亞熱帶八個闊葉林內(nèi)不同分布型植物的性狀和生態(tài)習(xí)性差異
2023-06-18 19:23:27蔣婭毛艷嬌鄭潔車儉張潮金毅
廣西植物 2023年5期
蔣婭 毛艷嬌 鄭潔 車儉 張潮 金毅

摘 要: 組成我國亞熱帶地區(qū)植被的植物區(qū)系來源多樣。為了解共同生活在該地區(qū)的不同區(qū)系成分間在生態(tài)位上存在何種差異,以提高對該地區(qū)生物多樣性維持機(jī)制的認(rèn)識,該研究以中國亞熱帶地區(qū)8個闊葉林動態(tài)樣地內(nèi)木本被子植物群落為研究對象,通過采用譜系線性回歸和方差分析(ANOVA)等統(tǒng)計方法,分析了溫帶和熱帶分布型屬植物在生活史性狀(葉習(xí)性和生長型)和生態(tài)習(xí)性(Ellenberg生態(tài)指示值)上的差異。結(jié)果表明:(1)對于涉及的265個屬,分布型屬具有一定的譜系保守性(Pagels λ=0.935, P<0.001)。(2)溫帶分布型屬含落葉闊葉植物和喬木較多,熱帶分布型屬則以常綠闊葉植物和灌木為主。(3)與熱帶分布型屬植物相比,溫帶分布型屬植物所處環(huán)境具有較強(qiáng)光照、較低溫度和濕度,且在控制了葉習(xí)性和生長型的影響后,兩者間的生態(tài)習(xí)性差異仍然存在。(4)對于所有植物,或者對于不同葉習(xí)性和生長型功能群而言,溫帶與熱帶分布型屬植物的相對優(yōu)勢(物種數(shù)的比值)均受到年平均氣溫的影響。該研究結(jié)果有助于理解區(qū)系來源對亞熱帶闊葉林內(nèi)木本被子植物的生活史性狀和生態(tài)習(xí)性均具有重要影響,以及不同區(qū)系來源的植物在我國亞熱帶闊葉林內(nèi)的地理更替受到年平均氣溫的驅(qū)動。
關(guān)鍵詞: 分布型, 生態(tài)位, 生態(tài)指示值, 亞熱帶森林, 中國森林生物多樣性監(jiān)測網(wǎng)絡(luò)
中圖分類號: Q948? 文獻(xiàn)標(biāo)識碼: A? 文章編號: 1000-3142(2023)05-0890-10
Abstract: The plants that comprise the vegetation of Chinese subtropics are diverse with respect to geographical distribution patterns. Understanding how the plants with different geographical distribution patterns that live together in this region differ in their ecological niches, would advance our knowledge of species coexistence mechanisms. Here, based on the data of woody angiosperm community composition in eight broad-leaved forest dynamics plots widely distributed in the Chinese subtropics, by adopting statistical methods such as phylogenetic linear model and analysis of variance (ANOVA), we examined the differences in life history traits, including leaf habit and growth form, as well as ecological behaviors (i.e., Ellenberg indicator values), between the plants of temperate distribution pattern genera and the plants of tropical distribution pattern genera. The results were as follows: (1) For the 265 study genera, distribution pattern genus showed a significant phylogenetic signal (i.e., Pagels λ = 0.935, P< 0.001). (2) The temperate distribution pattern genera comprised a higher ratio of deciduous broad-leaved plants and trees than the tropical distribution pattern genera, and there were more evergreen broad-leaved plants and shrub in tropical distribution pattern. (3) The plants of temperate distribution pattern genera exist in environments with high light, low temperature and water compared with the plants of tropical distribution pattern genera, regardless of the effects of leaf habit and growth form were controlled or not. (4) For all the plants, or for each leaf habit or growth form group separately, the ratio of species richness of the temperate distribution pattern genera to the tropical distribution pattern genera in the forest was controlled by mean annual temperature (MAT). The findings imply that the plants of temperate and tropical distribution pattern genera differe in both their life history traits as well as their ecological behaviors, and the geographical turnover of plants of these two distribution patterns in the Chinese subtropical broad-leaved forests is largely controlled by MAT.
