巖黃連種子萌發特性研究
2023-05-30 00:28:13潘燕林郭倫發王新桂張怡彬秦洪波
廣西植物 2023年3期
潘燕林 郭倫發 王新桂 張怡彬 秦洪波

摘 要:? 巖黃連是一種多年生草本植物,主要分布于我國西南部巖溶地區,具有重要的生態及醫藥價值,開發利用前景廣闊。由于生存環境脆弱和人為采挖的影響使得野生巖黃連資源面臨枯竭,因此被列入2021年國家重點保護野生植物名錄。為探索該物種瀕危的原因,該文研究了種子保存方法、化學試劑前處理、溫度、光照、干旱、pH以及混沙保濕冷藏處理對種子萌發的影響。結果表明:(1)低溫保存能夠延長巖黃連種子的活性,保存2 a的種子萌發率仍能達到30%。(2)化學試劑HCl及NaClO對種子的前處理使其發芽率提升50%~60%;種子在20 ℃時萌發率能達到50%,而30 ℃時基本不能萌發;黑暗條件下的萌發率顯著高于周期性光照條件;萌發率隨著干旱程度加深不斷下降;萌發率在pH值3.0~8.0的條件下無顯著變化。(3)混沙保濕冷藏使種子發芽勢和萌發率顯著提高,萌發率達到對照的2倍。綜上認為,巖黃連種子在室溫條件下易失活且不能在30 ℃以上高溫萌發的特性與其瀕危有較大關系;高效的種子萌發方法可以為巖黃連保護與產業化應用提供有效途徑。該研究結果為野生巖黃連的保育提供了理論和技術保障,為其大田栽培和產業化推廣提供了技術支持。
關鍵詞: 巖黃連, 種子貯藏, 萌發方法, 萌發力指標, 萌發特性
中圖分類號:? Q945
文獻標識碼:? A
文章編號:? 1000-3142(2023)03-0580-07
Seed germination characteristics of Corydalis saxicola
PAN Yanlin1,2, GUO Lunfa1,2*, WANG Xingui1, ZHANG Yibin1, QIN Hongbo1
( 1. Guangxi Institute of Botany, Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences, Guilin 541006, Guangxi, China;
2. Guangxi Key Laboratory of Plant Functional Phytochemicals and Sustainable Utilization, Guilin 541006, Guangxi, China )
Abstract:? Corydalis saxicola is a perennial herbaceous plant, which mainly distributed in the karst areas of southwest China. Due to its important ecological and medicinal value, Corydalis saxicola has great prospects for development and utilization. With fragile living environmental condition and anthropogenic harvesting pressure, the wild resource of Corydalis saxicola is on the verge of exhaustion, so it has been included in the list of national key protected wild plants in 2021. In order to explore the causes for the endangerment of Corydalis saxicola and provide reference for its conservation and seedling breeding, this paper investigated the influencing factors on seed germination that including seed preservation methods, chemical pre-treatments, temperature, light, drought, pH, mix with wet sand and refrigerate treatment. The results were as follows: (1) Low-temperature storage can prolonged the seed activity, germination rate of 2-years stored seed could still reached 30%. (2) Germination rates of seed pre-treatments by HCl and NaClO solutions can reached to 50%~60%; germination rate was 50% at 20 ℃, while closed to zero at 30 ℃; darkness was more favorable for seed germination than periodic light condition; the seed germination rate kept decreasing with deepening drought; the seed germination rate did not changed significantly under the conditions of pH 3.0~8.0. (3) Mix with wet sand and refrigerate treatment can significantly promoted the seed germination potential and germination rate, and the germination rate was twofold of the control. In summary, the seed inactivation at room temperature and germination inability at high temperatures above 30 ℃ are more related to its endangerment; efficient seed germination methods can provide an effective way for conservation and industrial application of Corydalis saxicola. The results provide a theoretical and technical guarantee for the conservation of wild Corydalis saxicola, and technically support is cultivation.
