摘葉和噴施外源過氧化氫對庫爾勒香梨粗皮果形成的影響
2023-05-30 13:17:56史智勇艾沙江買買提仙米斯婭塔依甫曹藝潔玉蘇甫阿不力提甫
新疆農業科學 2023年4期
史智勇,艾沙江·買買提,仙米斯婭·塔依甫,曹藝潔,玉蘇甫·阿不力提甫
(1.新疆農業大學林學與園藝學院,烏魯木齊 830052;2.新疆農業科學院園藝作物研究所,烏魯木齊 830091;3.新疆維吾爾自治區農藥檢定所,烏魯木齊 830049 )
0 引 言
【研究意義】庫爾勒香梨(PyrussinkiangensisYü)是新疆梨和西洋梨的自然雜交后代,食用品質優良[1]。庫爾勒香梨粗皮果的形成降低了其食用品質和經濟效益。研究庫爾勒香梨粗皮果形成的規律,為庫爾勒香梨粗皮病的防治提供相關理論基礎,對提高果實品質具有重要意義。【前人研究進展】目前有關庫爾勒香梨粗皮果形成的研究主要集中在砧木、礦質元素、生長調節劑、樹體營養狀況、結果部位等的影響[2-4];環境因子對于香梨粗皮果的形成也有較大的影響,如干旱[5]、光照條件[6]等。庫爾勒香梨粗皮果的形成不僅是樹體的內部作用,也是樹體對外界環境信號響應,是復雜的信號傳遞過程。逆境脅迫是非生物脅迫的一種,也是植物在正常生長中經常會面臨的一種脅迫類型。李英麗[7]發現高溫處理能夠顯著增加鴨梨果實內活性氧含量的積累,且高溫和強光具有脅迫增效作用,能夠加重氧化脅迫的發生。石細胞含量的增多是庫爾勒香梨粗皮果形成的主要原因,而在石細胞快速形成的時期,其質外體活性氧產生基因也在高水平表達[8]。且活性氧對于石細胞的主要成分木質素[9]的合成有一定的促進作用[10-11],活性氧脅迫和高溫強光脅迫會促使香梨的粗皮果的產生。【本研究切入點】庫爾勒香梨粗皮病是一種生理病害。發病果實的果面凸凹不平,并且萼端由綠變黃的速度減慢,對果實品質影響較大。需研究庫爾勒香梨上可能誘導粗皮果形成的脅迫因子。【擬解決的關鍵問題】設置不同的脅迫類型,測定果實染色切片的相關生理指標,研究脅迫作用對庫爾勒香梨粗皮果形成的影響,分析庫爾勒香梨粗皮果形成相關的脅迫類型,為解決庫爾勒香梨粗皮果的產生提供理論基礎。
1 材料與方法
1.1 材 料
以新疆巴音郭楞蒙古自治州輪臺縣輪臺果樹資源圃的長勢相同的多年生庫爾勒香梨樹為試驗材料,隨機選取每棵樹上相同方向的枝條為1個試驗組,3棵樹為1個平行組,共9棵樹。
1.2 方法
1.2.1 試驗設計
空白(CK)組:不處理。
摘葉(Y)組:在2021年7月5日,摘除遮擋結果枝光照50%的葉子(高溫光照復合脅迫);外源噴施H2O2(H)組:在2021年5月5日,任意選取3個結果枝用250、400和500 mmol/L H2O2噴施預處理,觀察果子的生長發育情況。以果實表型不發生改變的最大臨界濃度作為試驗處理濃度(活性氧脅迫)。
400和500 mmol/L H2O2噴施組果實表型均產生明顯病變,而250 mmol/L H2O2噴施組表型未產生明顯病變,于2021年7月5日對整個H組的結果枝用250 mmol/L的外源H2O2噴施。圖1