Key words: distribution pattern, niche, ecological indicatior value, subtropical forest, Chinese Forest Biodiversity Monitoring Network
植物區(qū)系成分研究中,分析區(qū)域性植物的地理成分組成,即溫帶性質(zhì)和熱帶性質(zhì)的分布型(distribution pattern)等植物組成及比例(王荷生, 1992)。吳征鎰等(2010)研究認(rèn)為區(qū)系成分組成可反映區(qū)域性植物與氣候和土壤等環(huán)境條件的關(guān)系。不同分布型植物的比例變化,綜合反映了區(qū)域性植物與環(huán)境關(guān)系的協(xié)同變化(王荷生, 1992)。一方面,分布型可反映植物類群在演化史上的分異情況和地理特征(吳征鎰等, 2010),相同分布型植物傾向于具有相似的生活史性狀,例如葉習(xí)性和生長型。葉習(xí)性中,落葉性狀有利于植物渡過高緯度地帶的季節(jié)性低溫等氣候時期,而常綠性狀則有助于適應(yīng)低緯度地區(qū)季節(jié)性差異小的氣候(Givnish, 2002)。由于冬季較長時間的低溫冰凍等,溫帶分布型植物可能多為冬季落葉植物(Givnish, 1979; Zanne et al., 2014)。此外,生長型是植物長期適應(yīng)氣候等自然條件所形成的總體形態(tài)特征(王荷生, 1992)。不同生長型植物對于氣候等環(huán)境條件的響應(yīng)存在差異(Xu et al., 2018; Zhao et al., 2018; Xu et al., 2021)。另一方面,不同分布型植物對區(qū)域尺度氣候條件的響應(yīng)也存在差異(Qian et al., 2003, 2006; Zhu et al., 2007; Wang et al., 2011; Xu et al., 2013; Su et al., 2020),提示分布型在一定程度上也反映了植物的生態(tài)習(xí)性(ecological behavior)特點(diǎn)。例如,由于溫帶分布區(qū)氣溫通常較低,溫帶分布型植物可能對溫度的要求較低(Su et al., 2020)。生態(tài)習(xí)性通常是多維度的,綜合反映植物在自然狀態(tài)下的生態(tài)需求(宋永昌, 2013)。例如,Ellenberg(1979)采用光照、溫度、大陸度、土壤濕度、土壤反應(yīng)和土壤含氮量這6個指標(biāo)來劃分中歐維管束植物的生態(tài)習(xí)性。宋永昌(2013)使用光照、溫度、濕度、土壤反應(yīng)和土壤肥沃度來劃分我國常綠闊葉林內(nèi)4 000余種維管束植物的生態(tài)習(xí)性。總之,以往研究顯示分布型與生活史性狀或生態(tài)習(xí)性均有密切關(guān)系。但是,還缺乏對于不同分布型植物的生活史性狀和生態(tài)習(xí)性差異,以及不同分布型植物在地理分布上發(fā)生更替的主要環(huán)境驅(qū)動因子的綜合分析。
探討群落內(nèi)不同分布型植物的生活史性狀和生態(tài)習(xí)性差異,以及驅(qū)動不同分布型植物相對優(yōu)勢變化的主要環(huán)境因子,可以進(jìn)一步豐富區(qū)系成分的意義(孫航等, 2017)。我國亞熱帶地區(qū)植被具有古老的歷史和豐富的植物多樣性(吳征鎰, 1965, 1980; Lu et al., 2018; Qian et al., 2019),是探討不同分布型植物的生活史性狀和生態(tài)習(xí)性差異以及相對優(yōu)勢驅(qū)動因子的理想地區(qū)。