Key words: Corydalis saxicola, seed storage, germination method, index of germination ability, germination characteristic
種子是植物有性繁殖的基礎,種子萌發是植物生長和發育的第一步。種子萌發受到其自身因素和外部環境的共同作用:種子的形態構造、化學成分、發育、成熟、休眠等均能影響萌發進程(Bareke, 2018);另外,環境因素如溫度、水分、氧氣、光照、土壤酸堿度等也會對種子萌發產生重要的影響(Carrera-Castao et al., 2020);細菌、真菌、病毒等微生物則是種子萌發的生物影響因素(Xiao et al., 2020)。種子萌發和幼苗定居障礙是許多珍稀瀕危植物致瀕的重要原因(Nicole et al., 2011; 盛茂銀等,2011)。研究瀕危植物的種子萌發特性,探尋種子萌發的限制因素,可以為其致瀕機制研究及保護策略的制定提供依據,也可以為其開發利用奠定基礎。
巖黃連(Corydalis saxicola),又名石生黃堇,是罌粟科(Papaveraceae)紫堇屬(Corydalis)植物,主要分布于廣西、貴州、云南等地的巖溶地區(余姣嬌等,2018)。巖黃連全株可入藥,主要藥用成分為生物堿類,包括脫氫卡維丁、巴馬汀和小檗堿等(Xie et al., 2021),具有抗炎、抗病毒和抗腫瘤的藥理作用(Guo et al., 2022),臨床上已被開發成相應的注射液和片劑用于肝炎、肝硬化及肝癌等治療。由于自身繁育困難與人為采挖,巖黃連野外種群數量銳減,被列入2021年國家重點保護野生植物名錄。為了解決野生資源保護與生產應用之間的矛盾,開展巖黃連種苗繁育、野外回歸保育及人工種植等工作勢在必行。
巖黃連植株多為叢生,7~14 cm長的總狀花序以頂生或葉對生方式從枝條上長出,授粉后形成的蒴果單個含20~30粒種子(蔣水元等,2002),30 d左右種子即能成熟。巖黃連種子細小,每克種子數量為1 700~2 000粒。以上這些特性對于種子育苗和人工種植都具有一定優勢。蔣水元等(2002)研究表明,巖黃連種子發芽率較低,即采即播發芽率最高為40%~50%,一般在20%~30%之間;隨著存儲時間增加發芽率不斷降低,室溫存儲半年后發芽率基本喪失。蔣運生等(2006)對巖黃連種子的不同著生部位、貯藏方法、貯藏時間和播種基質等影響因素進行實驗,證實著生在果穗中部的種子發芽率最高、紙袋密封和低溫貯藏的種子發芽率最好、火土加肥泥的播種基質更有利于種子萌發。另外,酸堿溶液對種子的萌發前處理也能影響其萌發率(陳柯羽,2011)。以上是關于巖黃連種子萌發的一些研究結果,但種子萌發特性在巖黃連致瀕原因、野外回歸保育及生產應用方面的研究還有待進一步加強,可推廣應用的優良種子萌發方法還需深入探索。
該研究以巖黃連種子為材料,針對種子保存條件和種子萌發處理方法進行研究,擬探究以下問題:(1)巖黃連種子較適合的保存方法;(2)化學試劑前處理、溫度、光照、干旱及pH對種子萌發的影響;(3)可推廣的巖黃連種子高效萌發方法。基于以上的研究,探究巖黃連的致瀕原因,從而為野生巖黃連的保育提供理論和技術保障;為其大田栽培和產業化推廣提供技術支持。
1 材料與方法
1.1 實驗材料
