圓唇苣苔屬(Gyrocheilos)花柱側偏彎折現象及其傳粉適應機制
2023-06-18 10:56:19孫浩然凌少軍任明迅
廣西植物 2023年5期
孫浩然 凌少軍 任明迅

摘 要: 圓唇苣苔屬(Gyrocheilos)是苦苣苔科的中國特有屬,有5種,全部狹域分布在我國西南及廣東的高海拔山區。圓唇苣苔屬所有物種的花柱側偏且花柱頂端呈90°彎折,使得柱頭位于花開口的中央位置。這種獨特的側偏彎折花柱結構,說明圓唇苣苔屬可能有著特殊的演化歷史和適應機制。為揭示這種特殊的花柱側偏彎折現象的發生范圍、發育過程及其傳粉適應機制,該研究在圓唇苣苔(Gyrocheilos chorisepalus)、折毛圓唇苣苔(G. retrotrichus)和微毛圓唇苣苔(G. microtrichus)3個物種中開展了花部綜合征觀察,并研究了廣東大霧嶺保護區內的折毛圓唇苣苔花發育過程、花部特征和繁育系統以及傳粉過程。結果表明:(1)微毛圓唇苣苔只有花柱左偏彎折現象,而圓唇苣苔和折毛圓唇苣苔雖然大部分花是花柱左偏彎折,但在部分個體中出現了少量的花柱右偏彎折現象(占種群總花數的2%~3%)。(2)傳粉觀察發現,折毛圓唇苣苔在花蕾期即出現了花柱彎折現象,2個可育雄蕊的花藥合生、位于花冠筒喉部中央位置,與側偏花柱不存在左右鏡像對稱關系。(3)折毛圓唇苣苔的花粉胚珠比(P/O)為456.98±15.55,屬于兼性異交繁育系統。折毛圓唇苣苔存在一定的傳粉限制,自交授粉可以結實,但異交種子萌發率更高,可能存在近交衰退。(4)折毛圓唇苣苔的訪花昆蟲較少,訪花頻率較低,主要訪花昆蟲有隧蜂、熊蜂、食蚜蠅等;熊蜂體型較大,訪花時降落在彎折花柱和花瓣下唇,胸部側面及下部能有效接觸到柱頭。(5)反射率結果顯示,折毛圓唇苣苔花瓣反射波長范圍集中在紫光和藍紫光區域,花冠的反射波長范圍與蜂類視覺范圍一致且花冠筒外側和花瓣下唇的反射強度最大,更容易吸引蜂類落置在花冠寬大的下唇;圓唇苣苔屬的花柱側偏彎折現象可能來自近緣的長蒴苣苔屬(Didymocarpus)的花柱下彎現象或鏡像花(mirror-image flowers)。綜上認為,這種側偏彎折的花柱,可能通過提供昆蟲降落平臺,使得柱頭位于花開口中央和花瓣下唇的上方位置,提高了柱頭接觸訪花昆蟲的概率,是適應高海拔地區低頻率訪花者的一種機制。
關鍵詞: 花部綜合征, 鏡像花, 傳粉機制, 繁育系統, 苦苣苔科
中圖分類號: Q944? 文獻標識碼: A? 文章編號: 1000-3142(2023)05-0817-10
Abstract: Gyrocheilos is a small genus of Gesneriaceae endemic to hight altitude mountains in Southwest China and Guangdong Province, with only five species. This genus is characterized by laterally bending style, which curves 90° at the top of the style and the stigma right at the mouth of floral tube. This unusual floral trait may have special evolutionary and adaptive mechanisms. In this paper, three Gyrocheilos species, i.e. G. chorisepalus, G. retrotrichus, G. microtrichus were examined to figure out the developmental patterns of floral lateral bending. Breeding systems, floral syndrome and pollination processes were also studied in G. retrotrichus at Dawuling Mountain, Guangdong Province, to explore its pollination adaptations. The results were as follows: (1) G. microtrichus had only left-bending style, while right-bending style were observed in several flowers (2%-3% of the total flowers) of G. chorisepalus and G. retrotrichus, although the individuals and populations were dominated by left-bending style. (2) Pollination observation found that style lateral bending occurred at the early stage of floral bud in G. retrotrichus, with two fertile stamens were anther-united and hidden at the middle of the floral throat, and there was no left and right mirror symmetry relationship with the lateral style. (3) The pollen-ovule ratio (P/O) was ( 456.98±15.55), belonging to facultative outcross breeding system. G. retrotrichus had a certain pollen limitation and self-pollination, but the germination rate of outcross seeds was higher, and there might be inbreeding decline. (4) There were few floral visitors and the frequency was low of G. retrotrichus. The main floral visitors were Halictidae, Bombus and Syrphidae. Bombus were large, and they landed on the bent style and the lower lip of the petals when visiting flowers. The side and lower part of the chest could effectively contact the stigma. (5) The detected reflected lights of flowers were composed of purple and blue-purple lights and the lower lips and outer floral tube with the highest reflection intensity, more likely to attract bees to land on the corollas lower lip. Molecular phylogeny indicated that the closest genus of Gyrocheilos was Didymorcarpus, which was characterized by style downward bending and mirror-image flowers, suggesting the style lateral bending in Gyrocheilos probably evolved from either of these conditions. In conclusion, the lateral bending style might provide landing platform for pollinators and the stigma above the lower lips increase contact probability the floral visitors, which is an adaptation to the very low insect visitation due to fragmented and foggy habitats in high altitude areas.