據(jù)此,本研究以我國亞熱帶地區(qū)為研究區(qū)域,依托廣泛分布的8個闊葉林動態(tài)監(jiān)測樣地(forest dynamics plot, FDP)內(nèi)木本被子植物群落(宋永昌等, 2015),采用譜系線性回歸和方差分析等統(tǒng)計方法,通過探討不同分布型植物的生活史性狀(葉習(xí)性和生長型)和生態(tài)習(xí)性的具體差異,以及驅(qū)動不同分布型植物在群落內(nèi)相對優(yōu)勢發(fā)生變化的環(huán)境因子,擬回答以下問題:(1)亞熱帶闊葉林內(nèi)的溫帶和熱帶分布型木本被子植物在葉習(xí)性和生長型組成,以及所處環(huán)境的光、溫、水和土壤條件存在什么差異。(2)亞熱帶闊葉林中的溫帶和熱帶分布型木本被子植物的相對優(yōu)勢(即物種數(shù)的比值),主要受到什么環(huán)境因子的控制。
1 材料與方法
1.1 研究地
本研究涉及木本被子植物788種,包括747個種和41個種下單元, 分屬于91科267屬,均為亞熱帶闊葉林的組成種,來源于8個亞熱帶闊葉林動態(tài)樣地(車儉等, 2020)。樣地面積為5~50 hm2,分布地理范圍為110°25′—121°47′ E、23°10′—29°48′ N。樣地年平均氣溫為11.5~20.9 ℃,年平均降水量為1 375.0~4 067.0 mm (宋永昌等, 2015; Ricklefs & He, 2016)。單個樣地所含木本被子植物種類物種數(shù)為110~238個。本研究的物種名錄、葉習(xí)性(leaf habit)、生長型(growth form)和生態(tài)習(xí)性(ecological behavior)參考宋永昌等(2015)。葉習(xí)性包括常綠闊葉和落葉闊葉;生長型包括喬木、灌木和藤本;生態(tài)習(xí)性包括光照(light, L)、溫度(temperature, T)、水分(water, W)、土壤反應(yīng)(soil reaction, R)和土壤肥力(soil fertility, N)共5項(xiàng)指標(biāo),各項(xiàng)指標(biāo)均劃分為1~9級。從1級的強(qiáng)陰生、冰雪、強(qiáng)旱生、強(qiáng)酸性土和極貧土到9級的強(qiáng)陽生、高溫、強(qiáng)濕生、堿性土和極肥土,植物所處環(huán)境的光照、溫度、水分、土壤pH和土壤肥力逐漸升高。屬的分布區(qū)類型參考吳征鎰等(2010),分為世界分布(cosmopolitan; 分布區(qū)類型1)、熱帶分布(tropical; 分布區(qū)類型2~7)和溫帶分布(temperate; 分布區(qū)類型8~15)。其中,因?yàn)槭澜绶植紖^(qū)類型僅2屬6種,數(shù)量過少,所以本研究分析只針對熱帶分布區(qū)和溫帶分布區(qū)類型植物,共265屬782種。782種木本被子植物的譜系樹通過R語言編寫的 “S.PhyloMaker”函數(shù)中的“Scenario 3”構(gòu)建 (Qian & Jin, 2016)。種以下分類單元被加到其物種枝長的 1/2 處。在該譜系樹的基礎(chǔ)上,每個屬保留一個分類單元,使用R軟件(R Core Team, 2019)ape軟件包的“drop.tip”函數(shù)(Paradis & Schliep, 2018)去掉多余的分類單元,獲得屬級譜系樹。此外,本研究中的環(huán)境因子包括樣地面積、島嶼(是否位于島嶼上)、年平均氣溫(mean annual temperature, MAT)、年降水量(annual precipiation, AP)和旱季持續(xù)時間(dry season length, DS)(Ricklefs & He, 2016)。其中,取樣面積(樣地面積)和空間隔離(島嶼)是影響植物分布的重要空間因素,氣溫和降水是影響植物地理分布的重要?dú)夂蛞蛩兀⊿hen et al., 2009; Qian et al., 2020)。
1.2 統(tǒng)計分析