圖1 外源噴施H2O2預試驗組
1.2.2 測定指標
1.2.2.1 環境因素
分別在CK組和Y組結果枝上各選取5個果子,選取7月連續晴朗的3 d,每隔2 h測定果面的光照度和溫度。
光照強度采用TES 1332A數位式照度計測定;采用FLUKE MT4 MAX輻射溫度計測定溫度。
1.2.2.2 生理指標
分別選取不同處理組枝條上的果實(花后100 d)若干,用蒸餾水清洗干凈后分別削取果皮以及果皮下2 mm左右的果肉,果皮果肉分別用液氮速凍后用 IKA A11 磨樣機磨成粉末狀混樣,做好采樣標簽,存于-80℃ 冰箱中備用。
采用硫酸蒽酮比色法測定植物纖維素含量[12];采用鹽酸水解法測定植物半纖維素含量,采用濃硫酸法測定植物木質素含量[13];可溶性蛋白采用考馬斯亮藍染色法測定;可溶性糖和淀粉含量采用蒽酮試劑法測定[14];丙二醛含量采用硫代巴比妥酸法測定[15];脯氨酸含量采用酸性茚三酮法測定[16];過氧化氫含量和超氧陰離子含量分別用索萊寶公司(Solarbio)的過氧化氫含量試劑盒(BC3590)和超氧陰離子含量試劑盒(BC1290)進行測定;酶活性測定利用賽默飛世爾(Thermo Fisher)酶標儀(1510)測定。
1.2.3 田間比例
在香梨果實成熟期,即2021年8月20日該對空白組以及2個處理組枝條上的所有果實的粗皮果和正常果數量并匯總。
1.2.4 果實石細胞染色
隨機選取不同處理組的香梨果實(花后100 d)若干,用蒸餾水清洗后分別對果實進行橫切與縱切。橫切與縱切的果實切片寬度約2 cm,厚度約5 mm,長度需要從果皮切到果核附近,用50% FAA固定液進行固定保存;香梨果實用番紅固綠染色液進行染色。
香梨果實的脫水和浸蠟通過徠卡組織樣品自動脫水機(Leica ASP200S)完成;香梨果實的包埋通過徠卡包埋機(Leica Arcadia H+C)完成;香梨果實的切片通過徠卡切片機(Leica RM2265)完成;香梨果實切片的染色通過徠卡攤片機(Leica HI 1210)、徠卡烘片機(Leica HI 1220)、徠卡自動染色機(Leica ST5020)完成。
1.3 數據處理
所有數據采用 Excel 2010 軟件進行匯總和處理,利用SPSS 26.0進行數據分析,利用 GraphPad Prism 7.0 進行作圖。
2 結果與分析
2.1 脅迫相關因子變化
2.1.1 摘葉組環境因子
研究表明,摘葉導致果實表面的光照強度以及溫度顯著增加。Y組果實表面的光照度明顯大于CK組,Y組的日平均光照度約為CK組的日平均光照度的18倍;Y組日平均溫度比CK組的日平均溫度高3.6℃。摘葉能使果實表面的光照和溫度明顯增加,對果實造成溫度和光照的復合脅迫。圖2
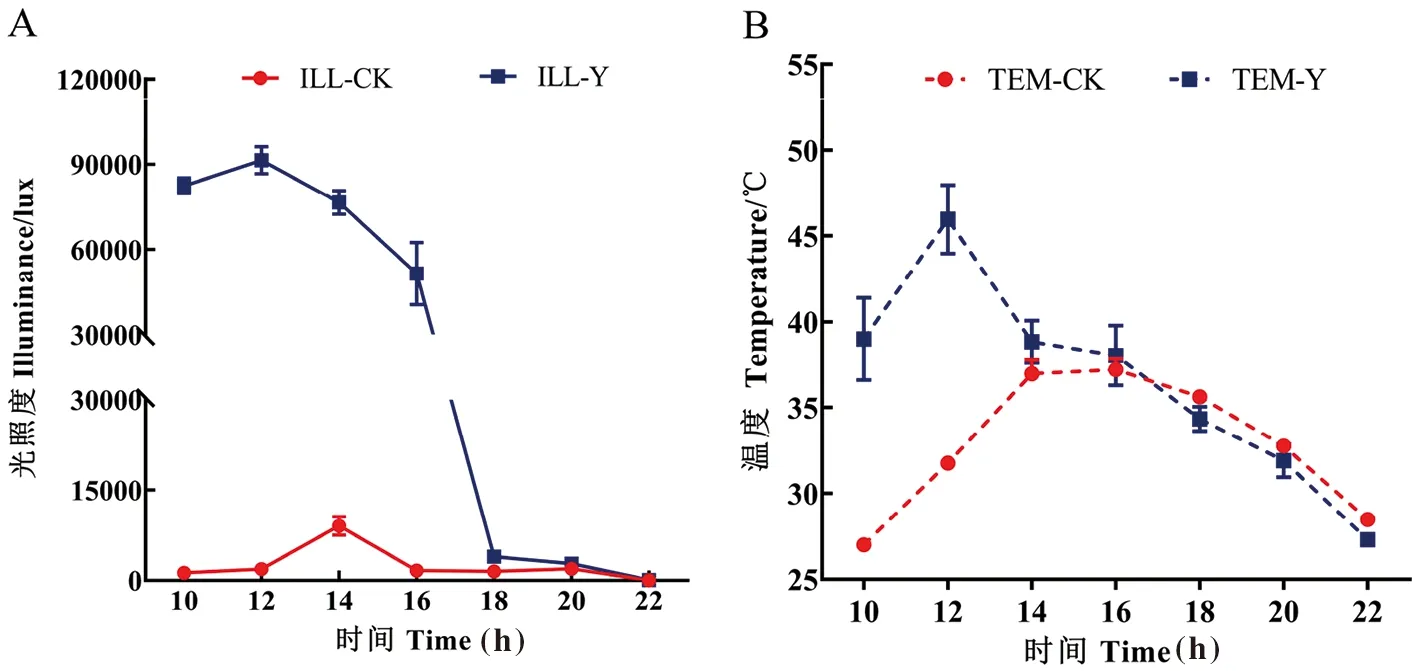
注:A圖為日光照變化;B圖為日溫度變化
2.1.2 不同脅迫因子對香梨果皮活性氧含量的影響