Key words: floral syndrome, mirror-image flowers, pollination mechanism, breeding systems, Gesneraiceae
被子植物花柱的形態與空間位置是決定柱頭落置花粉、胚珠授粉成功的一個關鍵因素,往往是由訪問最頻繁、最有效的傳粉者塑造的(Mayfield et al., 2001; Fenster et al., 2004)。在適應不同傳粉者類型與訪花行為以最大化柱頭接受同種花粉概率的選擇壓力下,花柱出現了極大的多態性(Barrett et al., 2000),如花柱運動(Li et al., 2001; Yu & Huang, 2004; 段友愛和李慶軍, 2008)、柱頭裂片開合(Fetscher, 2001; Yang et al., 2004)、異長花柱(Barrett et al., 2000)、鏡像花的側偏花柱(Jesson & Barrett, 2002a, b; Gao et al., 2006; Ren et al., 2013)等,成為研究花部綜合征(floral syndrome)、花-傳粉者協同進化的關鍵性狀之一(黃雙全,2007)。
苦苣苔科(Gesneriaceae)圓唇苣苔屬(Gyrocheilos)的花柱形態非常特別:花柱不僅偏轉在花冠筒的一側,而且花柱頂端呈90°彎折,使得花柱頂端2~3 mm的部分橫在花冠筒開口、柱頭處于花開口處的中央位置(韋毅剛,2010)。這種花柱側偏彎折現象不同于目前已知的花柱類型,可能存在不一樣的發育過程和傳粉適應機制,對認識苦苣苔科傳粉系統演化和物種形成機制具有積極意義。
圓唇苣苔屬是中國特有屬,僅5種2變種(王文采等,1990),狹域分布于廣西與廣東等地的高海拔山區的郁閉林下或溪谷山澗(韋毅剛,2010),生境呈典型的斑塊狀分布。本研究廣泛調查了圓唇苣苔屬3個物種的花柱側偏彎折現象及其主要花部特征,并以折毛圓唇苣苔(Gyrocheilos retrotrichus)為例,研究了花部綜合征及其發育過程、傳粉機制和繁育系統。本文主要回答2個科學問題:(1)圓唇苣苔屬花柱側偏彎折現象的發育過程和主要特征;(2)圓唇苣苔屬花柱側偏彎折現象的傳粉適應機制。
1 材料與方法
1.1 圓唇苣苔屬花柱側偏彎折現象的系統分布
2021年4—5月調查了正處于盛花期的廣西南寧大明山保護區天坪站附近石壁的圓唇苣苔(Gyrocheilos chorisepalus)、廣東大霧嶺大田頂林下山坡的折毛圓唇苣苔、廣東大霧嶺長坑林區山澗邊的微毛圓唇苣苔(G. microtrichus)3個物種的花柱彎折現象。選擇這3個種分布較集中的大種群,記錄開花個體數。隨機挑選不少于種群規模一半的個體,不少于200朵花,統計花柱側偏彎折的現象。
1.2 折毛圓唇苣苔花柱側偏彎折的發生過程
在折毛圓唇苣苔居群內,隨機選取不同植株的25朵花蕾進行標記,記錄雌雄蕊的變化。折毛圓唇苣苔的花蕾發育階段分為5個時期: 花蕾期I、花蕾期Ⅱ、花蕾期Ⅲ、花蕾期Ⅳ、花蕾期Ⅴ(花瓣裂片頂端部分松散,即將打開)、花開放期(花瓣上下唇裂片打開)。每個時期取5朵花進行解剖, 拍照并記錄花內各部位尤其是雌蕊與雄蕊結構的變化。
1.3 訪花昆蟲及訪花行為
