 25-33.
PASQUINI SC, WRIGHT SJ, SANTIAGO LS, 2015. Lianas always outperform tree seedlings regardless of soil nutrients: results from a long-term fertilization experiment [J]. Ecology, 96(7): 1866-1876.
PREZ-SALICRUP DR, CLAROS A, GUZMN R, et al., 2001. Cost and efficiency of cutting lianas in a lowland liana forest of Bolivia [J]. Biotropica, 33(2): 324-329.
PERRING MP, DE FRENNE P, HERTZOG LR, et al., 2020. Increasing liana frequency in temperate European forest understories is driven by ivy [J]. Front Ecol Evol, 18(10): 550-557.
PHILLIPS OL, VSQUEZ MARTNEZ R, ARROYO L, et al., 2002. Increasing dominance of large lianas in Amazonian forests [J]. Nature, 418(15): 770-774.
PHILLIPS OL, VSQUEZ MARTNEZ R, MONTEAGUDO MENDOZA A, et al., 2005. Large lianas as hyperdynamic elements of the tropical forest canopy [J]. Ecology, 86(5): 1250-1258.
PUTZ FE, 1984. The natural history of lianas on Barro Colorado Island, Panama [J]. Ecology, 65(6): 1713-1724.
PUTZ FE, WINDSOR DM, 1987. Liana phenology on Barro Colorado Island, Panama [J]. Biotropica, 19(4): 334-341.
REIS SM, MARIMON BS, MORANDI PS, et al., 2020. Causes and consequences of liana infestation in southern Amazonia [J]. J Ecol, 108(6): 2184-2197.
RESTOM TG, NEPSTAD DC, 2001. Contribution of vines to the evapotranspiration of a secondary forest in eastern Amazonia [J]. Plant Soil, 236(2): 155-163.
RESTOM TG, NEPSTAD DC, 2004. Seedling growth dynamics of a deeply rooting liana in a secondary forest in eastern Amazonia [J]. For Ecol Manage, 190(1): 109-118.
ROCHA EX, SCHIETTI J, GEROLAMO CS, et al., 2020. Higher rates of liana regeneration after canopy fall drives species abundance patterns in central Amazonia [J]. J Ecol, 108(4): 1311-1321.
ROEDER M, LIU JJ, PASION B, et al., 2019. Wood density, growth and mortality relationships of lianas on environmental gradients in fragmented forests of montane landscapes [J]. J Veg Sci, 30(6): 1143-1152.
SCHNITZER SA, 2005. A mechanistic explanation for global patterns of liana abundance and distribution [J]. Am Nat, 166(2): 262-276.
SCHNITZER SA, 2018. Testing ecological theory with lianas [J]. New Phytol, 220(2): 366-380.
SCHNITZER SA, BONGERS F, 2002. The ecology of lianas and their role in forests [J]. Trends Ecol Evol, 17(5): 223-230.
SCHNITZER SA, BONGERS F, 2011. Increasing liana abundance and biomass in tropical forests: emerging patterns and putative mechanisms [J]. Ecol Lett, 14(4): 397-406.
SCHNITZER SA, BONGERS F, BURNHAM RJ, et al., 2015. Ecology of lianas [M]//DEWALT SJ, SCHNITZER SA, ALVES LF, et al. Biogeographical pattern of liana abundance and diversity. Oxford: Wiley-Blackwell: 132-143.
SCHNITZER SA, CARSON WP, 2001. Treefall gaps and the maintenance of species diversity in a tropical forest [J]. Ecology, 82(4): 913-919.
SCHNITZER SA, CARSON WP, 2010. Lianas suppress tree regeneration and diversity in treefall gaps [J]. Ecol Lett, 13(7): 849-857.
SCHNITZER SA, DEFILIPPIS DM, VISSER M, et al., 2021. Local canopy disturbance as an explanation for long-term increases in liana abundance [J]. Ecol Lett, 24(12): 2635-2647.
SCHNITZER SA, GEERTJE VDH, MASCARO J, et al., 2016. Lianas in gaps reduce carbon accumulation in a tropical forest [J]. Ecology, 95(11): 3008-3017.
SCHNITZER SA, MICHEL NL, POWERS JS, et al., 2020. Lianas maintain insectivorous bird abundance and diversity in a neotropical forest [J]. Ecology, 101(12): e03176.
SCHNITZER SA, KUZEE ME, BON GERS F, 2005. Disentangling above- and below-ground competition between lianas and trees in a tropical forest [J]. J Ecol, 93(6): 1115-1125.
SCHNITZER SA, VAN DER HEIJDEN GMF, 2019. Lianas have a seasonal growth advantage over co-occurring trees [J]. Ecology, 100(5): e02655.
SMITH-MARTIN CM, BASTOS CL, LOPEZ OR, et al., 2019. Effects of dry-season irrigation on leaf physiology and biomass allocation in tropical lianas and trees [J]. Ecology, 100(11): e02827.
SMITH JR, QUEENBOROUGH SA, ALVIA P, et al., 2017. No strong evidence for increasing liana abundance in the Myristicaceae of aneotropical aseasonal rain forest [J]. Ecology, 98(2): 456-466.
SONG SL, 2019. Liana diversity and its effects on tree growth and community structure in Guanshan evergreen broad-leaved forest [D]. Nanchang: Jiangxi Agricultural University. [宋述靈, 2019. 官山大樣地木質藤本對樹木生長與群落結構的影響 [D]. 南昌: 江西農業大學.]
SWAINE MD, GRACE J, 2007. Lianas may be favoured by low rainfall: evidence from Ghana [J]. Plant Ecol, 192(2): 271-276.
