中藥寡糖研究進展△
2023-06-06 13:52:56李加玲劉永建宋金楊洪柳王夢丹彭太昇張園園劉永剛
中國現代中藥 2023年4期
李加玲,劉永建,宋金,楊洪柳,王夢丹,彭太昇,張園園*,劉永剛*
1.北京中醫藥大學 中藥學院,北京 102488;
2.北京盛妝家化有限公司,北京 102488
中醫藥是我國特有的醫療衛生資源,是我國最具原始創新性的科技資源之一。近年來,在國家政策的推動下,中藥的創新或二次開發受到了高度關注。研究者在中藥藥效物質基礎領域開展了廣泛而深入的研究。
糖類是中藥中普遍存在的一類成分,隨著研究的不斷深入,寡糖因其所具有的多種藥理活性而受到廣泛關注。寡糖是由2~10 個單糖分子通過脫水縮合形成的糖苷鍵連接組成的直鏈或支鏈碳水化合物,由于其聚合度低、極性大、結構相似,混合物較難分離。研究者對寡糖分離進行大量研究,根據其性質開發了多種分離方法。目前,市場上多依靠性狀判斷藥材品質,經驗難以傳承。引入中藥藥效物質作為質量控制指標,結合性狀從定量、定性兩方面辨別藥材真偽、評價優劣,有利于建立全面的藥材質量標準。寡糖的糖苷鍵在外界條件變化時會發生水解,聚合度隨之改變,在高效液相色譜法(HPLC)圖上轉化為峰數量或峰面積的改變,此種信號容易識別,因此寡糖成分適合作為中藥的質量控制指標[1-2]。
本文通過檢索中國知網、萬方等數據庫中藥寡糖近5 年的研究成果,從制備方法、純化分離、藥理活性方面進行梳理,匯總了從中藥中得到寡糖的方式,歸納了純化分離寡糖提取液的方法及應用原理,介紹了寡糖類成分在地黃[3]、黃芪[4]、遠志[5-6]、巴戟天[7]質量控制方面的應用;概述了中藥寡糖抗氧化[8]、雙向調節免疫[9-11]、治療糖尿病及其并發癥[12-14]、抗阿爾茨海默病(AD)[15-17]、抗抑郁[18-19]等藥理作用;對中藥寡糖的分離和應用進行探討和展望,以期為其進一步開發提供參考。
1 寡糖制備方式
目前,研究者主要利用3 種方式從中藥中獲取寡糖:1)直接提取法。利用相似相溶原理,直接提取寡糖,但同時會溶出其他極性物質,如多糖、蛋白質。2)降解法。水解多糖得到對應的低聚糖片段,研究者已發現一些中藥多糖的藥理活性,但是為了解決相對分子質量大帶來的溶解性差、生物利用率低的問題,多采用水解多糖以降低其相對分子質量,同時又保留其活性。但由于多糖本身由單糖組成且糖苷鍵連接方式復雜,再加上分支結構的存在,降解所得到的混合產物的分離過程比較復雜,而且結構分析困難。3)合成法。根據已知結構合成寡糖。目前,寡糖合成難度很大,且中藥中寡糖活性實驗研究對象往往是寡糖混合物,因此這方面尚待補充。
1.1 直接提取法
根據相似相溶原理,可用水、乙醇、甲醇來提取一些中藥中的寡糖,這種直接用溶劑提取寡糖的方法稱為溶劑提取法。溶劑提取法操作簡單、適用性強,是提取寡糖的常用方法。考慮到傳統中藥多以水煎劑的方式服用,以及節約成本的原則,多以水為溶劑,水提醇沉法是相應的一種常用提取純化方案。通常是用熱水提取多次后,加乙醇至體積分數為80%,此時利用溶解度的差異使多糖沉淀得到寡糖溶液[20]。郝林華等[21]利用此法從牛蒡根中提出了水溶性的低聚果糖。有研究常直接采用醇提寡糖的方式,從而避免除多糖這一步操作[22]。
單純的溶劑提取法一般耗時長、溶劑消耗量大、產率較低。借助超聲波、微波可以縮短提取時間,提高效率。超聲波可以破壞細胞結構,使其在瞬間破裂釋放成分。錢艷艷等[23]提取鮮地黃中的寡糖,熱水法提取2 次,每次4 h;超聲法提取2 次,加上停頓時間,每次需要45 min。以葡萄糖為標準品,采用苯酚-硫酸法測定2 組低聚糖中碳水化合物質量分數分別為948.06、901.39 mg·g–1,且均有抗氧化活性。
1.2 降解法
多糖是由10 個以上單糖分子通過脫水縮合形成糖苷鍵,連接組成的天然直鏈或支鏈多聚糖是一些中藥的主要活性成分,具有抗氧化、調節免疫、降血糖和改善腸道功能等活性[24-27]。但多糖的高聚合度和復雜的分支結構限制了其進一步的研究和應用。現在多借鑒蛋白質“自上而下”的研究策略,將多糖水解為結構相對簡單的低聚糖,從低聚糖水平闡釋其結構特征[28]。常用的方法有酸降解法和酶降解法。
1.2.1 部分酸降解法 部分酸降解法是將多糖溶于無機酸(鹽酸、硫酸、磷酸)和有機酸[三氟乙酸(TFA)]中進行降解。其中,鹽酸酸性強、降解速度快,但降解程度難以把控,對環境的污染也比較嚴重。硫酸降解之后需要加碳酸鋇,避免影響質譜信號[29]。磷酸降解得到的產物均一,相比之下酸性較弱,反應周期長。TFA 是常用的1 種酸,揮發性好,減壓蒸發就可除去。使用TFA 降解多糖可以避免中和步驟,不會成鹽進而影響質譜信號、干擾結構分析。曹宇欣等[30]通過正交試驗得到了TFA 水解黃芪多糖的最佳實驗條件,水解完成后直接旋干就可除去TFA,簡單易行。
1.2.2 酶降解法 酶降解法是利用多糖降解酶特異性地、選擇性地切斷多糖內糖苷鍵,降低多糖聚合度,得到低聚糖的方法。與部分酸降解相比,酶降解反應條件溫和、速度快,還克服了化學降解片段相對分子質量分布范圍寬、均一性差的缺點。但是,酶本身對周圍環境很敏感,反應溫度、pH 等都能顯著地影響酶的活性。李科等[31]嘗試用內切α-1,4-葡聚糖酶酶解黃芪多糖,利用單因素試驗和正交試驗得到了反應過程中酶濃度、反應溫度、酶解時間三因素的最佳組合。在體外免疫活性篩查實驗中,聚合度為10~18時糖活性表現高于底物黃芪多糖。