參照龔燕兵和黃雙全(2007)的方法,在折毛圓唇苣苔種群內開花個體集中的斑塊中,以50株個體為觀察對象,使用數碼相機對訪花昆蟲進行拍攝觀察。記錄訪花者形態種類、訪花行為和總訪問次數。通過訪花昆蟲是否接觸到柱頭、是否從花開口進入花冠筒等,判斷訪花昆蟲是不是可能的有效傳粉者。
1.4 繁育系統
1.4.1 異交指數與花粉胚珠比(P/O) 依據Dafni(1992)的標準,測量折毛圓唇苣苔30朵花的單花直徑、雌雄蕊成熟順序和柱頭與花藥的相對位置并計算異交指數(OCI)。異交指數越大,植物異交水平越大,越需要傳粉者。
隨機選取10朵花藥尚未開裂的花,放入FAA固定液中,帶回計算花粉和胚珠數量。取下花藥,制成1 000 μL的花粉懸浮液,用移液槍吸取少量花粉液在光學顯微鏡(上海普丹XTL-10)下統計花粉數量,每朵花重復計數3次。在體式顯微鏡(尼康SMZ745T)下剝離出胚珠,統計胚珠數量。
1.4.2 花粉與柱頭活性 參照Dafni(1992)的方法,隨機選取30個處于花蕾期V的花蕾,標記為0狀態。隨后每隔1 d取3朵花,檢測花粉活性和柱頭可授性。方法如下:取部分新鮮花粉撒在滴有15%蔗糖溶液(含硼酸)的凹面載玻片上,置于培養皿中恒溫培養10 h,然后在光學顯微鏡下統計花粉萌發率(花粉數>40為有效樣本)來判斷花粉活性的水平。柱頭浸泡在3%的雙氧水中,根據氣泡產生的速率與數量判斷柱頭活性。
1.4.3 花瓣反射率實驗 取剛開放的花,用便攜式光譜儀(賽曼S3000-UV-NIR)和光纖反射探針(Fiber Y 400 μm)測量花冠筒外側、花冠筒內側、花瓣下唇、花瓣上唇4個部位的花瓣反射率。光源采用脈沖氙燈(Xe-02),參考光使用標準反射白板(WB-02)。使用花下第一片葉作為背景光參照物。測量時將探針傾斜45°并置于距離花瓣表面5 mm處。用SpectraPro V3.3(杭州塞曼科技有限公司)整理反射率數據。
1.4.4 人工授粉套袋實驗 在花冠裂片未完全展開前,隨機選取10個植株上的花做人工處理(Dafni, 1992)。設置的5個處理如下。(1)自然對照:不做任何處理,作自然情況下的對照。(2)無處理套袋:不進行任何處理直接套袋。(3)去雄套袋:在即將開花之前,去掉花藥后套袋。(4)自交授粉:在開花前去雄套袋,花開后用同株的花粉涂在柱頭上,套袋。(5)異交授粉:花開前去雄套袋,花開后用其他植株的花粉涂在柱頭上,套袋。每組10朵,1個月后統計各處理的坐果率和結實率。
1.4.5 種子萌發實驗 取上述5組不同處理得到的膨大果實,每個果實各選取100粒種子,使用蒸餾水清洗3次,將種子置于鋪有滅菌濾紙且用蒸餾水浸潤的培養皿中,恒溫培養。以胚根長度超過種子長度作為種子萌發成功的標志,30 d后統計不同處理下的種子萌發率。通過自交和異交種子的萌發率差異,判斷是否存在近交衰退。
1.5 數據分析
采用SPSS 13.0 for Windows 統計軟件的one-way ANOVA分析套袋實驗中不同處理間的結實率、每果平均種子數的差異。數據間的兩兩比較采用Student-Newman-Keuls檢驗法(S-N-K法)(任明迅, 2009)。
2 結果與分析
2.1 圓唇苣苔屬花柱彎折現象
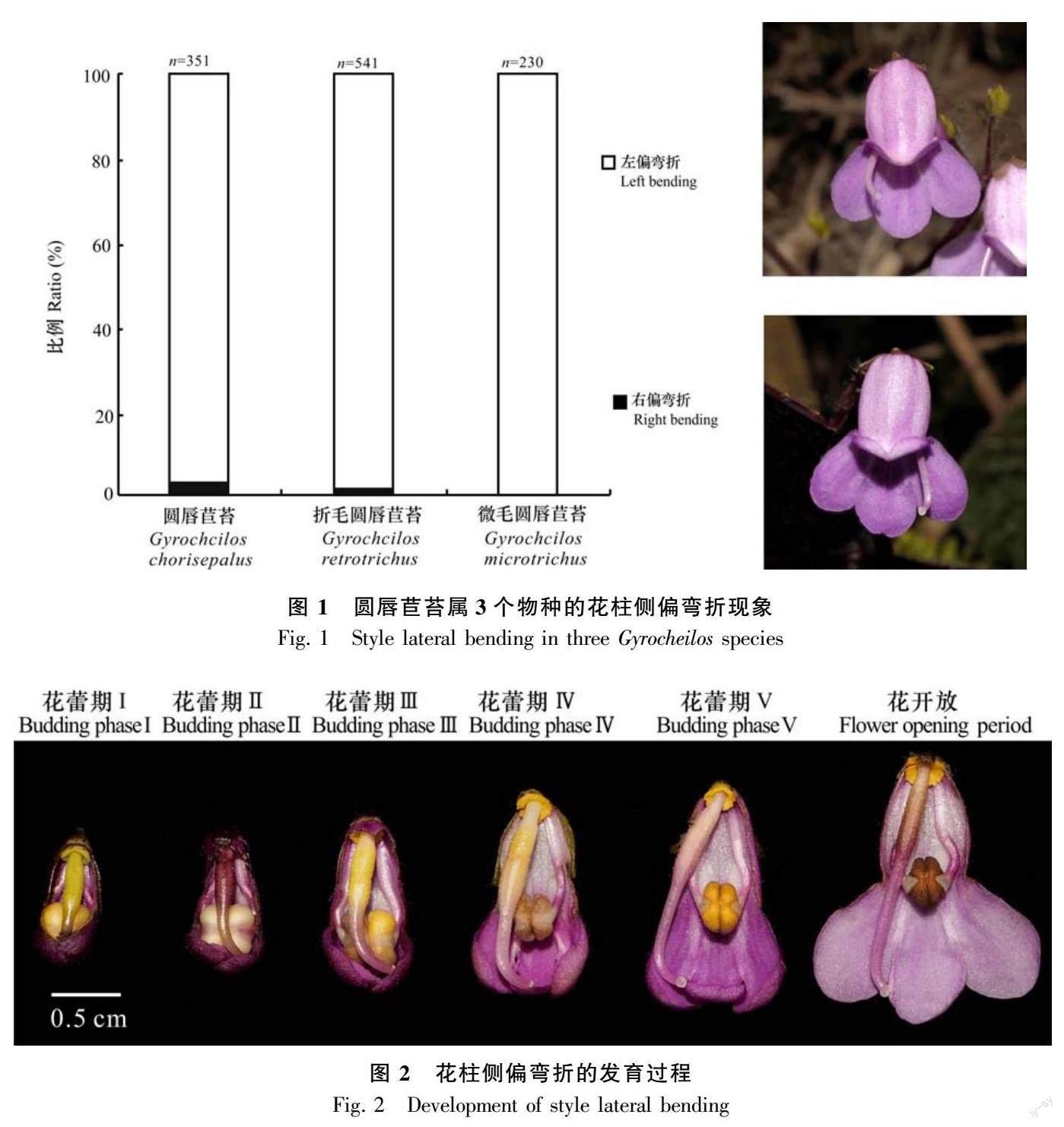