供試材料為2020年6月及2021年6月從廣西壯族自治區河池市東蘭縣桂源藥材專業合作社的栽培基地采收的巖黃連種子,將脫殼后的種子用塑料袋密封后置于4 ℃冰箱中避光保存備用。巖黃連種子呈橢圓形,種皮黑亮、質地堅硬,種子顆粒飽滿、健康,千粒重為0.465~0.472 g,水分含量為17.2%~18.5%。
1.2 實驗方法
1.2.1 溫度對巖黃連種子活力及萌發的影響 將適量2020年采收的巖黃連種子(簡稱2020年種子)和2021年采收的巖黃連種子(簡稱2021年種子)分裝到密封袋中,分別置于室溫和4 ℃冰箱中避光保存。分別取保存1、3、6個月后的種子,用4% NaClO溶液浸泡10 min除菌,無菌水洗6次后進行播種。每個培養皿播100粒左右的種子,每個處理3皿,每隔2~3 d加入1 mL無菌水保持種子濕潤,于25 ℃培養箱的暗培養條件下進行萌發。從萌發實驗的第10天開始,每隔2 d統計種子萌發數,若連續10 d無種子萌發則實驗結束。
另外,取4 ℃冰箱中保存的2021年種子,經 4% NaClO溶液浸泡及無菌水清洗后播種于培養皿中,分別置于20、25、30、35 ℃恒溫培養箱中,黑暗條件下進行萌發實驗。
1.2.2 化學試劑前處理對種子萌發的影響 取4 ℃冰箱中保存的種子,進行以下4種處理:①種子用清水漂洗后,直接用無菌水洗6次進行播種;②種子用4% NaClO溶液浸泡10 min,無菌水漂洗6次后播種;③種子用1 mol·L-1 HCl溶液浸泡1 h,無菌水漂洗6次后播種;④種子首先經1 mol·L-1 HCl溶液浸泡1 h,然后經4% NaClO溶液浸泡10 min,最后用無菌水漂洗6次后播種。在25 ℃黑暗條件下進行萌發實驗。
1.2.3 光照、干旱及pH對種子萌發的影響 取4 ℃冰箱保存的2021年種子,用1.2.2的4種處理方式處理種子后分別在25 ℃周期性光照(12 h光照/12 h暗)和完全黑暗條件下進行萌發實驗。
以PEG6000建立干旱脅迫,研究不同干旱脅迫下種子的萌發情況。取4 ℃冰箱保存的2021年種子,經4% NaClO溶液浸泡10 min后用無菌水清洗6次,分別置于去離子水對照和5%、10%、15%、20%的PEG6000溶液浸潤濾紙的培養皿中,每隔2~3 d向培養皿中加入1 mL的無菌水,保持PEG6000的相應濃度,于25 ℃黑暗條件下進行萌發實驗。
分別用HCl、NaOH溶液調節無菌去離子水pH值至3.0、4.0、5.0、6.0、7.0、8.0,取4 ℃冰箱保存的2021年種子,經4% NaClO溶液浸泡10 min后用無菌水清洗6次,置于不同pH值去離子水浸潤濾紙的培養皿中,每隔2~3 d向培養皿中加入1 mL相應pH值的無菌水,于25 ℃黑暗條件下進行萌發實驗。
1.2.4 混沙保濕冷藏對種子萌發的影響 將4 ℃冰箱保存的2020、2021年種子與干凈的河沙、無菌水按1∶10∶3的體積比例混合,置于密封袋中,并于4 ℃冰箱中避光保藏60 d,之后取出種子于室溫條件下進行萌發。同時,以干燥冷藏的種子作為對照,將其直接與河沙、無菌水混合后于室溫條件下進行萌發。
1.3 數據分析
萌發率=全部萌發種子數/播種種子數;發芽勢為萌發處理15 d后的種子萌發率。運用Excel軟件對數據進行統計和均值、方差分析;利用GraphPad Prism 5軟件進行顯著性分析和繪圖。