1.3 合成法
寡糖分子中有多個羥基,空間構型復雜多樣,如何讓糖基供體和受體在特定位置形成一定構型的糖苷鍵就成為了難點。研究者多利用化學法和酶法來合成復雜寡糖,化學法靈活多樣,酶法高效。把2種方法結合起來,又是另外一種方法,即化學酶法。
1.3.1 化學法
1.3.1.1 糖基供體 糖基供體一般是在異頭位含有1 個在糖苷化條件下容易活化的離去基團,在其他位置上羥基均含有保護基的糖基砌塊。由于糖結構中存在多個羥基和縮醛結構,反應存在困難,因而通常先用離去基團來活化異頭位,制備成糖基供體,如糖基碘化物、糖基硫苷、糖基三氯乙酰亞胺酯、硝基烯糖等,再在促進劑作用下與糖基受體生成糖苷化產物[32]25-26,28。
Koenigs和Knorr把乙酰化葡萄糖與溴化氫反應,在異頭碳上引入了1 個溴原子。溴原子很容易離去,異頭碳變為碳正離子,很容易和其他親核試劑反應生成糖苷[32]36。之后化學合成法多沿用此思路制備糖基供體。
碘代糖是研究者基于溴代糖或氯代糖的制備方法得到的另一種糖基供體,可以大大提高產物的收率和立體選擇性,但是該供體也因活性太高,導致儲存不方便。硫代糖苷供體只在促進劑存在時發生反應,活化前非常穩定,并且產物構型靈活,故常用于多種復雜寡糖的合成。三氯乙酰亞胺酯供體是另外一種穩定、產物靈活的供體,制備過程相對簡單,在適宜堿存在的條件下,異頭位的裸露羥基與三氯乙腈反應即可得到。2 位硝基取代的烯糖供體在糖苷化反應中可以得到天然難以得到的α構型產物。其他的糖基供體還有很多,不過任何一種供體都不能適用于所有的糖苷化反應,所以糖基供體的研究也一直都是糖化學研究的重點。
1.3.1.2 糖苷化反應 1)Koenigs-Knorr 糖苷化法。Koenigs-Knorr 糖苷化法是指從α-鹵代糖出發,在銀鹽的催化下產生β-苷,某些Lewis 酸和相轉移催化劑也可催化[33]。反應產物純度高、收率高,但反應條件苛刻,需避光、無水。中間體乙酰溴代糖具有較好的離去性能,同時穩定性很差,極易水解。但該法開辟了化學合成糖苷的先河,在糖化學中占有重要的地位[34]。2)Fischer糖苷化法。Fischer糖苷化法是低分子糖在酸性條件下與簡單醇的反應,反應可逆、操作簡單,但糖基供體(游離糖或半縮醛)活性低,因此只可用結構簡單的活潑醇作為糖基受體。
1.3.1.3 寡糖合成策略 1 個完整的寡糖分子的合成路線往往需要很多步反應。傳統的分步法需要反復分離反應中間體,對中間體施加保護基,不僅耗時耗力,收率還很低。“一鍋法”在有機合成中應用較多,在合成寡糖時,將反應所需的糖基砌塊按照一定的順序加入1 個反應容器中,待前一步糖苷化反應結束后再加入下一步反應的砌塊,與前一步反應的產物進行偶聯,如此反復進行,直至生成最終產物后進行純化,得到目標寡糖。反應過程中需要選擇合適的中間體來確保反應按照設計路線進行,可以基于化學選擇性、正交選擇性來合成糖基砌塊供體,要求供體之間有足夠大的活性梯度差異,或者離去基團不同。而“預活化法”不要求這些,該法在加入糖基受體之前,先用促進劑將容器內的糖基供體或上一步的產物活化,加入受體后就可以立即偶聯[32]41-44。
1.3.2 酶法 酶法合成中主要用到糖基轉移酶、糖苷酶、糖合成酶3 種酶。糖基轉移酶中應用較多的是Lelori 轉移酶,其將核苷酸活化的糖基供體轉移到合適的糖基受體上,反應的區域選擇性和立體異構性好,但糖基供體較為昂貴。糖苷酶本身可以切割糖苷鍵,但是在體外,通過控制反應條件,糖苷酶反而可以合成糖苷鍵。該酶可以通過逆水解和轉糖基2 種途徑,利用游離糖或簡單修飾的糖基供體來合成寡糖。逆水解途徑中,糖苷酶可以讓游離糖和受體羥基直接縮合,但反應的收率比較低。因此轉糖基反應在實際中使用較多。同化學合成反應相比,轉糖基反應不需加入促進劑,糖苷酶可將經過修飾的糖基供體和受體縮合成苷。但是由于糖苷酶自身潛在的水解活性,有時反應的產率仍然會過低。前2 種酶雖是自然界中存在的,但其推廣應用都受到了一定的限制,而糖合成酶是利用基因工程和蛋白質工程,對現有的糖苷酶進行改造而來,其糖基轉移活性大大提高[32]49-53。
2 寡糖純化分離工藝
寡糖混合物常利用彼此之間相對分子質量、極性、電荷的差異,通過液相色譜法、電泳法、膜分離法等進行純化分離,并根據需要選擇合適的檢測器。寡糖純化分離方法比較見表1。

表1 寡糖純化分離方法
2.1 凝膠色譜法
凝膠色譜法中,凝膠在柱中相當于分子篩,起到讓大分子組分先流出、小分子組分后流出的作用,因此使用這種填充劑的柱色譜法又被稱作分子排阻色譜法或分子篩色譜法,此法常使用的填充材料有聚丙烯酰胺凝膠、葡聚糖凝膠等,以水作洗脫液,操作簡便、樣品回收率高。聚丙烯酰胺凝膠由單體丙烯酰胺與交聯劑甲叉雙丙烯酰胺交聯而成,具有不同型號的商品。李科等[31]通過P-2凝膠柱分離黃芪多糖降解產物,可以得到4 組寡糖片段,分別為聚合度1~3 糖、3~6 糖、7~14 糖、10~18 糖。葡聚糖凝膠也有不同規格,串聯使用不同孔徑的凝膠色譜柱可以達到更好的分離純化效果。許定舟等[35-36]水提醇沉麥冬得寡糖粗液后,先經Sephadex G-75柱色譜脫色脫蛋白,寡糖質量分數為97.64%,樣品的顏色也變為純白色,再采用Sephadex G-15柱色譜對麥冬寡糖進行分級分離,出現2個洗脫峰,最終得到2種相對分子質量的麥冬寡糖。