廣西南寧大明山的圓唇苣苔種群個體數超過200株。對種群中351朵花的花柱彎折情況進行了調查,花柱左偏花與右偏花可以出現在同一個體上,其中花柱左偏花341朵、花柱右偏花10朵,花柱右偏花占2.85%(圖1)。
廣東大霧嶺大田頂的折毛圓唇苣苔種群約有2 000個開花個體。統計了種群中541朵花的花柱彎折情況,花柱左偏花與右偏花可以出現在同一個體上,其中花柱左偏花、右偏花分別有532、9朵,花柱右偏花占1.66%(圖1)。
廣東大霧嶺長坑林區的微毛圓唇苣苔有花個體數量約200株。一共調查了230朵花,全部都是左偏彎折花,沒有發現花柱右偏彎折現象(圖1)。
2.2 花部綜合征與花柱側偏彎折過程
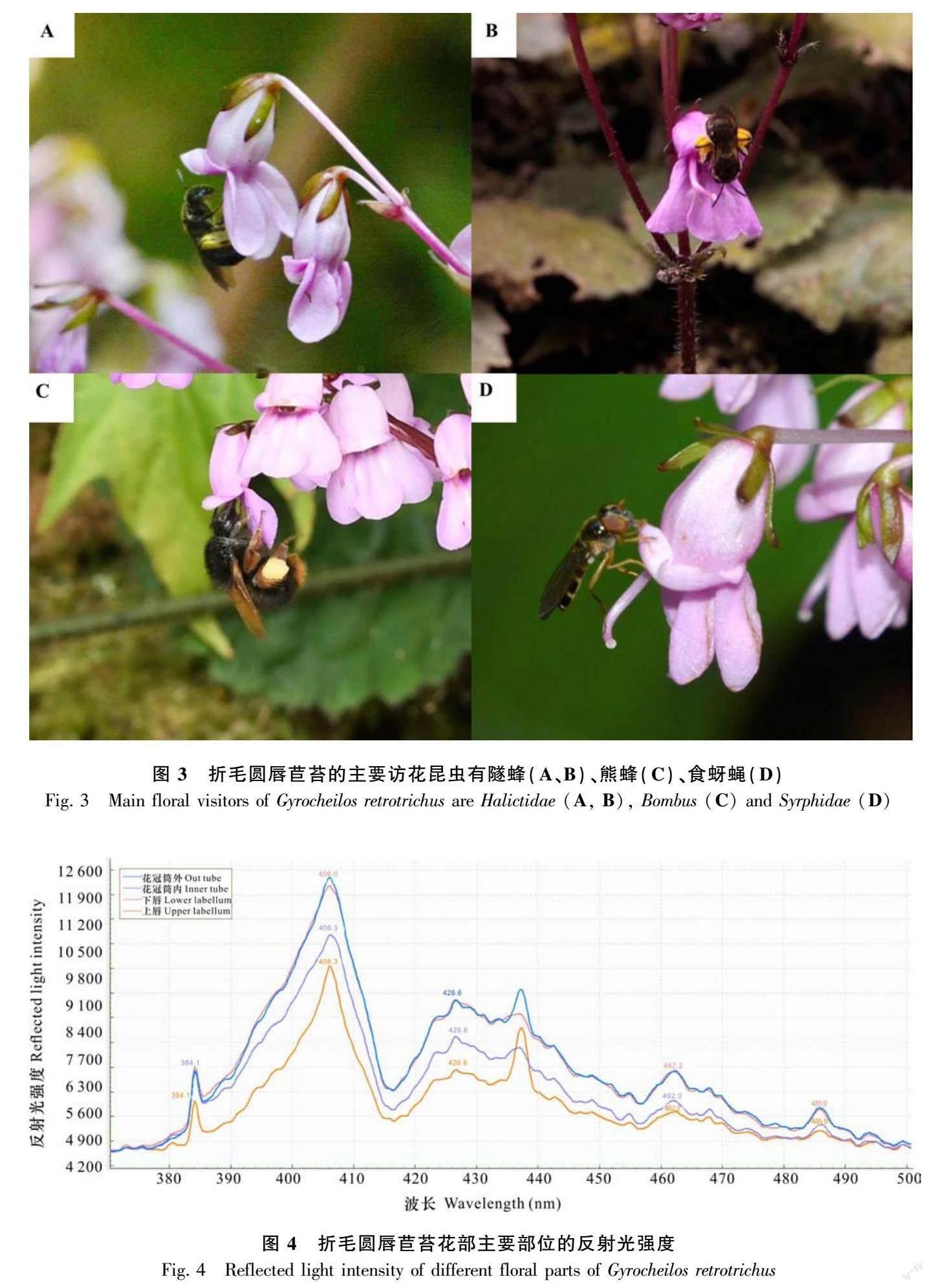
折毛圓唇苣苔花紫紅色,花序大而繁茂,花冠平均長約12 mm。每個植株有1~3條花序,每個花序有5~20朵花,為雙花聚傘花序。折毛圓唇苣苔花大多在凌晨開放,開花時,首先是下唇左右裂片先展開,2~3 h后下唇中裂片展開,上下唇瓣分離,即花開放。花開放后上唇邊緣逐漸向后翻折、下唇繼續向下展開,花柱向花筒外伸長。花蕾期花冠呈深紫紅色,在花開放時逐漸變為淡紫紅色(圖2)。單花壽命通常為6~7 d。
折毛圓唇苣苔花柱基部的花盤明顯,可能分泌有花蜜(圖2)。折毛圓唇苣苔的花柱在距基部約1 mm位置側偏,使整個花柱水平偏向花左側;除了花柱基部的側偏,在花柱頂端靠近柱頭3~5 mm處柱頭還向花中央彎折。折毛圓唇苣苔的雌雄蕊長度近似,花柱位于中央花藥的正上方(圖2:花蕾期I)。隨后,花柱逐漸伸長,柱頭開始向下鉤彎(圖2:花蕾期Ⅱ),并隨著花柱的伸長逐漸離開花藥位置向側面偏移(圖2:花蕾期Ⅲ)。隨著花柱的伸長,雌雄異位程度和花柱側偏更加明顯、花柱上端彎折程度逐漸增大至90°(圖2:花蕾期IV)。在花即將開放時(圖2:花蕾期V),花柱完全側偏在花冠筒側壁,彎折的柱頭位于雄蕊花藥正前方。花完全打開時,花柱遠遠伸出花冠口,彎折的花柱上端呈鐮刀狀橫在花開口處(圖2)。
2.3 傳粉過程
折毛圓唇苣苔主要訪花昆蟲為熊蜂,偶見有隧蜂或食蚜蠅訪問(圖3)。其中,熊蜂的訪問頻率最高,常見于11:00—13:00訪花。熊蜂訪花時常降落在花的下唇(圖3),用腳鉤住下唇或彎折花柱以固定身體。訪花時熊蜂胸部側面和下部能明顯接觸到位于花開口中央位置的柱頭,有時在收集完花粉后也會在花筒口將花粉梳理進花粉籃。熊蜂停留時間較短,平均訪花時間4 s左右,在飛離時,能明顯觀察到由熊蜂振翅引起的花粉散出。
2.4 花瓣反射率
折毛圓唇苣苔花瓣各部位的反射波長集中在384 nm(紫光)、406 nm(紫光)、426~437 nm(紫光)處,在462 nm(藍光)處也有較高的反射強度。花冠的反射波長在蜂類視覺范圍內,說明折毛圓唇苣苔主要靠吸引蜂類傳粉,與傳粉觀察結果一致。折毛圓唇苣苔的花冠筒外側和花瓣下唇的反射強度最大,高于花冠筒內和上唇的反射率(圖4),花冠下唇更高的反射率使得蜂類傳粉時更容易落置在下唇,從而控制其訪花行為。
2.5 繁育系統
2.5.1 異交指數與花粉胚珠比(P/O) 折毛圓唇苣苔的花較大,柱頭與花藥間存在明顯的雌雄異位,雄蕊先熟。根據Dafin(1992)對異交指數的計算標準,折毛圓唇苣苔的異交指數為3,繁育系統為兼性異交,自交親和,可能需要傳粉者。