首先,統(tǒng)計了不同分布區(qū)植物的葉習(xí)性和生長型組成。其次,通過R軟件phytools軟件包的“phylosig”函數(shù)(Revell, 2012),分析了屬分布型的譜系信號。然后,通過R 軟件的“chisq.test”函數(shù)進(jìn)行卡方檢驗(yàn),對比了熱帶和溫帶分布型屬植物在葉習(xí)性和生長型的物種數(shù)量比例在地區(qū)和樣地尺度上是否存在差異。通過R軟件caper軟件包的“pgls”函數(shù)和“anova”函數(shù)(Orme et al., 2018)分別進(jìn)行譜系線性回歸(phylogenetic linear model)和方差分析(analysis of variance),對比了熱帶和溫帶分布型屬植物的生態(tài)習(xí)性差異,以及不同葉習(xí)性和生長型的熱帶和溫帶分布型屬植物的生態(tài)習(xí)性差異。其中,對比不同生長型的熱帶和溫帶分布型屬植物生態(tài)習(xí)性差異時,由于已有數(shù)據(jù)顯示溫帶和熱帶分布型屬木質(zhì)藤本分別僅有3屬6種和12屬14種,數(shù)量過少,且部分樣地未公布藤本數(shù)據(jù),所以僅對比灌木和喬木。通過“pgls”函數(shù)和“anova”函數(shù),控制了葉習(xí)性和生長型的作用,以了解屬分布型對于各項(xiàng)生態(tài)習(xí)性的獨(dú)立影響。最后,通過R軟件“l(fā)m”函數(shù)進(jìn)行線性回歸,以及MuMIn軟件包的“dredge”“model.sel”和“model.avg”函數(shù)(Bartoń, 2019)進(jìn)行模型選擇,分析了環(huán)境因子對于溫帶分布型屬和熱帶分布型屬植物的物種數(shù)量比值的影響。模型選擇過程中,模型間擬合顯著差異的評價標(biāo)準(zhǔn)為兩者的AICc差異≥5。若存在與最優(yōu)模型(AICc值最小模型)無顯著差異的模型,則將其和最優(yōu)模型進(jìn)行模型平均(model averaging),得到條件平均(conditional average)作為最優(yōu)擬合結(jié)果。以上所有分析在R 3.5.3軟件中進(jìn)行(R Core Team, 2019)。
2 結(jié)果與分析
本研究涉及的265屬782種被子植物中,熱帶分布型占167屬483種,溫帶分布型占98屬299種。對265個屬所構(gòu)建的屬級譜系樹(圖1)進(jìn)行分析,結(jié)果顯示屬分布型具有顯著的譜系信號 (λ=0.935,P<0.001)。
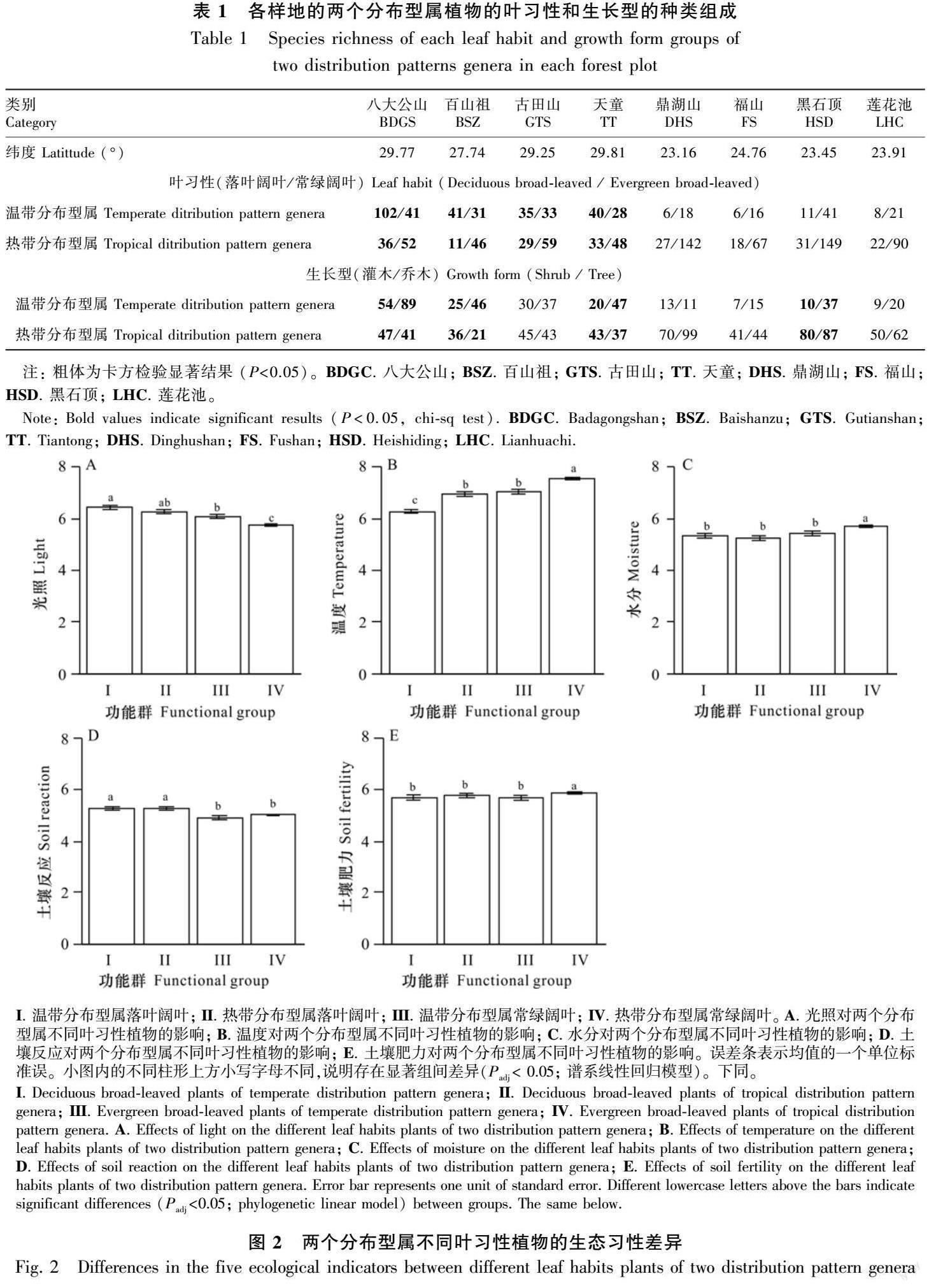
卡方檢驗(yàn)顯示,在地區(qū)尺度上,按葉習(xí)性劃分,溫帶分布型屬植物有落葉闊葉158種,常綠闊葉141種,熱帶分布型屬植物有落葉闊葉123種,常綠闊葉360種,從而得出熱帶分布型屬植物多為常綠闊葉種,溫帶分布型屬則含落葉闊葉種較多(χ2 = 58.94,P<0.001);按生長型劃分,溫帶分布型屬植物有灌木110種,喬木植物183種,而熱帶分布型屬植物灌木231種,喬木植物238種,與熱帶分布型屬相比,溫帶分布型屬所含喬木比例較高(χ2 = 9.53,P= 0.002)。在樣地尺度上,按葉習(xí)性分,4個較高緯度樣地內(nèi)的溫帶分布型屬植物比熱帶分布型屬植物含有更大比例的落葉植物,而在較低緯度這種差異不明顯;按生長型分,也有4個樣地呈現(xiàn)出差異,其中3個樣地所在緯度較高,表現(xiàn)為溫帶分布型屬植物含有的喬木比例高于熱帶分布型屬植物(表1)。
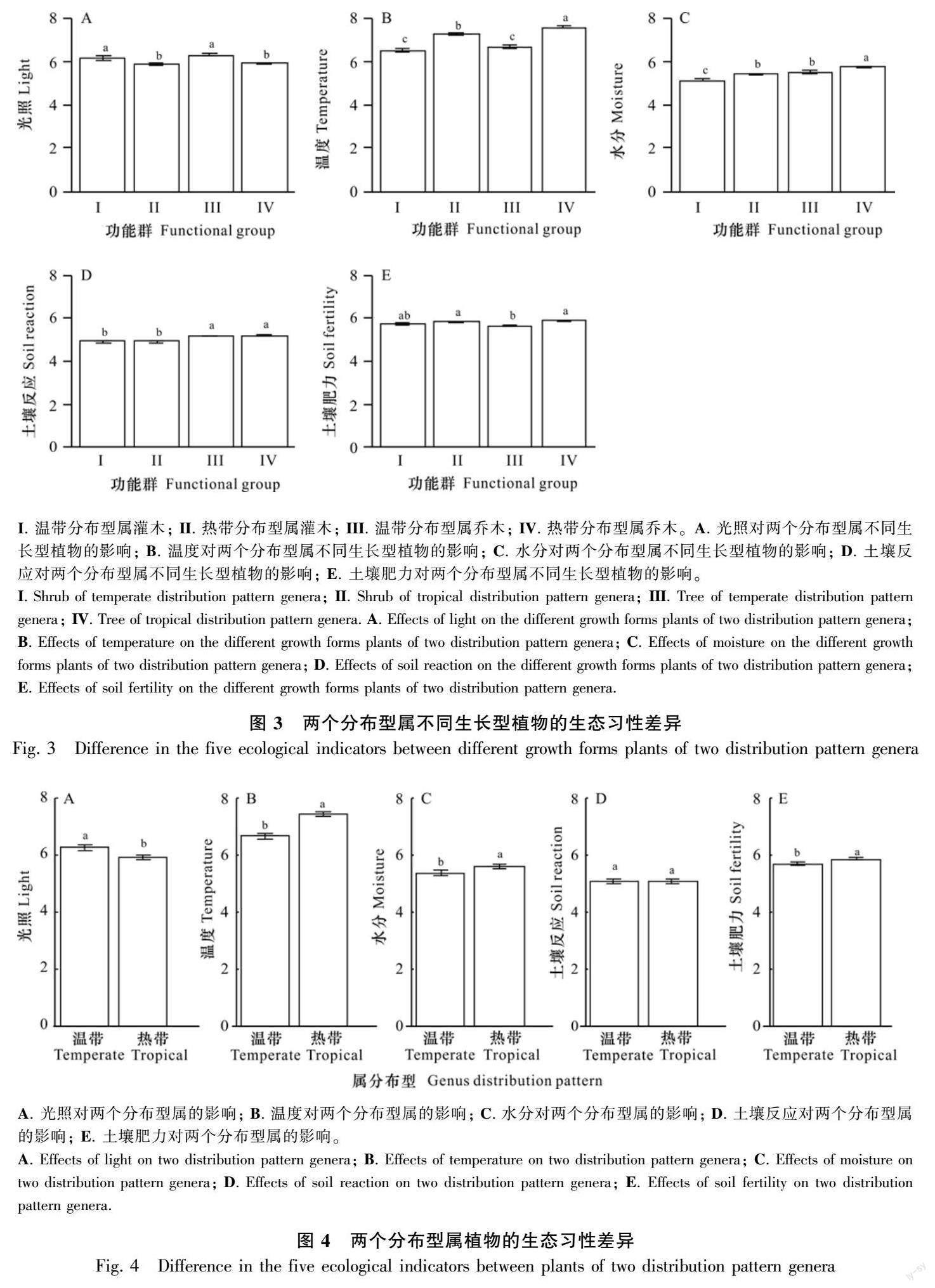