TANG Y, KITCHING RL, CAO M, 2012. Lianas as structural parasites: a re-evaluation[J]. Chin Sci Bull, 57(4): 307-312.
TOBIN MF, WRIGHT AJ, MANGAN SA, et al., 2012. Lianas have a greater competitive effect than trees of similar biomass on tropical canopy trees [J]. Ecosphere, 3(2): 1-11.
TOLEDO-ACEVES T, SWAINE MD, 2008. Effect of lianas on tree regeneration in gaps and forest understorey in a tropical forest in Ghana [J]. J Veg Sci, 19(5): 717-728.
UMAA MN, FORERO-MONTAA J, NYTCH CJ, et al., 2019. Dry conditions and disturbance promote liana seedling survival and abundance [J]. Ecology, 100(1): e02556.
UMAA MN, MANZAN-PINZN E, COMITA LS, 2020. Long-term dynamics of liana seedlings suggest decelerating increases in liana relative abundance over time [J]. J Ecol, 108(2): 460-469.
UWALAKA NO, MUOGHALU JI, 2017. Nine years of climber community dynamics in a Nigerian lowland rain forest 31 years after a ground fire [J]. Biodivers Conserv, 26(4): 997-1008.
VAN DER HEIJDEN GMF, POWERS JS, SCHNITZER SA, 2015. Lianas reduce carbon accumulation and storage in tropical forests [J]. Proc Natl Acad Sci USA, 112(43): 13267-13271.
VAN DER HEIJDEN GMF, POWERS JS, SCHNITZER SA, 2019. Effect of lianas on forest-level tree carbon accumulation does not differ between seasons: results from a liana removal experiment in Panama [J]. J Ecol, 107(4): 1890-1900.
VAN DER HEIJDEN GMF, PROCTOR ADC, CALDERS K, et al., 2022. Making (remote) sense of lianas [J]. J Ecol, 110(3): 498-513.
VAN DER SANDE MT, POORTER L, SCHNITZER SA, et al., 2019. The hydraulic efficiency-safety trade-off differs between lianas and trees [J]. Ecology, 100(5): e02666.
VISSER MD, MULLER-LANDAU HC, SCHNITZER SA, et al., 2018. A host-parasite model explains variation in liana infestation among co-occurring tree species [J]. J Ecol, 106(6): 2435-2445.
VLEUT I, PREZ-SALICRUP DR, 2005. Lianas and their supporting plants in the understorey at Los Tuxtlas, Mexico [J]. J Trop Ecol, 21(5): 577-580.
WAITE CE, VAN DER HEIJDEN GMF, FIELD R, et al., 2019. A view from above: unmanned aerial vehicles (UAVs) provide a new tool for assessing liana infestation in tropical forest canopies [J]. J Appl Ecol, 56(4): 902-912.
WANG YS, YANG XJ, CHEN LJ, et al., 2014. Biodiversity of wild woody vine resources in Yunshan, Hunan [J]. Acta Bot Boreal-Occident Sin, 34(3): 604-613. [王業社, 楊賢均, 陳立軍, 等, 2014. 湖南云山野生木質藤本植物資源及多樣性研究 [J]. 西北植物學報, 34(3): 604-613.]
WANG ZW, ZHANG KM, WU N, et al., 2020. Effect of liana on the understory species richness in deciduous broadleaf forest of Jigong Mountain in Henan [J]. J Yunnan Agric Univ (Nat Sci Ed), 35(3): 476-482. [王忠偉, 張科萌, 鄔娜, 等, 2020. 河南雞公山落葉闊葉林藤本植物對林下物種多樣性的影響 [J]. 云南農業大學學報(自然科學版), 35(3): 476-482.]
WANG BS, PENG SL, LI DJ, et al., 2009. Research progress on Merremia boisiana [J]. Chin J Ecol, 28(11): 2360-2365. [王伯蓀, 彭少麟, 李代江, 等, 2009. 金鐘藤研究述評 [J]. 生態學雜志, 28(11): 2360-2365.]
WRIGHT SJ, CALDERN O, HERNANDZ A, et al., 2004. Are lianas increasing in importance in tropical forest?A 17-year record from Panama [J]. Ecology, 85(2): 484-489.
WRIGHT SJ, SUN IF, PICKERING M, et al., 2016. Long-term changes in liana loads and tree dynamics in a Malaysian forest [J]. Ecology, 96(10): 2748-2757.
YORKE SR, SCHNITZER SA, MASCARO J, et al., 2013. Increasing liana abundance and basal area in a tropical forest: the contribution of long-distance clonal colonization[J]. Biotropica, 45(3): 317-324.
ZHANG ZY, ZHOU FX, XU GF, 2009. Application of liana in ecological restoration of side slope [J]. Res Soil Water Conserv, 16(3): 291-293. [張朝陽, 周鳳霞, 許桂芳, 2009. 藤本植物在邊坡生態恢復中的應用 [J]. 水土保持研究, 16(3): 291-293.]
ZHU SD, CAO KF, 2009. Hydraulic properties and photosynthetic rates in co-occurring lianas and trees in a seasonal tropical rainforest in southwestern China [J]. Plant Ecol, 204(2): 295-304.
ZHU SD, CAO KF, 2010. Contrasting cost-benefit strategy between lianas and trees in a tropical seasonal rain forest in southwestern China [J]. Oecologia, 163(3): 591-599.
ZOTZ G, CUENI N, KOERNER C, 2006. In situ growth stimulation of a temperate zone liana (Hedera helix) in elevated CO2 [J]. Funct Ecol, 20(5): 763-769.
(責任編輯 李 莉 王登惠)