折毛圓唇苣苔單花的花粉數為(7.33±0.34)×105粒,胚珠數為(1 604±93)個;花粉胚珠比(P/O)為456.98±15.55。依據Cruden(1977)的標準, 折毛圓唇苣苔的繁育系統為兼性異交。
2.5.2 花粉與柱頭活性 雖然折毛圓唇苣苔的花粉在花瓣裂片打開時就有一定活性,但其活性較低 (25%)(圖5), 花粉活性在花開后第3天達到最大值(80%),之后逐漸降低,第8天完全為0。柱頭在開花前沒有活性,在花開放后第3天活性達到最高值,并持續3~4 d(圖5)。
2.5.3 人工套袋 自然條件下,折毛圓唇苣苔的坐果率為36%、種子數為(406±126.03)粒(表1)。去雄套袋結實率為0,不存在無融合生殖。無處理套袋有1朵花成功坐果、種子數為240粒,可能存在一定程度自交。人工授粉的折毛圓唇苣苔顯著高于自然對照的坐果率,顯示折毛圓唇苣苔自然繁殖存在一定的傳粉限制。異交授粉的坐果率略高于自交授粉坐果率,但不存在顯著差異(表1)。
2.5.4 種子萌發率 自然結實的種子萌發率可達(52 ± 11.01)%,無處理套袋的種子萌發率為(24±1.21)%,表明自交的種子也具有一定的活性,人工自交授粉種子的萌發率略高于無處理套袋的萌發率,可能是由無處理套袋果實樣本量極少導致的數據偏差。人工異交授粉的種子萌發率(61±9.86)%顯著高于自交種子的萌發率和自花授粉種子的萌發率(36±9.36)%,說明折毛圓唇苣苔可能存在近交衰退(圖6)。
3 討論
3.1 圓唇苣苔屬花柱側偏彎折現象的特征與演化
圓唇苣苔屬的花柱側偏彎折現象,在形態上類似于鏡像花(mirror-image flowers)。鏡像花是一種特化的昆蟲傳粉機制,通過花柱偏左或偏右,形成鏡像對稱的2種花型促進異交傳粉(Barrett et al., 2000; Jesson & Barrett, 2002c; Ren et al., 2013)。但是,在本研究中,圓唇苣苔屬的花柱側偏彎折現象在2個方面不同于鏡像花:(1)鏡像花通常都有1個對應側偏在另一側的可育雄蕊(Barrett et al., 2000; 張大勇, 2004; Ren et al., 2013),而圓唇苣苔屬的可育雄蕊位于花冠筒內上方的中央位置,不側偏;(2)鏡像花的花柱通常僅側偏、不彎折,柱頭位于花的側面,而圓唇苣苔屬的花柱上端顯著呈90°彎折,使得柱頭位于花開口的中央位置。因此,圓唇苣苔屬花柱側偏彎折現象可能有著特殊的演化歷史和傳粉適應機制。
與圓唇苣苔屬親緣關系最近的長蒴苣苔屬(Didymocarpus)的溫州長蒴苣苔(D. cortusifolius)、沅陵長蒴苣苔(D. yuenlingensis)等植物的花柱也存在彎折現象(韋毅剛, 2010;Roalson & Roberts, 2016),但本研究花柱位于花冠筒正上方,花柱頂端朝下彎折。這種下彎花柱有利于接觸到訪花者背部攜帶的花粉(Armbruster et al., 2003; Huang & Shi, 2013)。我們根據Roalson和Roberts(2016)的分子系統關系,推測圓唇苣苔屬的祖先狀態可能是這種處于花冠筒中央位置的下彎花柱。但是,系統樹改自Roalson和Roberts (2016)。