譜系線性回歸分析顯示,葉習(xí)性和生長型對生態(tài)習(xí)性均有顯著影響(圖2和圖3)。具體而言,相同葉習(xí)性的不同分布型屬植物在生態(tài)習(xí)性上存在差異(圖2)。對于落葉闊葉植物,溫帶分布型除了溫度低于熱帶分布型外,在光照、水分、土壤反應(yīng)和土壤肥力上與熱帶分布型均無區(qū)別;對于常綠闊葉植物,溫帶分布型除了土壤反應(yīng)與熱帶分布型無區(qū)別外,在光照、水溫度、水分和土壤肥力上與熱帶分布型均存在顯著區(qū)別。相同生長型的不同分布型屬植物的生態(tài)習(xí)性也存在較多差異(圖3)。對于灌木,溫帶分布型在土壤反應(yīng)和土壤肥力上與熱帶分布型無區(qū)別,而在光照、溫度和水分方面與熱帶分布型存在明顯區(qū)別;對于喬木,溫帶分布型除了土壤反應(yīng)與熱帶分布型無區(qū)別外,在光照、溫度、水分和土壤肥力上與熱帶分布型均存在明顯區(qū)別。
不控制葉習(xí)性和生長型的作用時,溫帶分布型屬植物有更高的光照、更低的溫度和水分需求,而土壤肥力和土壤反應(yīng)則與熱帶分布型屬植物無區(qū)別(圖4)。當(dāng)控制了葉習(xí)性和生長型的作用后,兩個分布型屬植物在光照、氣溫和水分需求方面仍然存在同樣的差異(表2)。
線性回歸模型顯示,無論對于所有植物,還是根據(jù)性狀劃分為不同功能群,溫帶分布型屬和熱帶分布型屬植物的物種數(shù)量比例都受到年平均氣溫的控制。均表現(xiàn)為隨著年平均氣溫上升,溫帶分布型屬植物的比例下降,熱帶分布型屬植物的比例上升。但對于常綠闊葉植物,還受到年降水量的微弱影響,表現(xiàn)為隨年降水量增大,溫帶分布型屬常綠植物的比例下降,熱帶分布型屬常綠植物的比例上升(表3)。
3 討論與結(jié)論
本研究所在的亞熱帶地區(qū)介于熱帶與北溫帶之間,該地區(qū)植被具有復(fù)雜的區(qū)系組成(吳征鎰 1965; 王荷生, 1979),古老的歷史和豐富的植物多樣性(Lu et al., 2018;Qian et al., 2019)。本研究所涉及植物均為亞熱帶地區(qū)闊葉林內(nèi)木本植物,來源于265個屬。這些屬的分布型呈現(xiàn)出一定程度的譜系保守性(phylogenetic conservatism),可能是受到歷史(Ricklefs, 1987; Brown, 2014)和生態(tài)位的譜系保守性(Donoghue, 2008)等過程所驅(qū)動。
不同屬分布型植物的生活史性狀具有明顯的差異。一方面,在地區(qū)尺度上,溫帶分布型屬含有較大比例的落葉種類,可能與氣候等環(huán)境因素有關(guān)(Givnish, 1979; Terborgh, 1985)。溫帶分布型屬所在的高緯度溫帶氣候通常具有明顯的季節(jié)性變化,而落葉被認(rèn)為在季節(jié)性差異明顯的氣候中更具有優(yōu)勢(Givnish, 1979; Edwards et al., 2017; Zanne et al., 2018)。在樣地尺度上,該格局僅出現(xiàn)在較高緯度的4個闊葉林內(nèi),表明溫帶分布型屬的落葉物種在適應(yīng)季節(jié)性氣候方面的優(yōu)勢(Givnish, 2002; Zanne et al., 2014)也顯現(xiàn)在群落層面,在較高緯度地帶更加顯著。另一方面,在地區(qū)尺度上,溫帶分布型屬植物含有較大比例的喬木種類,這在一定程度上可能是由于中高緯度地帶的太陽光入射傾斜角度大,穿透到林下的直射光線少,導(dǎo)致林內(nèi)垂直分層不明顯(Terborgh, 1985),林相結(jié)構(gòu)簡單所致;而越往低緯度地帶移動,太陽光入射角度越垂直,更易于穿透到林下,林內(nèi)獲得的直射光更多,且能量密度更大,垂直分層更明顯,林相結(jié)構(gòu)復(fù)雜,因此林下能夠維持的灌木等植物也就較多(Terborgh, 1985; Spicer et al., 2020)。在樣地尺度上,該格局主要出現(xiàn)在較高緯度的闊葉林樣地內(nèi),傾向于支持溫帶分布型屬的喬木對于適應(yīng)較高緯度地帶季節(jié)性氣候較灌木具有一定的優(yōu)勢,部分原因可能是由于灌木根系通常較淺,而寒冷氣候條件下土壤淺層水分凝凍等風(fēng)險較大,導(dǎo)致灌木的水分利用受到限制(Morales et al., 2012; Yang et al., 2020)以及根系損害等。