2.2 親水作用色譜法(HILIC)
較復雜糖類化合物在傳統的反向液相色譜柱上難以保留,也很難實現有效分離。目前主要采用2種策略:一是對糖類樣品進行衍生化,在結構上引入生色團或熒光團,降低寡糖的極性,從而提高分離效率和檢測信號;二是使用HILIC,以極性分離材料為固定相、水溶性有機溶劑為流動相,極性越大的物質在色譜柱上的保留越強,流出順序與傳統的正向色譜柱相同,常選擇示差折光檢測器(RID)或蒸發光散射檢測器(ELSD)。
RID 通過檢測參比池與測量池中折射率之差來檢測寡糖含量,不需對寡糖進行衍生化處理,但是對壓力、流動相組成及流速的變化都較敏感,因此只能使用等度洗脫,并且該檢測器靈敏度不高。邱建國等[37]利用HPLC-RID 測定了水蘇糖成分含量。與RID 相比,ELSD 靈敏度較高[38],其要求使用具有揮發性的緩沖鹽及流動相,可以消除溶劑的干擾,基線穩定、沒有折光視差效應、使用梯度洗脫靈敏度更高。孫恬等[39]考察色譜柱和流動相種類對巴戟天寡糖的分離效果影響時發現,XBridge Amide色譜柱(250 mm×4.6 mm,3.5 μm),流動相為0.2%三乙胺乙腈溶液(A)-0.2%三乙胺水溶液(B)梯度洗脫時對各成分的分離效果最好。
2.3 高效陰離子交換色譜法(HPAEC)
在堿性條件下,糖鏈上的羥基可以形成氧負離子。不同寡糖中的羥基的電離常數是有細微差別的,導致陰離子交換樹脂和氧負離子的相互作用產生差別。HPAEC 根據這種原理,利用強堿洗脫劑導致不同寡糖出現在不同的洗脫位置,從而對其進行分離,一般與脈沖安培檢測器(PAD)聯用,不需衍生、簡便易行、專屬性強。由于洗脫劑為強堿,儀器耐堿性要求高。強堿往往會導致糖的異構化,所以需要在短時間內迅速分離來減少對寡糖結構的影響。
馬務迢等[40]為考察巴戟天不同生長期中的寡糖類成分含量變化規律,采用 RCX-10 色譜柱分離寡糖溶液,柱材料為316L 不銹鋼和聚醚醚酮,屬于耐堿材料,以100 mmol·L–1NaOH 溶液作為A 液,100 mmol·L–1NaOH+500 mmol·L–1NaOAc混合溶液作為B 液,組成流動相進行梯度洗脫,滿足了HPAEC-PAD對于色譜柱和流動相的要求。色譜圖反映葡萄糖、果糖、蔗糖、1-蔗果三糖、耐斯糖、1F-果呋喃糖基耐斯糖6 種寡糖成分與相鄰色譜峰之間的分離度均大于1.5,說明分離效果好。
2.4 毛細管電泳法(CE)
CE 是以毛細管為分離通道,高壓直流電場作為驅動力,具有樣品需求量少、靈敏度高、分離效率高的優點,應用于糖類分析的主要有毛細管區帶電泳(CZE)、毛細管凝膠電泳(CGE)及膠束電動毛細管電泳(MEKC)。
CZE 中寡糖需要帶電,方法包括在硼酸鹽緩沖液中與硼酸根絡合,在強堿緩沖液中電離,進行衍生化,接著帶電寡糖在緩沖液中和電場作用下因淌度不同而實現分離。CZE 是電泳法中相對簡便,應用廣泛的一種分離方法。孫燕[41]在降解褐藻膠時發現,酶降解所得寡糖在非還原端碳4、5 位形成雙鍵與碳6 位的羧基中的羰基共軛,在235 nm 處具有特征紫外吸收,于是不經衍生,直接經高效毛細管電泳-紫外分光檢測方法(HPCZE-UV)分離檢測了酶降解產物;酸降解所得寡糖無此共軛體系,并沒有特征紫外吸收,也沒有熒光發光基團,如果直接分析檢測就比較困難,便將酸降解寡糖與8-氨基芘-1,3,6-三磺酸鈉(APTS)熒光衍生化試劑反應,并優化了衍生化條件和電泳條件,經HPCZE-激光誘導熒光檢測法(LIF)實現了高效分離,檢測限達到了1×10–17mol水平。
CGE 是從區帶電泳中派生出來的把凝膠作為毛細管中支持物的電泳方式。一方面,凝膠在管中起到分子篩的作用;另一方面,由于凝膠黏度大,又可以減少溶質的擴散。MEKC 是通過在緩沖液中加入表面活性劑,使其形成膠束作為準固定相。中性物質在水相和該相的分配系數不同而得到分離,達到與CZE互補的效果,擴大電泳法的分離范圍。
2.5 熒光輔助糖電泳法(FACE)
FACE是通過衍生化反應使糖還原端結合上熒光基團,然后在一定濃度的聚丙烯酰胺凝膠上進行分離的一種分析方法,可以根據電泳條帶數量及熒光強度來判斷出寡糖的成分及相對含量,直觀易讀,具有高分辨率和高靈敏度。目前,有若干種衍生方法和相應的衍生試劑,其中8-氨基萘基-1,3,6-三磺酸(ANTS)應用較多,不僅為糖分子提供了熒光基團,還提供了3 個負電荷,使原來的中性糖分子帶電,有足夠的動力在電場力作用下進行電遷移。姚喜梅等[42]從ANTS、2-氨基苯甲酸(2-AA)和2-氨基吖啶酮(AMAC)中選擇葡甘露寡糖的熒光衍生劑時,發現ANTS 組條帶更加清晰且數量更多,因此確定ANTS 為實驗熒光衍生劑,并從分離膠濃度、ANTS 用量、衍生化反應溫度、衍生化反應時間等方面對衍生條件進行了優化。李曉霞等[4]將黃芪多糖經內切-1,4-β-半乳聚糖酶降解寡糖同樣用ANTS 衍生,通過FACE 獲得糖指紋圖譜,結合主成分分析,找出了區分不同種質資源黃芪的差異性糖組分。
2.6 石墨化碳液相色譜法