在后期的演化中,控制花柱細胞分裂的基因發生突變,花絲基部內外側細胞分裂速率與方向存在差異導致花柱側偏, 逐漸形成了側偏的彎折花柱(Du et al., 2012)。這種側偏彎折花柱可以利用訪花者胸部或腹部側面進行傳粉,降低了其與其他植物競爭傳粉者背部傳粉導致的傳粉干擾(Huang & Shi, 2013);同時,側偏彎折的花柱避免了大型訪花昆蟲快速粗暴訪花行為對雌蕊造成的物理傷害,具有保護雌蕊的作用(Barrett et al., 2000)。隨后,在提高柱頭花粉落置成功率的選擇壓力下,花柱頂端彎折程度逐漸增大到90°,形成了圓唇苣苔屬特有的花柱側偏彎折現象。
在本研究中,圓唇苣苔屬的近緣屬長蒴苣苔屬出現了鏡像花,而苦苣苔科蛛毛苣苔屬(Paraboea)鏡像花中也存在著花柱頂端的彎折現象(Gao et al., 2006; 韋毅剛,2010);此外,本研究的圓唇苣苔屬也出現了少量的花柱右偏彎折的花型,與占據絕對優勢的花柱左偏彎折花型呈現出鏡像對稱。這暗示著圓唇苣苔的花柱側偏彎折現象可能與鏡像花有著密切的聯系。但是,鏡像花的異交傳粉機制在傳粉者訪花頻率極低的生境中難以實現與維持(Jesson & Barrett, 2002c)。鏡像花由1個位點的2個等位基因控制,其中右偏花柱是顯性,左偏花柱是隱性(Jesson & Barrett, 2002a,b)。圓唇苣苔屬的生境郁閉度高、濃霧較大,可能導致祖先類群的鏡像花異交機制受限,在經歷了遺傳瓶頸和較高頻率的自交繁殖保障之后,導致只有隱性純合的花柱左偏這一種花型占據絕對優勢(Jesson & Barrett, 2002b),并在后期逐漸增大花柱頂端的彎折程度。鏡像花崩潰成單一或單優花型的現象,在百合科(Liliaceae)和血草科(Haemodoraceae)折扇草屬折扇草(Wachendorfia thyrsiflora)等植物中也有報道(Jesson & Barrett, 2002b,2003)。今后,需要開展深入的分子系統學和遺傳發育機制的研究,確定圓唇苣苔屬花柱彎折和花柱側偏2個性狀的演化順序,才能更好地確定圓唇苣苔屬花柱側偏彎折現象的起源與演化歷史。
3.2 花柱側偏彎折現象的傳粉適應機制
花部綜合征和人工授粉實驗都表明:折毛圓唇苣苔屬于以異交為主的繁育系統,但傳粉限制明顯;自交親和,但存在一定的自交衰退,可能是由脅迫生境造成的傳粉條件與傳粉質量的下降導致的高自交親和度(Schoen & Brown, 1991)。圓唇苣苔屬主要的有效傳粉者熊蜂體型較大,在訪花時只能降落在寬大的花瓣下唇,不能進入花冠筒內部。熊蜂訪花時,經常用前足抓住花瓣上唇和彎折花柱維持身體平衡。在這個過程中,柱頭得以接觸到熊蜂胸部的側面和下部,實現了傳粉。同時,花柱彎折使得柱頭位于花中央、反射率最高的下唇瓣的上方位置,這里是熊蜂等訪花者降落頻率最高的部位,可以最大化提高柱頭接觸傳粉者的概率。因此,彎折的花柱既給熊蜂增加了降落平臺,也提高了柱頭接觸傳粉者的概率和花粉落置成功率,極大促進了傳粉成功率。這可能是圓唇苣苔屬植物對高海拔、濃霧山地傳粉者訪花頻率極低環境的一種適應。