2 結果與分析
2.1 溫度對巖黃連種子活力及萌發率的影響
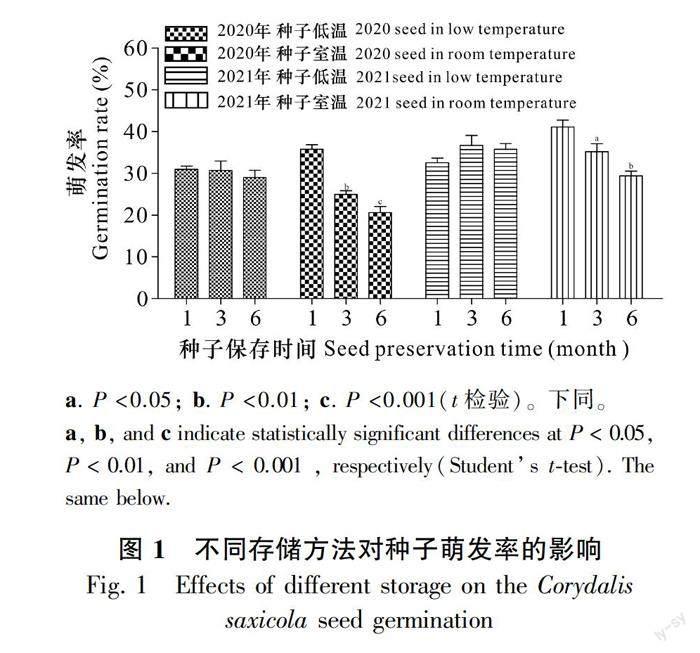
取保存于4 ℃冰箱的2020、2021年種子,于2021年11月開始分別進行室溫和4 ℃冰箱存儲,分別在存儲1、3、6個月后對種子進行萌發實驗。圖1結果顯示,4 ℃冰箱存儲的2020年種子發芽率基本沒有降低,保持在30%左右;4 ℃冰箱存儲的2021年種子的發芽率不僅沒有降低,還稍微有所提升;2020年種子經室溫保存后,發芽率隨保存時間增加不斷降低;2021年種子與2020年種子相似,發芽率隨室溫保存時間增加也逐漸降低。這說明相比于室溫,4 ℃冰箱保存更有利于延長種子活力,4 ℃冰箱保存2 a的種子萌發率仍能達到30%。
以2021年種子為材料,分別在20、25、30、35 ℃的恒溫培養箱中進行萌發實驗。圖2結果顯示,20 ℃條件下的種子發芽率最高,25 ℃發芽率與20 ℃相比雖稍微有所降低,但不顯著;在30 ℃時,種子基本不能萌發,35 ℃時種子完全不能萌發,與20 ℃條件相比均呈顯著降低。這說明高溫明顯不利于種子的萌發。
以上結果說明,溫度是影響種子活力及萌發率至關重要的因素之一。與室溫相比,低溫保存更有利于延長種子的活力;種子在20~25 ℃條件下的發芽勢及發芽率都能保持在較高水平,而在30 ℃以上的高溫下,種子幾乎完全失活,從而喪失萌發率。
2.2 化學試劑處理對巖黃連種子萌發的影響
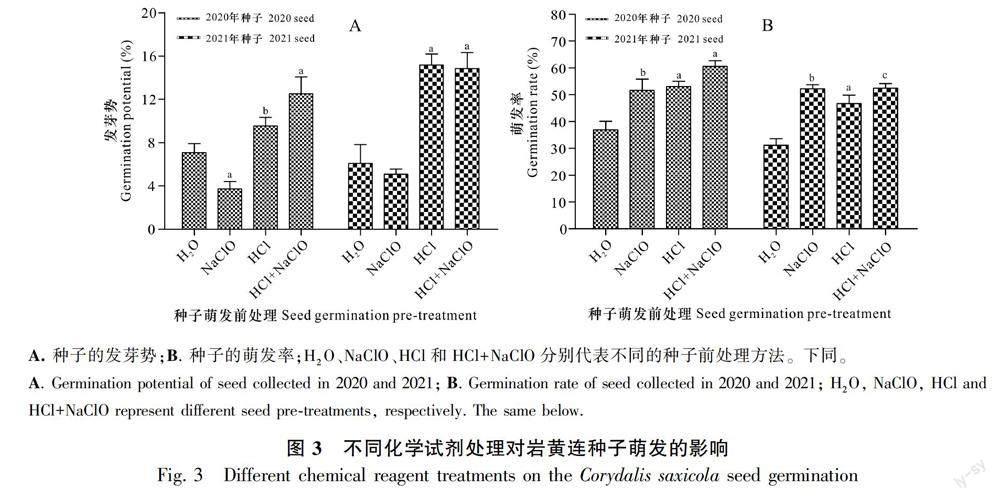
圖3結果顯示,與H2O對照相比,4% NaClO溶液處理種子對發芽勢無促進作用,甚至會使發芽勢稍微降低,而1 mol·L-1 HCl、HCl+NaClO處理均能顯著提高種子發芽勢(圖3:A)。無論是1M HCl、4% NaClO單獨處理還是兩種試劑合并處理,均能顯著提高種子發芽率(圖3:B)。除利用HCl和NaClO溶液處理種子外,還嘗試使用NaOH溶液處理種子,處理后基本未能觀察到種子萌發。這說明酸性化學試劑對種子的前處理能顯著提高其發芽率,其中HCl、HCl+NaClO溶液的處理能同時顯著提高發芽勢和發芽率,是較優的種子前處理方法。
2.3 光照和化學試劑處理對巖黃連種子萌發的影響
圖4結果顯示,以2021年種子為材料,經4種前處理的種子在黑暗條件下的發芽率都顯著高于光照條件,并且經NaClO+HCl處理后,暗培養條件下種子發芽率的增高率比直接水浸更為顯著。以上結果說明,黑暗條件更有利于種子的萌發。
2.4 干旱對巖黃連種子萌發的影響
以2021年種子為材料,以H2O為對照,以5%、10%、15%、20%PEG6000溶液(分別代表不同干旱程度)為不同干旱條件,于25 ℃黑暗條件下進行萌發實驗。圖5結果顯示,種子在H2O條件下發芽勢和萌發率最高;在5% PEG6000溶液處理下發芽勢降低,萌發率與對照無明顯差異;10%、15% PEG6000溶液處理下的種子發芽勢和萌發率與對照相比均顯著降低;在20% PEG6000處理下,種子萌發明顯滯后且萌發率顯著低于對照,為對照組的50%左右。這說明干旱脅迫對種子的萌發具有一定影響,隨著干旱程度增加,雖然發芽勢逐漸降低,但只有達到較高程度的干旱時,其種子萌發率才會顯著降低。