石墨化碳液相色譜法的固定相是多孔石墨化碳(PGC),PGC 是由sp2 雜化的碳原子呈六邊形排列,通過共價鍵相連構成層,層間通過范德華引力相互作用結合在一起構成帶,石墨帶再交織而成。PGC主要是利用分子結構和分子極性的不同進行分離。分子結構越平,其與石墨化碳的結合就越緊密。具有大極性基團的分子可以使石墨化碳表面極化,進而導致兩者之間作用力加強,提高分離效率。寡糖分子中具有多個羥基,復雜的寡糖異構體很適合用這種方法分離。
2.7 高速逆流色譜法(HSCCC)
HSCCC 是一種高效快速的液-液分配色譜,固定相和流動相都是液體。兩相互不相溶,在高效運動的螺旋管內建立了單向性流體動力學平衡。樣品中各組分由于在兩相中分配系數存在差異而實現分離,在流動相中的分配比例大的組分先被洗脫下來。沒有固相載體,避免了不可逆吸附而帶來的損失,基本上能回收全部樣品。
符穎[43]先用分析型HSCCC 篩選出乙酸乙酯-正丁醇-水(4∶1∶5)最佳溶劑系統,再放大柱體積,采用制備型HSCCC,重新考察流速、轉速等條件,摸索不同上樣量對分離效果的影響,以建立合適的制備方法。最后成功從400 mg 遠志樣品中分離得到3,6?-二芥子酰基蔗糖酯(DISS)和遠志蔗糖酯A,HPLC檢測純度分別為93%、95%。
2.8 膜分離法
膜分離法依據相對分子質量截留值分為微濾膜、超濾膜和納濾膜。超濾膜能夠除去蛋白質、多糖等大分子物質,納濾膜能夠除去半乳糖等小分子物質,從而得到高純度的寡糖混合物,操作比較簡單,過程中沒有化學變化。楊菁等[44]用超濾膜除去地黃提取液中相對分子質量>5000 的物質,收集超濾液后再利用活性炭脫色。但是一般會使用不同膜分級分離。如果截留值差距較大,得到的以某一相對分子質量范圍為主的組分中往往會存在相對分子質量或高或低的組分。
3 寡糖的質量控制應用
寡糖是中藥中新發現的成分,其聚合度低、空間結構獨特、具有較好的藥理活性,為人們尋找新的中藥質量指標提供了思路。研究者為此開展了一系列研究工作,試圖在中藥材種植采收、炮制工藝中找到獨特的寡糖成分,以期將其作為中藥材質量控制指標。
3.1 地黃
地黃中含有蔗糖、棉子糖、甘露三糖、水蘇糖等寡糖成分[45]。張汝學團隊研究地黃多年,優化地黃寡糖提取分離工藝,考慮到生地黃藥材傳統服藥方式,選擇水提寡糖,在之后發現不同產地的生地黃和熟地黃寡糖成分類型和含量都存在較大差異[46-48]。結合HPLC 圖發現,在蒸法炮制熟地黃的過程中,水蘇糖和棉子糖均失去1 個果糖而變成相應的甘露三糖和楝二糖。并且蒸制時間達到10 h,達到《中華人民共和國藥典》(以下簡稱《中國藥典》)2010 年版規定熟地黃性狀要求時,已檢測不到水蘇糖、棉子糖和梓醇,同時甘露三糖、楝二糖及單糖的含量明顯增加[1]。同樣利用HPLC 觀察另一種炮制方法——酒燉法,依靠傳統辨別經驗“燉至酒盡”,燉4 h,發現也未達到《中國藥典》2010 年版規定性狀,根據HPLC 圖顯示,水蘇糖、棉子糖和梓醇也均沒有完全消失,甘露三糖和楝二糖的增幅也不大[3]。傳統炮制依據“蒸治黑潤”或“燉至酒盡”來肉眼判斷是否炮制成熟,這種經驗不易傳授,而且藥材質量無可避免地存在差異,會影響臨床療效。如果利用HPLC 來指導炮制進程的話,容易判斷是否炮制成熟,準確度高。
3.2 黃芪
黃芪是一種傳統補益藥,本身用量大,加上野生資源枯竭,多地進行人工引種,栽培1 年便采收。這種栽培方式得到的黃芪稱為速生黃芪,性狀與野生黃芪大為不同。關于速生黃芪品質好壞存在很大爭議,為找到可以全面準確評價野生黃芪和移栽黃芪品質的指標,“多糖受體”學說推測多糖分子中存在一種或多種寡糖片段的“活性中心”[49]。李曉霞等[50]模擬人工胃液條件處理黃芪細胞可溶性多糖及糖綴合物將降解得到不同聚合度的寡糖,證明黃芪多糖經口服后也主要以寡糖形式顯效。李曉霞等[4]繼續利用內切-1,4-β-半乳聚糖酶水解了蒙古黃芪(野生芪和移栽芪)與膜莢黃芪(野生芪和移栽芪)的可溶性多糖,通過FACE 獲得糖指紋圖譜,結合主成分分析法,發現酶解產物中的五糖、六糖可以作為區分蒙古黃芪的移栽芪和野生芪的差異性糖片段,而四糖、五糖可以作為區分膜莢黃芪的移栽芪和野生芪的差異性糖片段。王迪等[51]發現,內切-1,3-β-葡聚糖酶酶解產物寡糖中五糖和六糖可以作為區分膜莢黃芪(野生芪和移栽芪)的差異性糖片段。不過,并未提及差異性糖片段的活性。曹宇欣[52]通過水提醇沉、超濾截留從山西仿野生黃芪中得到3 組不同相對分子質量的多糖,其中,相對分子質量約為10的黃芪多糖APS-Ⅱ,以葡萄糖作為主要單體,在體內外免疫活性篩選實驗中活性最強。李科等[31]利用內切α-1,4-葡聚糖酶降解APS-Ⅱ,得到了免疫活性強于APS-Ⅱ的聚合度10~18糖混合物。
3.3 遠志