在本研究中,與苦苣苔科其他植物相比,圓唇苣苔屬還有著明顯扁平的花冠筒開口。扁平的花冠筒限制了訪花昆蟲的訪花位置,提高了位于花冠筒開口處的彎折花柱接觸訪花者身體部位的精確性。此外,我們在野外觀察到,折毛圓唇苣苔花在第6天左右開始凋謝,脫落的花冠筒會在風的作用下順著花柱滑落,彎折花柱的柱頭能夠接觸到自花的花藥,此時柱頭有較高的活性,花粉活性較低但仍然具有一定活性,可能在一定程度上促進了自交繁殖保障。這種現象在唇形目紫葳科(Bignoniaceae)等植物也有發現(Lloyd & Schoen, 1992; Qu et al., 2007),被證實是在異交失敗或受限情況下的一種延遲自交機制(Qu et al., 2007)。
因此,圓唇苣苔屬花柱側偏彎折現象是特化適應熊蜂等大型昆蟲胸部側面及下部傳粉的一種機制,具有延遲自交的作用,提高了圓唇苣苔屬在郁閉度高、濃霧、傳粉服務較低等生境的適應能力,對于認識苦苣苔科物種適應分化具有積極意義。這種特殊的花部特征,既有可能是從復雜的特化傳粉系統鏡像花崩潰而來,又有可能是從位于花中央的下彎花柱逐漸偏轉演化而來。在今后的研究中,可以結合圓唇苣苔屬及其近緣屬開展分子系統發育的研究,明確花柱側偏和花柱彎折2個性狀的演化順序,才能更好地確定圓唇苣苔屬花柱側偏彎折現象的起源與適應歷史。
參考文獻:
ARMBRUSTER WS, SHI XQ, HUANG SQ, 2014. Do specialized flowers promote reproductive isolation?realized pollination accuracy of three sympatric Pedicularis species [J]. Ann Bot, 113(2): 331-340.
BARRETT SCH, COLE WW, ARROYO J, et al., 1997. Sexual polymorphisms in Narcissus triandrus (Amaryllidaceae): is this species tristylous? [J]. Heredity, 78: 135-145.
BARRETT SCH, JSEEON LK, BAKER AM, 2000. The evolution and function of stylar polymorphisms in flowering plants [J]. Ann Bot, 85(Suppl. A): 253-265.
CRUDEN RW, 1977. Pollen-Ovule ratios: A conservative indicator of breeding systems in flowering plants [J]. Evolution, 35: 964-974.
DAFNI A, 1992. Pollination ecology: a practical approach [M]. Oxford: Oxford University Press.
DU W, QIN KZ, WANG XF, 2012. The mechanism of stamen movement in Chimonanthus praecox (Calycanthaceae): differential cell growth rates on the adaxial and abaxial surfaces of filaments after flower opening [J]. Plant Syst Evol, 298(3): 561-567.