2.5 不同pH條件對巖黃連種子萌發的影響
以2021年種子為材料,分別置于pH值為3.0、4.0、5.0、6.0、7.0、8.0的無菌去離子水潤濕培養皿中,于25 ℃黑暗條件下進行萌發實驗。圖6結果顯示,6個不同pH條件下的種子萌發率均在40%左右浮動,發芽勢差異不大。這說明種子在pH值3.0~8.0不同條件下萌發,發芽勢和萌發率均無顯著差異。
2.6 混沙保濕冷藏對巖黃連種子萌發的影響
以2020、2021年種子為材料,采用混沙保濕冷藏60 d后室溫萌發和直接室溫萌發2種方式,研究巖黃連種子適宜的發芽方式。圖7結果顯示,由于冬季室溫偏低,室溫條件下種子需要在38 d后開始萌發,在76 d后萌發基本結束;而經混沙保濕冷藏處理的種子,置于室溫條件下僅需4 d即可萌發且快速達到萌發高峰。從發芽率結果(圖7)來看,經過低溫混沙濕藏的種子萌發率顯著高于直接室溫萌發的發芽率。這說明混沙保濕冷藏處理能顯著提高種子的發芽勢和萌發率,有利于提升苗的整齊度。
3 討論與結論
3.1 環境因子對巖黃連種子萌發的影響
種子生存能力受其自身特性和儲藏條件的共同影響,低溫干燥保存有利于降低種子變質、減緩種子呼吸和其他代謝進程,是一種常用的種子保存方法(Bareke,2018)。蔣水元等(2002)研究證實,巖黃連種子壽命很短,常溫保存幾個月后就會喪失大部分活力。本研究結果顯示,低溫密封保存種子的萌發率顯著高于常溫密封保存,常溫保存6個月的種子萌發率為低溫保存的2/3左右,說明巖黃連種子較適宜低溫干燥的保存方式。本研究結果常溫保存種子萌發率與蔣水元等(2002)報道的結果有所差異,推測保存室溫相對偏低(本研究保存時間為11月至次年5月)和密封保持種子水分含量等方式能夠有效延緩巖黃連種子活力降低的進程。
適宜的溫度有助于種子體內酶的活化,促使赤霉素和脫落酸等影響種子萌發的激素水平發生改變,從而促進種子萌發(Carrera-Castao et al., 2020)。本研究的溫度梯度實驗證實,20~25 ℃對種子的萌發較為有利,30~35 ℃時種子發芽率基本喪失, 說明巖黃連種子萌發對溫度的耐受范圍較窄。巖黃連植株生長期最適宜的溫度為10~20 ℃,高于25 ℃后,各器官的衰老加劇。這說明巖黃連從種子萌發到生長發育階段都偏好于涼爽的氣候,高溫等極端天氣可能是加劇該物種瀕危的重要因素。
種皮是種子與外界環境的屏障,具有保護種胚和防止病蟲害入侵的作用,但種皮也會對種子萌發造成障礙。韓鳳等(2012)研究證實,毛黃堇種子外被蠟質,影響了吸水膨脹和通氣,對萌發有一定的抑制作用。巖黃連種子與其近源種毛黃堇種子類似,其種皮對種子萌發可能存在抑制作用。適量的酸能腐蝕種皮、增強種子的透水透氣性、解除種子內外物質交換的障礙, 從而提高其發芽率和縮短萌發周期(Bareke,2018)。此外,酸還能降解種皮上的部分抑制性成分、殺死細菌或真菌以減少霉變,從而促進萌發(李兵兵等,2013)。本研究中, 利用HCl和NaClO對巖黃連種子進行萌發前處理,其發芽勢和萌發率都得到了顯著提升,說明適當的酸處理能夠解除巖黃連種皮對萌發的抑制作用,從而促進種子萌發。