遠志來源于遠志科植物遠志的根,其中的寡糖酯類成分是一類主要以蔗糖為共同母核,以不同形式的糖苷鍵連接葡萄糖或鼠李糖(較少)成為寡糖后,再與有機酸(乙酸、苯甲酸或苯丙烯酸)類成分形成的酯,其是不超過五分子糖的一類化合物[53],其中,3,6′-二芥子酰基蔗糖是《中國藥典》2020 年版評價遠志藥材質量的標志性成分。Wu 等[54]發現可以選擇超高效液相色譜-質譜法(UPLC-MS)中鑒定的遠志寡糖酯進行質量評價。研究者一般常通過測定多個而非單一寡糖酯類成分來反映遠志藥材的內在品質。相關研究比較了遠志根皮(生品)與遠志木心中西伯利亞遠志糖A5、西伯利亞遠志糖A6、球腺糖A、遠志蔗糖酯B、3,6′-二芥子酰基蔗糖、黃花遠志素、遠志蔗糖酯A 及遠志蔗糖酯C 的含量[5-6],發現根皮中該類成分含量均明顯高于木心,從而證明了古代遠志去木心的科學性。另外,遠志經甘草汁煮后,上述寡糖酯除球腺糖A 外含量均降低。宋夢晗等[55]還發現,在此過程中芥子酸、對羥基苯甲酸、對香豆酸、阿魏酸等8 種有機酸含量均顯著增加。相關研究采用HPLC-飛行時間質譜法(TOF/MS)和已知對照品模擬炮制方法,模擬遠志經水煮、甘草汁煮的過程,確證遠志中寡糖酯類成分性質不穩定,在甘草水煮制過程中發生了酯鍵水解和構型改變[56-57]。考慮到寡糖酯類在對照品藥材中含量低、極性大、不易制備,張曉玲等[58]選用遠志中含量高、易制備的DISS作為內參物,引入一測多評法,實現了多種糖酯成分的同步測定,該方法成本低、準確度高。
3.4 巴戟天
巴戟天中低聚糖成分主要為菊粉型低聚糖(GFn),是由(1→2)-β-D-呋喃果糖苷鍵結合而成的線性直鏈型低聚糖,其一端是蔗糖,糖鏈增長基團為β-呋喃型果糖,末端帶有1 個葡萄糖殘基,聚合度通常為2~60,聚合度雖增加,增長的基團和連接方式不變[59]。馮峰[60]從巴戟天中分離得到了3個全由果糖組成的果聚糖。
《中國藥典》2020 年版規定巴戟天藥材里耐斯糖不得低于2.0%[61]。在傳統的中藥材交易中,按照“辨狀論質”理論,巴戟天以條粗、肉肥厚而色紫藍、味甜、木心細者質佳[62]。楊麗等[63]利用超聲醇提法從生巴戟天樣品得到低聚糖溶液,經HILICELSD 分析發現,巴戟天根直徑越小斷面越紫、木心越細,蔗果三糖、耐斯糖、蔗果五糖含量越高,與傳統理論部分不一致,值得思考。一些研究者注意到巴戟天寡糖的活性,將其作為考察巴戟天藥材的栽培年限、儲存條件、炮制規范時的指標。楊麗等[64]繼續考察巴戟天生長年限與巴戟天寡糖含量的關系,在廣東省德慶縣巴戟天規范化種植基地采集樣品,發現5 年生的巴戟天中耐斯糖與蔗果五糖含量達到最高,4 年生蔗果三糖含量最高,考慮到藥用價值、種植成本和土地利用效率等因素,建議巴戟天的最佳栽培年限為4~5 年。冼麗鏵等[65]在廣東省德慶縣使用隨機多點采樣法,采集了不同生長年限的巴戟天樣品,同樣發現4 年生巴戟天的蔗果三糖含量最高,并且顯著高于2年生巴戟天,但4年生與5 年生的耐斯糖和蔗果五糖含量無明顯差異,因此認為4年生的巴戟藥材質量最佳。孫恬等[66]繼續考察采后儲存溫度和時間對巴戟天藥材中寡糖含量影響,發現在25 ℃有水分存在的條件下,巴戟天寡糖會發生水解反應,導致GF2、GF3、GF4的減少,而D-果糖、葡萄糖、蔗糖增加,因此建議60 ℃下烘干或4 ℃下低溫保存藥材。相關研究建立了GF2~GF11的含量測定方法[67-68],比較飲片巴戟天、巴戟肉和鹽巴戟天中單糖和低聚糖含量的差異,發現巴戟肉和鹽巴戟天中10 種低聚糖GFn(n=2~11)的含量均分別低于飲片巴戟天中相應成分的含量(P<0.01),其中,多批巴戟肉中耐斯糖含量低于《中國藥典》2020 年版中限量要求,這為飲片巴戟天、巴戟肉、鹽巴戟天的質量標準修訂和提升提供了參考。
4 寡糖的藥理活性
現代研究表明,中藥寡糖在抗氧化、調控免疫系統,治療生殖障礙、糖尿病、AD 和抑郁癥(MDD)等方面都有貢獻。
4.1 抗氧化
在生理情況下,人體內的自由基處于不斷產生與清除的動態平衡中。有時受到來自機體自身活動、情志因素、外界因素干擾,平衡狀態遭到破壞。自由基清除不及時,就會使機體組織和細胞受到損害,繼而造成多種疾病的發生。中藥寡糖具有良好的抗氧化活性,可以直接清除自由基,提高抗氧化酶的活性。
中藥寡糖對各種自由基具有清除能力。于冰莉等[8]發現,甘草地上部分寡糖在體外可劑量依賴性地清除1,1-二苯基-2-三硝基苯肼(DPPH)自由基,并可以延長線蟲在H2O2應激環境中的壽命。錢艷艷等[23]研究不同提取方法對鮮地黃寡糖的影響時發現,超聲法和熱水提取法2 種方式得到的低聚糖片段對超氧陰離子自由基和DPPH 自由基均表現出一定的清除能力。金釵石斛低聚糖對2,2-聯氮-二(3-乙基-苯并噻唑-6-磺酸)二銨鹽(ABTS+·)自由基的清除能力優于對DPPH 自由基的清除能力[69]。一些中藥寡糖表現出優于多糖的自由基清除能力。冀建偉等[70]以2,6-二叔丁基-4-甲基苯酚(BHT)為陽性對照,發現厚樸花中低聚糖的體外抗氧化活性優于多糖組分,這可能與兩者結構差異有關。馬銘等[71]發現,黨參中粗低聚糖的抗氧化活性強于多糖,接著用Sephadex G-25 柱純化粗低聚糖,發現在質量濃度均為5.00 mg·mL–1時,精制黨參低聚糖(CPO)的DPPH自由基清除活性強于粗低聚糖。
中藥寡糖還可以通過調控機體酶抗氧化系統,增強抗氧化酶活性起到抗氧化作用。抗氧化酶包括超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、谷胱甘肽還原酶(GR)及谷胱甘肽過氧化物酶(GSH-Px)。CPO 在體外顯示出極強的抗氧化活性[71]。周靜[72]利用D-半乳糖(D-gal)制備大鼠衰老模型進行體內實驗發現,CPO可以提高血清中CAT、GSH-Px 和SOD 活性,并且降低脂質氧化變性后產物丙二醛(MDA)的水平,改善了D-gal 誘導的活性氧產生過剩引發的氧化應激。程亮星[73]發現巴戟天寡糖可以升高抗氧化酶水平,降低MDA和過氧化脂質的水平,發揮抗氧化作用,保護子宮組織。