研究表明,噴施外源H2O2能導致庫爾勒香梨果皮中的H2O2含量明顯升高。H組H2O2含量約為CK組的2.0倍,并且也顯著高于其他2個處理組。外源噴施250 mmol/L H2O2能對庫爾勒香梨果皮造成一定的活性氧脅迫,而摘葉處理則沒有對庫爾勒香梨果皮造成明顯氧化脅迫或脅迫程度較小。圖3
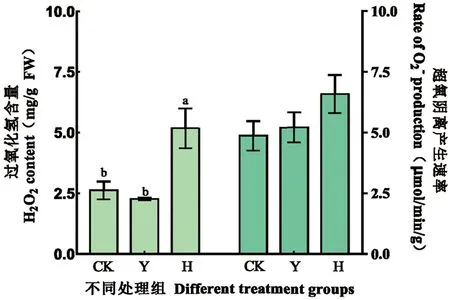
注:數據(平均值±標準差)后用不同小寫字母表示在P<0.05 差異顯著;無標注即代表沒有顯著性差異,下同
2.1.3 不同脅迫處理對庫爾勒香梨果實MDA含量的影響
研究表明,H組果肉中的MDA含量明顯高于CK組,為CK組的1.8倍。噴施250 mmol/L H2O2對果實的脅迫作用更加明顯,而摘葉處理對香梨果實MDA含量幾乎沒有影響。圖4
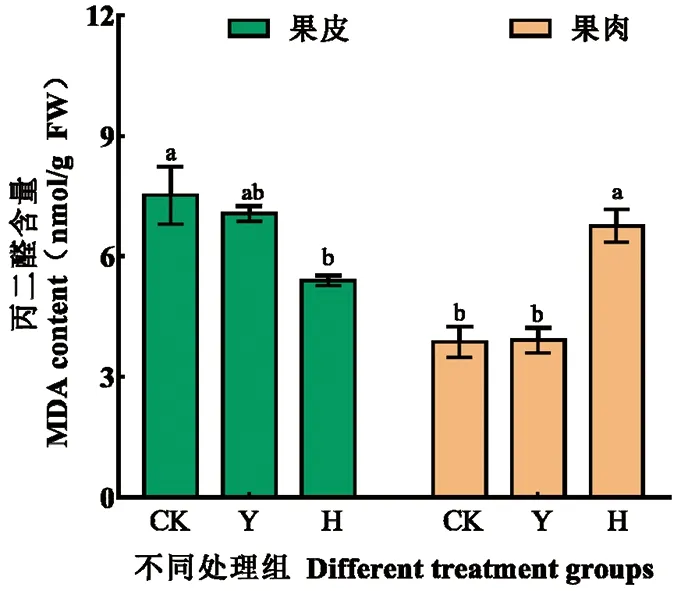
注:數據(平均值±標準差)后用不同小寫字母表示在P<0.05 差異顯著
2.2 不同脅迫因子對內源滲透物質的影響
研究表明,摘葉處理后,果皮和果肉中的可溶性糖含量和果肉中的淀粉含量均顯著性升高,分別為對照組的1.4、1.2倍和2.0倍;果肉中的可溶性蛋白顯著下降,相比對照組下降了49%。外源H2O2處理后,果皮中的可溶性糖和淀粉含量均顯著升高,分別為對照組的1.4倍和1.4倍;果皮中的脯氨酸含量顯著下降,相比對照組下降了14%。圖5

圖5 庫爾勒香梨果實不同處理組內源滲透物質含量變化
2.3 脅迫處理對庫爾勒香梨粗皮果形成的影響
2.3.1 脅迫處理對庫爾勒香梨粗皮果率的影響
研究表明,3種脅迫處理均能導致香梨的粗皮果率顯著升高。H組果實的粗皮果率為CK組的2.9倍,Y組果實的粗皮果率是CK組的1.6倍。且H組相對Y組均導致了其結果枝的粗皮果率的顯著升高。表1
2.3.2 脅迫處理對庫爾勒香梨組織染色切片的影響
研究表明,2種不同的脅迫處理后切片單位視野中石細胞的數量均體現出增加的趨勢。其中, H組染色圖中單位視野內的石細胞數量相對于Y組色圖中單位視野內石細胞數量增多趨勢更加明顯。與田間調查中的3個處理組對粗皮果率的結果相一致,活性氧脅迫對于果實內部石細胞含量的增多以及粗皮果的形成有較大的影響,高溫光照復合脅迫對于果實內部石細胞含量的增多以及粗皮果的形成影響較小亦或光照強度沒有達到對果實產生逆境脅迫的程度。圖6

表1 庫爾勒香梨結果枝不同處理組的粗皮果率變化
2.3.3 脅迫處理對庫爾勒香梨果實細胞壁物質含量的影響
研究表明,2種處理,果皮、果肉的木質素含量都有所增加。H組果皮和Y組果肉的木質素含量與CK組相比有顯著升高,H組果皮果肉和Y組果肉中的纖維素含量與CK組相比有顯著升高,H組果皮果肉和Y組果肉的半纖維素含量與CK組相比有顯著提高。H2O2噴施處理對果皮的木質素、纖維素、半纖維素含量以及果肉中的纖維素、半纖維素含量有顯著影響;摘葉處理對果肉中木質素含量、纖維素含量、半纖維素含量以及果皮中的半纖維素含量有顯著影響。圖7