水分和光照是種子萌發的關鍵影響因素。種子吸水能觸發呼吸活動的恢復、能量的產生、修復機制的激活、蛋白質的生物合成和運輸等生理生化反應(Carrera-Castao et al., 2020)。不同物種的種子萌發對光照的需求差異很大(Yang et al., 2020),光抑制種子萌發被認為是避免種子在土壤表面或接近土壤表面發芽,從而保護幼苗在極端高溫下免受脫水和暴露的一種生理適應性(Carta et al., 2017)。野生巖黃連植株常生長于無直射光且雨水不能直接淋濕的陡壁石縫中(陸瑞群,2006),具有避強光性和一定的干旱耐受性。本研究中,種子的發芽勢隨著干旱程度的增加而逐漸降低,低程度干旱脅迫下的發芽率與對照相比無顯著差異,只有在較高干旱脅迫時發芽率才會明顯降低,說明巖黃連種子萌發能耐受一定的干旱脅迫。光照和黑暗條件下的萌發實驗證實,巖黃連種子在黑暗條件下萌發率更高,說明強光對巖黃連種子萌發具有抑制作用。
種子萌發和幼苗生長階段對酸堿環境的適應性是決定植物能否生存的關鍵(申玉華等,2016)。本研究中,pH值為3.0~8.0的條件下,種子發芽勢及萌發率均無顯著差異,說明巖黃連種子萌發能耐受較寬范圍的酸堿度變化,這與其植株對黑色石灰土與酸性紅壤的較強適應能力具有一致性(蔣水元等,2002)。
陸瑞群(2006)研究結果證實,巖黃連種子的發芽過程慢且不整齊;本研究中,種子在恒溫條件下萌發需要1個月左右完成,室溫條件萌發則需要2個月左右。該種子萌發特性可能對巖黃連這類珍稀瀕危植物類群在惡劣生境中的持續存在具有重要意義,發芽過程緩慢有助于部分種子先避開高溫、干旱和強光等惡劣環境后再進行萌發、生長和繁衍。
3.2 巖黃連種苗繁育困難的原因分析及對其野外保育的啟示
巖黃連母株的種子產量較高,而其野外的幼苗更新卻非常困難,推測是由種子難萌發、幼苗難存活等因素共同造成的。本研究證實,巖黃連種子在溫度超過30 ℃時基本不能萌發,并且隨著干旱程度加深種子發芽勢和發芽率都會降低。自然環境下,巖黃連植株于3月下旬至4月進入盛花期,種子成熟后掉落萌發時間在6—8月之間(錢月華,2019)。6—8月的氣溫基本都會超過25 ℃,常在30~35 ℃之間波動,嚴重制約了巖黃連種子的萌發;另外,空氣濕度變化大、巖黃連所生長的巖縫環境缺少水分等因素也不利于巖黃連種子的萌發;巖黃連幼苗具有高度依賴外部環境和易感病害等特性,使得能存活長大的植株數量進一步減少(蔣水元等,2006)。以上因素共同導致了巖黃連野外幼苗更新困難,而人為采挖進一步威脅了種群的更新和擴大。
本研究中,低溫保存2 a的種子發芽率都能達到30%,有效延長了巖黃連種子的活力;化學試劑對種子的前處理能進一步將萌發率提高至50%左右。這證明巖黃連播種不必受限于即采即播,可以先在種子采集后進行低溫冷藏以保持其活力,再選擇適當的氣候進行播種;另外,還可以在播種前通過化學試劑處理來進一步提高其萌發率。在巖黃連野外回歸保育上,采取錯峰播種使其種子更易萌發,或許可以獲得更多的幼苗,從而提高其種群存活數量,實現其野外保育的目的。
3.3 可推廣應用的巖黃連種苗繁育方法
巖黃連除了用于藥品生產外,還用于巖溶區的植被恢復、林下種植、園林景觀等領域,應用前景廣闊。由于生產應用條件的限制,人們較難對巖黃連種子的消毒滅菌和萌發條件進行嚴格的控制。本研究中,與直接室溫萌發相比,對巖黃連種子先用混沙保濕冷藏一段時間后再進行室溫萌發的操作能夠顯著提高種子的發芽勢和萌發率,該方法操作簡單,所需設備僅為4 ℃冰箱,在實際生產上具有普遍推廣和應用的潛能。