4.2 調節免疫
人體在免疫低下的狀態,如果受到了外界有害因素的侵襲,就容易發生疾病。機體會自發產生炎癥免疫反應(IIR),進行自我保護和修復。但是過度的IIR也會損害機體自身。市面上已有的一些抗炎藥雖能起到抗炎作用,但同時會產生一系列不良反應,對機體器官造成損傷。研究發現中藥寡糖既可以抵抗免疫低下,增強機體免疫能力,又可以控制炎癥反應。一方面,中藥寡糖可以通過非特異性免疫和特異性免疫2 種途徑,提高巨噬細胞吞噬功能,促進細胞因子的分泌,加強抗體的生產,起到增強免疫的作用。另一方面,寡糖也能從核轉錄因子(NF)-κB 和促分裂原活化蛋白激酶(MAPKs)2 種途徑,通過阻止細胞因子的產生與釋放抑制環氧化酶(COX)和一氧化氮合成酶(NOS)等酶活性起到控制IIR的作用。
環磷酰胺是一種烷化類的免疫抑制劑,可殺傷淋巴細胞,抑制特異性抗體反應。劉少芳[9]利用環磷酰胺制備免疫低下小鼠模型,灌胃牛蒡寡糖(BFO),發現BFO 可提高巨噬細胞的吞噬功能,促進脾細胞增殖,顯著提高血清溶血素的生成,顯著增多B 細胞介導的溶血反應,這說明BFO 可以增強免疫低下小鼠的非特異性及特異性免疫能力。盛榮華等[10]研究發現BFO 還可以顯著促進正常小鼠脾細胞分泌細胞因子白細胞介素-2(IL-2)和γ-干擾素(IFN-γ)。
劉晶[11]用BFO(純度>95%)粉劑配置成高、中、低劑量組的BFO溶液,利用脂多糖(LPS)誘導小鼠巨噬細胞RAW264.7制備細胞炎癥模型,檢測細胞因子轉錄翻譯水平,測定NF-κB和MAPKs通路蛋白磷酸化水平,結果發現BFO可顯著抑制LPS上調的IL-6、單核細胞趨化蛋白-1(MCP-1)和腫瘤壞死因子(TNF)-α轉錄水平和蛋白表達水平,同時抑制誘導型COX-2及一氧化氮合酶(iNOS)的表達,并且抑制NF-κB p65 和κB 抑制因子(IκB)磷酸化水平及細胞外信號調節激酶(Erk)、p38、c-Jun 氨基末端激酶(JNK)磷酸化水平,因此推測BFO 可能是通過抑制模型細胞中NF-κB和MAPKs通路降低細胞因子的活性,從而發揮抗炎活性。
CPO 也具有類似BFO 的雙向免疫調節、抵抗免疫低下和抗炎作用。馬銘[74]制備環磷酰胺小鼠免疫低下模型,灌胃CPO(50、100、150 mg·kg–1),發現CPO 可顯著提高小鼠巨噬細胞吞噬活性,增強小鼠的遲發型變態反應的耳廓腫脹程度,促進脾淋巴細胞增殖和轉化,顯著提高細胞因子IL-2 和IFN-γ、NO 的分泌水平及血清免疫球蛋白G(IgG)、血清免疫球蛋M(IgM)的含量,發揮增強免疫的作用,抵抗環磷酰胺導致的免疫低下。周靜[72]制備衰老大鼠模型,增加了給藥劑量(230、400、700 mg·kg–1),繼續研究CPO 的免疫活性,發現CPO 此時可以阻止衰老大鼠血清炎癥因子TNF-α、IL-6、IL-1β的過度表達,抑制NF-κB 和MAPKs 通路的活化,改善D-gal引起的炎癥。由于所用動物品種及實驗指標不同,尚無法確定CPO的雙向調節作用機制。
4.3 調節血糖
糖尿病是全球高發內分泌系統疾病,常累及其他器官,如肝臟和腎臟。隨著該疾病的發生,患者體內會出現持續有高血糖及糖脂代謝紊亂的狀況,包括膽固醇(TC)、三酰甘油(TG)、低密度脂蛋白膽固醇(LDL-C)的增加和高密度脂蛋白膽固醇(HDL-C)的減少。血管內皮生長因子(VEGF)是一種高度特異性的促血管內皮細胞生長因子,可因高血糖引起組織缺氧而被激活表達。糖脂代謝紊亂和氧化應激會進一步損傷機體器官,導致并發癥的產生。中藥寡糖可改善糖尿病的高糖癥狀,中藥寡糖對糖尿病并發癥也有一定的作用。
4.3.1 降血糖 張汝學等[75-77]對地黃寡糖(RGOS)的降血糖活性進行了系統的研究,發現RGOS 不僅可以調節實驗性糖尿病的糖代謝紊亂和生理性高血糖狀態,還可以逆轉去胸腺大鼠和老年大鼠的異常糖代謝,但對正常大鼠無作用。胰島素抵抗是多種代謝相關疾病的共同病理基礎和危險因素,是2 型糖尿病發病的中心環節。郭麗民等[78]采用高胰島素建立HepG2 細胞株胰島素抵抗模型,發現地黃寡糖可以增加模型對葡萄糖的消耗。
α-葡萄糖苷酶是人體內關鍵的淀粉消化酶之一,活性物質可通過抑制該酶的活性從而減緩葡萄糖的釋放速度,避免血糖大的波動。體外實驗表明相對分子質量為500~1500 的麥冬寡糖可以顯著抑制α-葡萄糖苷酶活性[36]。以與人類2型糖尿病發病較為相似的自發性2 型糖尿病db/db 小鼠為模型,灌胃給予麥冬寡糖,發現麥冬寡糖可以改善小鼠表現的肥胖、高血糖癥、血脂異常和腎損傷[79]。
4.3.2 抗糖尿病肝病、腎病 糖尿病性腎病是糖尿病的主要并發癥之一。陳海蘭等[12]利用高脂飼料聯合鏈脲佐菌素(STZ)制備糖尿病大鼠模型,灌胃ROS(200 mg·kg–1)22 d,觀察血脂代謝和腎臟組織中VEGF 情況,結果顯示,ROS 降低了TC 和TG水平,升高了HDL-C 水平,并且TG 降低效應比陽性對照藥二甲雙胍更明顯,一定程度上逆轉了糖脂代謝紊亂;其還降低了腎臟質量及腎臟指數,改善了腎臟組織結構,一定程度上修復了腎臟組織,同時減少了VEGF 蛋白相對表達量。因此推測ROS 改善糖尿病腎病的作用與抑制VEGF的表達有關。