不同分布型屬植物的生態(tài)習(xí)性也存在明顯差異。在與本研究相同的森林中,車儉等(2020)發(fā)現(xiàn)葉習(xí)性對木本被子植物的生態(tài)習(xí)性存在廣泛的影響。在此基礎(chǔ)上,本研究進(jìn)一步發(fā)現(xiàn)在控制了葉習(xí)性等生活史性狀對生態(tài)習(xí)性的影響后,溫帶和熱帶分布型屬植物在溫度、水分和光照等生態(tài)習(xí)性上,仍然存在顯著的區(qū)別,這表明不同分布型屬植物之間存在獨(dú)立于葉習(xí)性等生活史性狀的生態(tài)習(xí)性差異。例如,雖然均為落葉闊葉喬木, 但歸類于溫帶分布型屬的闊葉槭(Acer amplum)和水青岡(Fagus longipetiolata)所處環(huán)境的溫度卻要低于熱帶分布型屬的樸樹(Celtis sinensis)和赤楊葉(Alniphyllum fortunei) (宋永昌等, 2015)。由于亞熱帶森林中的熱帶分布型屬被子植物通常較為古老(吳征鎰, 1965),本研究所發(fā)現(xiàn)的熱帶分布型屬植物對于高溫、濕潤和低光照環(huán)境的偏好,在一定程度上可能與其生態(tài)習(xí)性的保守性以及地球氣候的歷史變化有關(guān)(Donoghue, 2008; Qian & Ricklefs, 2016)。被子植物開始快速分化時的地球環(huán)境,被認(rèn)為類似于溫暖、潮濕和季節(jié)性不明顯的熱帶環(huán)境;而新生代以來全球氣溫的下降,低溫、干燥的溫帶環(huán)境逐漸盛行,蘊(yùn)育了溫帶分布型屬植物(Qian & Ricklefs, 2016; Igea & Tanentzap, 2020)。
亞熱帶地區(qū)是溫帶和熱帶分布型屬植物在地理上形成充分交錯分布的緯度帶(Qian et al., 2003)。本研究中,從兩個分布型屬植物在亞熱帶森林中的更替隨年平均氣溫變化的趨勢可以推測氣溫可能是兩個分布型屬植物生態(tài)習(xí)性差異的主要環(huán)境因子,這個結(jié)果也與以往對整個東亞尺度的分析結(jié)果一致(Qian et al., 2003)。溫度對于溫帶分布型屬和熱帶分布型屬植物截然相反的影響,在一定程度上說明兩個分布型屬植物對于低溫冰凍等的適應(yīng)能力存在差異(Wang et al., 2011; Zanne et al., 2014)。此外,本研究發(fā)現(xiàn)兩個分布型屬常綠闊葉植物的地理更替除了受到氣溫影響,還受到降水的微弱影響,反映出熱帶分布型屬常綠植物對于水分的要求比溫帶分布型屬常綠植物更高,這可能與代謝等因素有關(guān)(Brown et al., 2004)。
總之,本研究提示亞熱帶地區(qū)的溫帶和熱帶分布型屬植物在生活史性狀和生態(tài)習(xí)性上存在廣泛的區(qū)別。可能正是由于這些區(qū)別,兩類植物雖然共同存在于亞熱帶地區(qū)的森林中,但對于溫度等氣候因子的響應(yīng)特點(diǎn)迥異。氣候變化場景下,這些差異有可能會驅(qū)動亞熱帶森林的區(qū)系特征發(fā)生改變。值得注意的是,本研究僅針對不同屬分布型進(jìn)行了分析,研究發(fā)現(xiàn)也僅適用于屬這一層級。對于本研究發(fā)現(xiàn)是否也適用于屬以下的分類層級,例如物種層級,需要更為深入的研究。
致謝 感謝中國森林生物多樣性監(jiān)測網(wǎng)絡(luò)的建設(shè)者們。感謝兩位審稿人提出的寶貴建議。
參考文獻(xiàn):
BARTO K, 2019. MuMIn: multi-model inference. R package version 1.43. 15 [J]. Retrieved May, 11: 2020.