參考文獻:
BAREKE T, 2018. Biology of seed development and germination physiology [J]. Adv Plants Agric Res, 8(4): 336-346.
CARRERA-CASTAO G, CALLEJA-CABRERA J, PERNAS M, et al., 2020. An updated overview on the regulation of seed germination [J]. Plants, 9(6): 703.
CARTA A, SKOURTI E, MATTANA E, et al., 2017. Photoinhibition of seed germination: occurrence, ecology and phylogeny [J]. Seed Sci Res, 27(2): 131-153.
CHEN KY, 2011. Molecular cloning and expression analysis of involved in berberine alkaloids biosynthesis in Corydalis saxicola Bunt. and microbial transformation of steroidal alkaloids in Veratrum nigrum Linn [D]. Shanghai: Second Military Medical University: 1-176.? [陳柯羽, 2011. 巖黃連中小檗堿型生物堿生物合成途徑相關基因的研究及藜蘆中甾體生物堿的微生物轉化 [D]. 上海: 第二軍醫大學: 1-176.]
GUO YR, ZHAO LJ, CHAO BT, et al., 2022. The traditional uses, phytochemistry, pharmacokinetics, pharmacology, toxicity, and applications of Corydalis saxicola Bunting: a review [J]. Front Pharmacol, 13: 822792.
HANG F, LING MX, XIAO JY, et al., 2012. Preliminary study on the seed germination characteristics of Corydalis tomentella [J]. Hunan Agric Sci, (4): 20-21.? [韓鳳, 林茂祥, 肖杰易, 等, 2012. 毛黃堇種子萌發特性的初步研究 [J]. 湖南農業科學, (4): 20-21.]
JIANG SY, HU XH, ZHAO RF, 2002. Study on the introduction and cultivation of Corydalis saxicola Bunting [J]. Guihaia, 22 (5): 469-473.? [蔣水元, 胡興華, 趙瑞峰, 2002. 巖黃連引種栽培研究 [J]. 廣西植物, 22(5): 469-473.]