糖尿病性肝病、生殖功能障礙和性功能減退也是糖尿病的常見并發癥。持續高血糖及糖脂代謝紊亂會導致脂肪肝和氧化應激的出現與發生,氧化應激又會進一步損傷肝臟組織和睪丸組織。侯慶華等[13-14]以高脂飼料聯合STZ 法制備2 型糖尿病大鼠模型,灌胃給予昆布寡糖8 周,考察其對糖尿病大鼠肝臟和睪丸的保護作用。8 周后,糖尿病大鼠血糖水平降低、胰島素水平升高,表明糖脂代謝紊亂得到逆轉;其肝臟中天冬氨酸氨基轉移酶(AST)和丙氨酸氨基轉移酶(ALT)水平降低,血清中睪酮含量升高,肝臟和睪丸組織得到修復,且肝臟組織和睪丸組織中MDA含量顯著降低,SOD、CAT活性顯著升高,表明昆布寡糖在一定程度上抵抗了氧化應激。因此,昆布寡糖對糖尿病大鼠的肝臟和睪丸具有保護作用。并且,大鼠睪丸組織中VEGF 表達減少,推測昆布寡糖對糖尿病睪丸的保護作用可能也與抑制VEGF有關。
4.4 改善生殖功能
勃起功能障礙(ED)是臨床常見的男性性功能障礙疾病,表現為陰莖持續不能達到或維持足夠的勃起以完成滿意的性生活。中藥寡糖,如BFO、巴戟天寡糖,有改善生殖功能的作用。
孟宇等[80-81]發現,BFO可以促進體外培養的大鼠陰莖海綿體平滑肌細胞增殖,繼續通過大鼠雙側髂內動脈結扎術來制備動脈性勃起功能障礙(A-ED)模型,實驗結果表明,BFO 可以顯著上調動脈結扎模型中神經型一氧化氮合酶(nNOS)和內皮型一氧化氮合酶(eNOS)基因的表達,影響NO 的合成。NO作為一種神經遞質,在平滑肌松弛的信號傳導通路中發揮著重要作用。說明BFO 通過影響NO 的合成,影響NO-環磷酸鳥苷酸(cGMP)通路,介導平滑肌松弛,促使陰莖勃起,來改善ED。趙君等[82]采用“勞倦過度、房事不節”法制備小鼠腎陽虛模型,發現巴戟天寡糖一邊調節垂體-下丘腦-性腺軸從而調節生精過程,一邊發揮抗氧化作用,保護睪丸精子免受氧化損傷。
4.5 改善學習記憶能力及抗AD
AD是一種慢性神經退行性疾病,主要臨床表現包括記憶力逐漸減退、認知功能發生障礙、行為異常和社交障礙等。病理特征主要為老年斑(SP)、神經元纖維纏結(NFT)及神經元數量減少[17]。近現代多從膽堿能系統障礙、神經細胞凋亡、氧化應激、神經炎性病變等角度進行研究并尋找有效藥物。目前,美國食品藥品監督管理局(FDA)批準的5 種抗AD 藥物屬于對癥治療,影響AD 進程的藥物仍處于初步研究階段。中藥在治療AD 疾病方面有著悠久的歷史和豐富的經驗,近年來國內外眾多學者從中藥及其有效成分著手,針對治療AD 的天然藥物進行了廣泛的研究,如人參水溶性寡糖、遠志寡糖酯成分。
孫麗麗等[15-16]采用東莨菪堿(scop)誘導癡呆小鼠,發現水溶性人參寡糖(WGOS)能夠改善小鼠模型的空間學習記憶能力,同時能夠抑制小鼠海馬組織中乙酰膽堿酯酶(AchE)的活性。許婷[83]試圖從炎癥反應角度探討WGOS 的神經保護作用機制,結果再次證實WGOS 可顯著改善scop 誘導癡呆小鼠的學習記憶能力,同時還抑制海馬組織及血清中IL-1β、TNF-α的表達和海馬組織中星形膠質細胞的激活。因此推測WGOS 是通過抑制中樞系統的炎癥反應進程,起到抗AD的作用。
遠志是一種傳統類益智藥,具有安神益智、交通心腎、祛痰、消腫的功效,用于心腎不交引起的失眠多夢、健忘驚悸等。遠志寡糖酯類成分具有神經保護、抗氧化應激、調整神經遞質水平等藥理活性,提示這類成分具有抗AD 的潛能。研究發現,富含50%寡糖酯類成分的遠志醇提物可以減少AD模型小鼠海馬區大腦皮層的NFT 和SP,促進大腦皮層神經元生長,改善AD 藥理癥狀[17]。相關研究發現,遠志寡糖酯可以改善氫溴酸東莨若堿所誘導的小鼠學習記憶障礙,從β-淀粉樣蛋白(Aβ)毒性假說角度探討遠志寡糖酯抗AD 作用機制,發現其可以對抗Aβ25-35 誘導的SH-SY5Y 細胞損傷,減少細胞凋亡,激活蛋白激酶B/環磷酸腺苷反應元件結合蛋白/腦源性神經營養因子(Akt/CREB/BDNF)信號通路,推測是通過正反饋加強了BDNF 的神經保護作用[84-86]。
4.6 抗抑郁
MDD 是現在比較常見的一種心理疾病,以長期連續的情緒低落為主要的臨床特征,屬中醫學“郁證”范疇。傳統中醫學對郁證有較為系統的理論認識及豐富的診治經驗,在MDD尤其是輕中度抑郁的防治中可以發揮積極作用,寡糖在其中起了重要作用,如遠志寡糖酯、巴戟天寡糖。
遠志醇提物經大孔吸附樹脂上梯度洗脫的過程中,經50%乙醇洗脫得到的活性部位被稱為YZ-50。YZ-50 中富含寡糖酯類成分,在小鼠強迫游泳實驗(FST)和小鼠懸尾實驗(TST)中表現出抗抑郁效果。由于經典單胺理論認為,MDD 的發生與腦內單胺遞質,尤其是去甲腎上腺素(NE)和5-羥色胺(5-HT)功能不足有關,于是進一步從神經遞質方面深入研究YZ-50 抗抑郁機制,發現其能顯著增強育亨賓的毒性,并且抑制阿撲嗎啡所引起的小鼠體溫下降,由此推測遠志通過阻斷單胺類遞質的重攝取從而發揮抗抑郁效果[18]。從YZ-50中分離出DISS,發現DISS 同樣具有抗抑郁活性,可以增強5-HT 和NE神經功能[87]。