BROWN JH, 2014. Why are there so many species in the tropics? [J]. J Biogeogr, 41(1): 8-22.
BROWN JH, GILLOOLY JF, ALLEN AP, et al., 2004. Toward a metabolic theory of ecology [J]. Ecology, 85(7): 1771-1789.
CHE J, ZHENG J, JIANG Y, et al., 2020. Separation of phylogeny and ecological behaviors between evergreen and deciduous woody angiosperms in the subtropical forest dynamics plots of China [J].Chin J Plant Ecol,44(10): 1007-1014. [車儉, 鄭潔, 蔣婭, 等, 2020. 中國亞熱帶森林動態(tài)監(jiān)測樣地常綠和落葉木本被子植物譜系結(jié)構(gòu)及生態(tài)習(xí)性差異 [J]. 植物生態(tài)學(xué)報, 44(10): 1007-1014.]
DONOGHUE MJ, 2008. A phylogenetic perspective on the distribution of plant diversity [J]. Proc Natl Acad Sci USA, 105(Supplement 1): 11549-11555.
EDWARDS EJ, CHATELET DS, CHEN BC, et al., 2017. Convergence, consilience, and the evolution of temperate deciduous forests [J]. Am Nat, 190(S1): S87-S104.
ELLENBERG H, 1979. Zeigerwerte der Gefpflanzen Mitteleuropas, 2. Aufl., Scripta, GeobotanicaIX [M]. Gttingen: Verlag E. Goltze.
GIVNISH TJ, 1979.On the adaptive significance of leaf form [J]. Top Plant Pop Biol: 375-407.
GIVNISH TJ, 2002. Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox [J]. Silv Fenn, 36(3): 703-743.
IGEG J, TANENTZAP AJ, 2020. Angiosperm speciation cools down in the tropics [J]. Ecol Lett, 23(4): 692-700.
LU L, MAO L, YANG T, et al., 2018. Evolutionary history of the angiosperm flora of China [J]. Nature, 554(7691): 234-238.
MORALES AG, MENDOZA JMO, GOZALBO ME, et al., 2012. Arboreal and prostrate conifers coexisting in Mediterranean high mountains differ in their climatic responses [J]. Dendrochronologia, 30(4): 279-286.
ORME D, FRECKLETON R, THOMAS G, et al., 2018. caper: Comparative analyses of phylogenetics and evolution in R [J/OL]. R package version 1.0.1. https://CRAN.R-project.org/package=caper.
PARADIS E, SCHLIEP K, 2018. ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R [J]. Bioinformatics, 35(3): 526-528.
QIAN H, JIN Y, 2016. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure [J]. J Plant Ecol, 9(2): 233-239.
QIAN H, RICKLEFS RE, 2016. Out of the tropical lowlands: latitude versus elevation [J]. Trend Ecol Evol, 31(10): 738-741.
QIAN H, SONG J, KRESTOV P, et al., 2003. Large-scale phytogeographical patterns in East Asia in relation to latitudinal and climatic gradients [J]. J Biogeogr, 30(1): 129-141.
QIAN H, WANG S, HE J, et al., 2006. Phytogeographical analysis of seed plant genera in China [J]. Ann Bot, 98(5): 1073-1084.
QIAN H, DENG T, JIN Y, et al., 2019. Phylogenetic dispersion and diversity in regional assemblages of seed plants in China [J]. Proc Natl Acad Sci USA, 116(46): 23192-23201.
QIAN H, JIN Y, LEPRIEUR F, et al., 2020. Geographic patterns and environmental correlates of taxonomic and phylogenetic beta diversity for large-scale angiosperm assemblages in China [J]. Ecography, 43(11): 1706-1716.
R Core Team, 2019. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria [EB/OL]. [2021-03-22]. https://www.R-project.org/.
REVELL LJ, 2012. phytools: An R package for phylogenetic comparative biology (and other things) [J].Method Ecol Evol, 3(2): 217-223.