DUAN YA, LI QJ, 2008. The pollination biology of Phrynium oliganthum (Marantaceae) [J]. J Syst Evol, 46(4): 545-553. [段友愛, 李慶軍, 2008. 少花柊葉傳粉生物學的研究 [J]. 植物分類學報, 46(4): 545-553.
FENSTER CB, ARMBRUSTER WS, WILSON P, et al., 2004. Pollination syndromes and floral specialization [J]. Ann Rev Ecol Syst, 35(1): 375-403.
FETSCHER AE, 2001. Resolution of male-female conflict in an hermaphroditic flower [J]. Biol Sci, 268(1466): 525-529.
GAO JY, REN PY, YANG ZH, et al., 2006. The pollination ecology of Paraboea rufescens (Gesneriaceae), a buzz-pollinated tropical herb with mirror-image flowers [J]. Ann Bot, 97(3): 371-376.
GONG YB, HUANG SQ, 2007. On methodology of foraging behavior of pollinating insects [J]. Biodivers Sci, 15(6): 576-583. [龔燕兵, 黃雙全, 2007. 傳粉昆蟲行為的研究方法探討 [J]. 生物多樣性, 15(6): 576-583.]
HUANG SQ, 2007. Studies on plant-pollinator interaction and its significances [J]. Biodivers Sci, 15(6): 569-575. [黃雙全, 2007. 植物與傳粉者相互作用的研究及其意義 [J]. 生物多樣性, 15(6): 569-575.]
HUANG SQ, SHI XQ, 2013. Floral isolation in Pedicularis: how do congeners with shared pollinators minimize reproductive interference [J]. New Phytol, 199(3): 858-865.
JESSON LK, BARRETT SCH, 2002a. Solving the puzzle of mirror-image flowers [J]. Nature, 417(6890): 707.
JESSON LK, BARRETT SCH, 2002b. The genetics of mirror-image flowers [J]. Proc Biol Sci, 269(1502): 1835-1839.
JESSON LK, BARRETT SCH, 2002c. Enantiostyly in Wachendorfia (Haemodoraceae): the influence of reproductive systems on the maintenance of the polymorphism [J]. Am J Bot, 89(2): 253-262.
JESSON LK, BARRETT SCH, 2003. The comparative biology of mirror-image flowers [J]. Int J Plant Sci, 164(S5): S237-S249.
LI QJ, XU ZF, KRESS WJ, et al., 2001. Flexible style that encourages outcrossing [J]. Nature, 410: 432.
LI ZY, WANG YZ, 2005. Plants of Gesneriaceae in China [M]. Zhengzhou: Henan Science and Technology Publishing House. [李振宇, 王印政, 2005. 中國苦苣苔科植物 [M]. 鄭州: 河南科學技術出版社.]
LLOYD DG, SCHOEN DJ, 1992. Self and cross fertilization in plants: The selection of self-fertilization [J]. Intern J Plant Sci, 153: 370-380.
MAYFIELD MM, WASER NM, PRICE MV, 2001. Exploring the ‘Most Effective Pollinator Principle with complex flowers: bumblebees and Ipomopsis aggregate [J]. Ann Bot, 88(4): 591-596.
QU RM, LI XJ, LUO YB, et al., 2007. Wind-Dragged corolla enhances self-pollination: a new mechanism of delayed self-pollination [J]. Ann Bot, 100(6): 1155-1164.
REN MX, 2009. Floral syndrome and breeding system of Lobelia chinensis, a perennial herb with stamen fusion [J]. Chin J Plant Ecol, 33(2): 130-137. [任明迅, 2009. 雄蕊合生植物半邊蓮的花部綜合征與繁育系統 [J]. 植物生態學報, 33(2): 130-137.]
REN MX, ZHONG YF, SONG XQ, et al., 2013. Mirror-image flowers without buzz pollination in the Asian endemic Hiptage benghalensis (Malpighiaceae) [J]. Bot J Linn Soc, 173(4): 764-774.
ROALSON EH, ROBERTS WR, 2016. Distinct processes drive diversification in different clades of Gesneriaceae [J]. Syst Biol, 65(4): 662-684.
SCHOEN DJ, BROWN AHD, 1991. Whole-and part-flower self-pollination in Glycine clandestina and G. argyrea and the evolution of autogamy [J]. Evolution, 45(7): 1651-1665.
WEI YG, 2010. Gesneriaceae of South China [M]. Nanning: Guangxi Science and Technology Press. [韋毅剛, 2010. 華南苦苣苔科植物 [M]. 南寧: 廣西科學技術出版社.]
YANG SX, YANG CF, ZHANG T, et al., 2004. A mechanism facilitates pollination due to stigma behavior in Campsis radicans (Bignoniaceae) [J]. Acta Bot Sin, 46(9): 1071-1074.
YU Q, HUANG SQ, 2004. Flexible stigma presentation assists context-dependent pollination in a wild columbine [J]. New Phytol, 169(2): 237-241.
ZHANG DY, 2004. Plant life-history evolution and reproductive ecology [M]. Beijing: Science Press. [張大勇, 2004. 植物生活史進化與繁殖生態學 [M]. 北京: 科學出版社.]
(責任編輯 鄧斯麗 李 莉)