注:A~C. CK,H,Y組的果實果皮附近染色圖,1、2、3分別為外果皮部位同一切片上的全景掃描圖的3個連續片段3段不同部位;a~c.處理組CK,H,Y組的果實果肉部分染色圖

注:A、B、C分別為果皮、果肉中木質素含量、纖維素含量、半纖維素含量;數據(平均值±標準差)后用不同小寫字母表示在P<0.05 差異顯著
3 討 論
庫爾勒香梨的粗皮果的形成原因較為復雜。徐慶岫等[5]認為7月的高溫干旱是導致粗皮果形成的主要原因;李疆等[3]通過對庫爾勒香梨進行田間調查,發現內膛枝、樹勢旺盛以及凸頂果和凹頂果的比率升高都會導致粗皮果率升高;此外,也有報道噴施鈣溶液可以影響南果梨粗皮果率[17]。香梨粗皮果的形成,不僅是樹體內部自身作用的結果,更是樹體對逆境環境的響應的結果。研究發現摘葉處理和噴施H2O2后果實果皮中的活性氧含量顯著增加,處理對香梨果實產生一定程度的氧化脅迫,其中H2O2處理的作用更明顯,所有影響細胞氧化還原狀態的脅迫因子都有潛力誘導粗皮果的產生。逆境脅迫會導致植物體內活性氧含量的升高[18],植物細胞在自身承受范圍之內通過保護酶系統或滲透調節物質來清除多余的活性氧來防御逆境脅迫[19-20]。脅迫處理后香梨果實內可溶性糖、可溶性蛋白、脯氨酸等滲透調節物質不同程度的提高,果實細胞確實感受到逆境脅迫,并對其做出響應。當逆境脅迫程度超過細胞的承受范圍后,植物組織為了整張發育主動犧牲部分細胞來適應逆境環境,即通過細胞程序性死亡(Programmed cell death,PCD)來促進部分細胞的分化[21]。其結果一是為剩余細胞的正常代謝活動提供能量;二是將部分細胞分化為厚壁組織或輸導組織的一部分來提高組織對逆境的適應性[22-23]。庫爾勒香梨粗皮果的形成是其果實中石細胞增多的結果,石細胞是由于木質素沉積而導致次生壁加厚的細胞分化過程[11,24]。
前期的研究中發現在石細胞形成的關鍵時期與質外體活性氧產生相關的酶基因顯著上調表達,且表達趨勢與石細胞的變化趨勢大致相同,石細胞分化和質外體活性氧積累之間存在協同關系。在果實發育的早期石細胞主要分布在維管束周圍,而維管束周圍的細胞一般處于一種“半分化”的狀態,后期不利的環境因素會優先導致維管束周圍的薄壁細胞進行分化[8,25]。庫爾勒香梨果實在受到一定的環境脅迫條件下,如外源的活性氧脅迫和高溫光照復合脅迫,會導致庫爾勒香梨的石細胞增多,相對應的處理組枝條的粗皮果率也會顯著增加。逆境脅迫、活性氧爆發、PCD、次生壁沉積這四者相結合,共同促進了庫爾勒香梨果實中石細胞的增多,進而導致粗皮果的產生。
4 結 論
4.12種處理均能顯著提高庫爾勒香梨果實內的木質素含量和處理枝條上的粗皮果率。2種處理后果皮的木質素含量以及粗皮果率分別為對照組的1.4、2.9倍和1.1、1.6倍。
4.2其中摘葉處理后,果肉中的滲透調節物質相對變化較為明顯,主要影響果肉中的石細胞積累;噴施H2O2后果皮中的ROS代謝相關物質、MDA含量以及滲透調節物質變化都較為明顯,主要影響果皮中的石細胞積累。