RICKLEFS RE, HE F, 2016. Region effects influence local tree species diversity [J]. Proc Natl Acad Sci USA, 113(3): 674-679.
RICKLEFS RE, 1987. Community diversity: relative roles of local and regional processes [J]. Science, 235(4785): 167-171.
SHEN G, YU M, HU XS, et al., 2009. Species-area relationships explained by the joint effects of dispersal limitation and habitat heterogeneity [J]. Ecology, 90(11): 3033-3041.
SONG YC, 2013. Evergreen broad-leaved forests in China [M]. Beijing: Science Press. [宋永昌, 2013. 中國常綠闊葉林 [M]. 北京: 科學(xué)出版社.]
SONG YC, YAN ER, SONG K, 2015. Synthetic comparison of eight dynamics plots in evergreen broadleaf forests, China [J]. Biodivers Sci, 23(2): 139-148. [宋永昌, 閻恩榮, 宋坤, 2015. 中國常綠闊葉林8大動態(tài)監(jiān)測樣地植被的綜合比較 [J]. 生物多樣性, 23(2): 139-148.]
SPICER ME, MELLOR H, CARSON WP, 2020. Seeing beyond the trees: A comparison of tropical and temperate plant growth forms and their vertical distribution [J]. Ecology, 101(4): e02974.
SUN H, DENG T, CHEN YS, et al., 2017. Current research and development trends in floristic geography [J]. Biodivers Sci, 25(2): 111-122. [孫航, 鄧濤, 陳永生, 等, 2017. 植物區(qū)系地理研究現(xiàn)狀及發(fā)展趨勢 [J]. 生物多樣性, 25(2): 111-122.]
SU X, SHRESTHA N, XU X, et al., 2020. Phylogenetic conservatism and biogeographic affinity influence woody plant species richness-climate relationships in eastern Eurasia [J]. Ecography, 43(7): 1027-1040.
TERBORGH J, 1985. The vertical component of plant species diversity in temperate and tropical forests [J]. Am Nat, 126(6): 760-776.
WANG HS, 1979. The basic features of Chinese flora [J]. Acta Geogr Sin, 34: 224-237. [王荷生, 1979. 中國植物區(qū)系的基本特征 [J]. 地理學(xué)報, 34: 224-237.]
WANG HS, 1992. Floristic Geography [M]. Beijing: Science Press. [王荷生, 1992. 植物區(qū)系地理 [M]. 北京: 科學(xué)出版社.]
WANG Z, FANG J, TANG Z, et al., 2011. Patterns, determinants and models of woody plant diversity in China [J]. Proc Roy Soc Biol Sci, 278(1715): 2122-2132.
WU ZY, 1965. Tropical relatives of Chinese flora [J]. Sci Bull, 16: 25-33. [吳征鎰, 1965. 中國植物區(qū)系的熱帶親緣 [J]. 科學(xué)通報, 16: 25-33.]
WU ZY, 1980.Vegetation of China [M]. Beijing: Science Press. [吳征鎰, 1980. 中國植被 [M]. 北京: 科學(xué)出版社.]
WU ZY, SUN H, ZHOU ZK, et al., 2010. Floristics of seed plants form China [M]. Beijing: Science Press. [吳征鎰, 孫航, 周浙昆, 等, 2010. 中國種子植物區(qū)系地理 [M]. 北京: 科學(xué)出版社.]
XU W, SVENNING JC, CHEN G, et al., 2018. Plant geographical range size and climate stability in China: Growth form matters [J]. Glob Ecol Biogeogr, 27(5): 506-517.
XU X, WANG Z, RAHBEK C, et al., 2013. Evolutionary history influences the effects of water-energy dynamics on oak diversity in Asia [J]. J Biogeogr, 40(11): 2146-2155.
XU Y, SHEN Z, ZHANG J, et al., 2021. The effects of multi-scale climate variability on biodiversity patterns of Chinese evergreen broad-leaved woody plants: growth form matters [J]. Front Ecol Evol, 8: 540948.
YANG J, COOPER DJ, LI Z, et al., 2020. Differences in tree and shrub growth responses to climate change in a boreal forest in China [J]. Dendrochronologia, 63: 125744.
ZANNE AE, TANK DC, CORNWELL WK, et al., 2014. Three keys to the radiation of angiosperms into freezing environments [J]. Nature, 506(7486): 89-92.
ZANNE AE, PEARSE WD, CORNWELL WK, et al., 2018. Functional biogeography of angiosperms: life at the extremes [J]. New Phytol, 218(4): 1697-709.
ZHAO Y, CAO H, XU W, et al., 2018. Contributions of precipitation and temperature to the large scale geographic distribution of fleshy-fruited plant species: Growth form matters [J]. Sci Rep, 8(1): 17017.
ZHU H, MA Y, YAN L, et al., 2007. The relationship between geography and climate in the generic-level patterns of Chinese seed plants [J]. Acta Phytotax Sin, 45(2): 134-166.
(責(zé)任編輯 周翠鳴)
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
保健醫(yī)苑(2021年7期)2021-08-13 08:48:02
學(xué)生天地(2020年36期)2020-06-09 03:12:30
小學(xué)科學(xué)(學(xué)生版)(2020年5期)2020-05-25 07:11:32
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