結合慢性不可預見性應激和孤養制備大鼠慢性應激抑郁模型,進一步探究YZ-50 在神經內分泌、細胞凋亡、神經營養因子方面的作用。下丘腦-垂體-腎上腺(HPA)是神經內分泌調節的樞紐,起到維持內環境穩定的重要作用,如果持續亢進會引起機體系統功能的紊亂。模型大鼠的HPA 功能亢進,對外界的興奮性、反應性及快感也明顯減退,給藥YZ-50 后,促腎上腺皮質激素釋放激素(CRH)、促腎上腺皮質激素(ACTH)、血清皮質醇(COR)水平降低,說明遠志寡糖酯可以抵抗HPA 的亢進狀態,同時大鼠行為學癥狀也得到了改善[88]。YZ-50還可以促進模型大鼠海馬CA3 區細胞凋亡相關因子B 淋巴細胞瘤-2(Bcl-2)的表達、抑制Bcl-2 相關X的蛋白質(Bax)蛋白的表達,調控Bcl-2/Bax 比例而抑制神經細胞的凋亡,保護神經細胞[89]。酪氨酸蛋白激酶B(TrkB)是神經營養因子BDNF 的一種高親和力受體。實驗結果表明,YZ-50 可以提高造模大鼠海馬各區的BDNF 和TrkB 信使核糖核酸(mRNA)的表達。結合前期實驗,推測遠志寡糖酯通過提高神經營養因子活性促進慢性應激、HPA 軸亢進等所造成損傷神經元的恢復,從而改善應激模型大鼠的MDD 癥狀,有效發揮抗抑郁作用[90]。此外,近期研究結果表明,遠志醇提物還可以調節抑郁大鼠腸道內的菌群失調[91]。
巴戟天寡糖膠囊是推薦使用的抗抑郁中成藥之一[19]。一項臨床研究結果顯示,巴戟天寡糖膠囊治療輕中度MDD(腎陽虛證)患者8 周,可顯著降低漢密爾頓抑郁量表(HAMD)評分、腎陽虛證候評分、漢密爾頓焦慮量表(HAMA)評分,改善MDD狀及腎陽虛證候,并且有效率與鹽酸帕羅西汀陽性對照組的差異無統計學意義,療效不亞于鹽酸帕羅西汀[92]。對巴戟天寡糖膠囊治療MDD 的臨床隨機對照試驗進行Meta 分析,發現其對于MDD 患者臨床癥狀的改善作用與氟西汀相似[93]。
近年來,研究發現巴戟天寡糖膠囊與西醫常用抗抑郁藥聯合使用療效較好,并試圖從中發現巴戟天寡糖膠囊治療MDD 的機制。曹重陽等[94]發現,巴戟天寡糖膠囊與帕羅西汀聯合用藥8 周,治療效果優于單獨使用帕羅西汀。一項臨床研究顯示,巴戟天寡糖膠囊聯合阿戈美拉汀,可以顯著降低HAMD、HAMA 和中醫證候量化評分,緩解MDD癥狀,檢測發現患者血清中神經元特異性烯醇化酶(NSE)和髓鞘堿性蛋白(MBP)水平顯著降低,與單獨使用阿戈美拉汀組相比,巴戟天寡糖膠囊增加了阿戈美拉汀應用于老年MDD 的治療效果[95]。另一項臨床研究顯示,巴戟天寡糖膠囊還可以增加文法拉辛治療MDD的效果[96]。
4.7 其他
中藥寡糖主要藥理活性見表2,除此之外,其還有抗疲勞、治療心力衰竭等作用。郭敏等[97]發現BFO 具有明顯的抗疲勞作用,其能夠提高機體對運動負荷的適應能力,提高運動耐力,快速消除代謝廢物。還有研究發現,地黃寡糖可以通過影響干細胞移植來促進受損心肌細胞修復和再生,以治療心力衰竭[98-99]。小型豬脂肪組織間充質干細胞ADMSCs可在5-氮雜胞苷(5-aza)誘導下分化為心肌樣細胞,而地黃寡糖可促進第3 代ADMSCs 及5-aza 誘導后ADMSCs 的增殖[98]。此外地黃寡糖可減輕H2O2誘導的人脂肪源性間充質干細胞凋亡[99]。
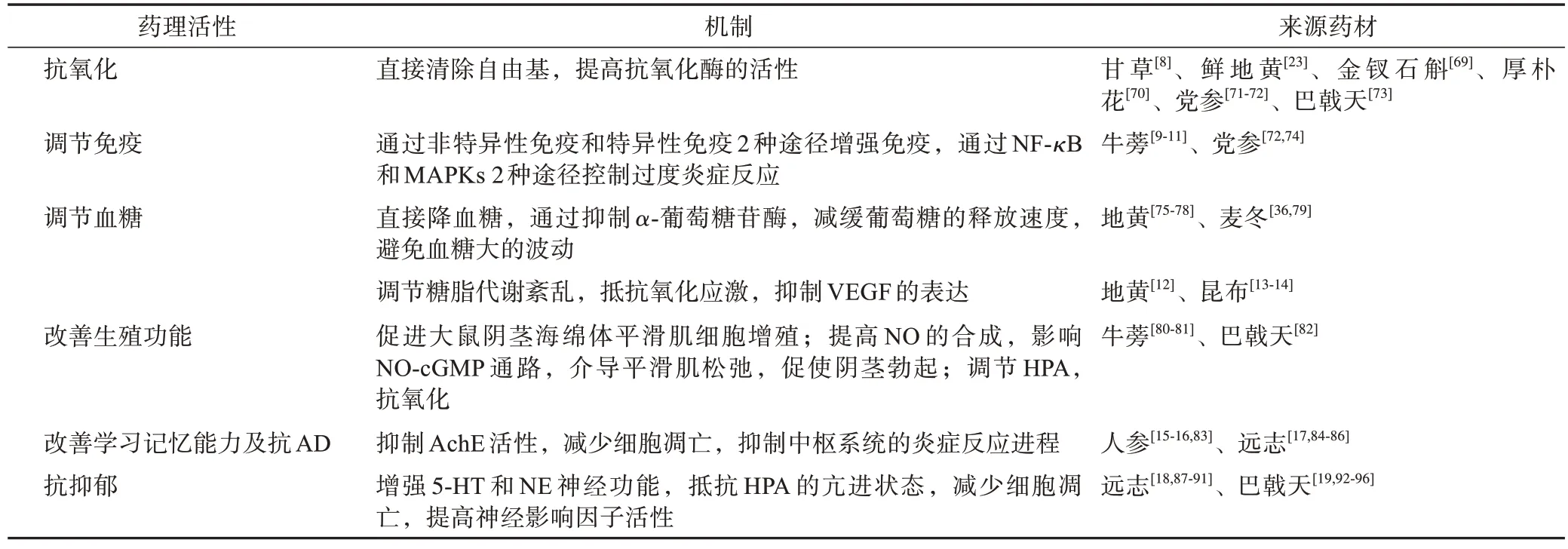
表2 中藥寡糖主要藥理活性
5 問題與展望
寡糖的提取方法有直接提取法、多糖降解法、合成法。前2 種方法較常使用,合成法由于技術手段的限制進展很慢。在研究多糖的低聚糖片段活性時,研究者常用TFA 降解的方法得到片段。相較于其他酸,TFA 酸性強,容易揮去,避免干擾后續實驗。相較于酶法,其對環境條件的要求低,成本也相對較低。但是,由于多糖的復雜結構,其聚合度、單糖組成、連接方式等即使是在同一批藥材中也會存在很大差異,加上無法控制水解位點,實驗中很難得到固定結構片段。筆者也發現目前關于多糖降解片段的研究多聚焦于聚合度范圍較窄的低聚糖混合物,而非單一寡糖。
在一些中藥中,糖類成分是主要活性成分。研究者利用HPLC,以結構相對簡單的寡糖的含量為指標,指導種植、采收、炮制、儲存過程,并試圖區分藥材的種質資源。寡糖含量的變化為中藥質量控制提供了新思路。也有人整合糖組學與代謝組學,對糖類組分和非糖類小分子組分同時進行系統比較分析,從整體角度研究不同炮制過程所得飲片的藥效物質基礎[100]。因此中藥糖類分子在質量控制方面具有較大的潛力。
目前的分離方法雖已經改善了傳統分離方法的不足之處,但仍存在一些問題,在分離效率上仍然需要提高,因此還需繼續探索有益、高效的分離純化方法。
中藥寡糖在免疫系統、內分泌系統、神經系統等方面都有著不可忽視的藥理價值,為尋找天然且不良反應低的替代藥提供了豐厚的資源基礎。在MDD 的治療中,巴戟天寡糖膠囊與帕羅西汀或文法拉辛聯合用藥的臨床研究結果提示中西醫結合可以協同增效。化學藥往往作用機制明確、針對性強,但是治療周期長、費用較高、不良反應多;中醫藥以辨證論治、整體調節為特點,作用溫和,不良反應少,但是診療標準不一。因此,中西醫結合可取長補短,實現優勢互補,達到協同增效的目的。
前期提取分離方法的優化為中藥寡糖的含量測定和活性研究提供了便利,使中藥寡糖在醫藥等領域發揮更加重要的作用。