JIANG SY, WEI X, LI H, et al., 2006. GAP standard operating procedure on Corydalis saxicola Bunting [J]. Guihaia, 26(6): 675-680.? [蔣水元, 韋霄, 李虹, 等, 2006. 巖黃連規范化種植標準操作規程(SOP) [J]. 廣西植物, 26(6): 675-680.]
JIANG YS, ZHU HJ, JIANG SY, et al., 2006. Seeding propagation of Corydalis saxicola Bunting [J]. Guangxi Sci, 13(4): 324-326.? [蔣運生, 朱鴻杰, 蔣水元, 等, 2006. 巖黃連種子繁殖研究 [J]. 廣西科學, 13(4): 324-326.]
LI BB, WEI XH, XU Y, 2013. The causes of Gentiana straminea Maxim. seeds dormancy and the methods for its breaking [J]. Acta Ecol Sin, 33(15): 4631-4638.? [李兵兵, 魏小紅, 徐嚴, 2013. 麻花秦艽種子休眠機理及其破除方法 [J]. 生態學報, 33(15): 4631-4638.]
LU RQ, 2006. The geologic background and the tissue culture technique research of Corydalis Saxicola Bunting [D]. Guilin: Guangxi Normal University: 1-45.? [陸瑞群, 2006. 巖黃連的地質背景及組織培養技術研究 [D]. 桂林: 廣西師范大學: 1-45.]
NICOLE F, DAHLGREN JP, VIVAT A, et al., 2011. Interdependent effects of habitat quality and climate on population growth of an endangered plant [J]. J Ecol, 99(5): 1211-1218.
QIAN YH, 2019. Cultivation techniques for rare medicinal plant Corydalis saxicola [J]. Anhui For Sci Technol, 45(2): 28-30. [錢月華, 2019. 珍稀藥用植物巖黃連的栽培技術 [J]. 安徽林業科技, 45(2): 28-30.]
SHENG MY, SHEN CZ, CHEN X, et al., 2011. Resources situation and conservation strategies of endangered wild plants in China [J]. Chin J Nat, 33(3): 149-154.? [盛茂銀, 沈初澤, 陳祥, 等, 2011. 中國瀕危野生植物的資源現狀與保護對策 [J]. 自然雜志, 33(3): 149-154.]
SHEN YH, WANG JH, KANG JH, et al., 2016. Study on seed germination characteristics of Medicago sativa L. under acid and alkali stress [J]. Seed, 35(9): 68-72.? [申玉華, 王嘉慧, 亢俊樺, 等, 2016. 酸堿脅迫下紫花苜蓿種子萌發期的抗逆性研究 [J]. 種子, 35(9): 68-72.]
XIAO ZX, ZOU T, LU SG, et al., 2020. Soil microorganisms interacting with residue-derived allelochemicals effects on seed germination [J]. Saud J Biol Sci, 27(4): 1057-1065.
XIE GY, JIN SY, LI HT, et al., 2021. Chemical constituents and antioxidative, anti-inflammatory and anti-proliferative activities of wild and cultivated Corydalis saxicola [J]. Ind Crop Prod, 169(537): 113647.
YANG LW, LIU SR, LIN RC, 2020. The role of light in regulating seed dormancy and germination [J]. J Integr Plant Biol, 62(9): 1310-1326.
YU JJ, QIU ZX, LIU QY, et al., 2018. Research advance on Corydalis saxicola Bunting [J]. J Pharm Res, 137(6): 342-345.? [余姣嬌, 邱志霞, 劉秋燕, 等, 2018. 巖黃連的研究進展 [J]. 藥學研究, 137(6): 342-345.]
(責任編輯 蔣巧媛)
收稿日期:? 2022-09-06
基金項目:? 中央引導地方科技發展資金項目(桂科ZY21195035); 桂林市重點研發計劃項目(20210211-1); 廣西科學院基本科研業務費項目(CQZ-C-1913)。
第一作者: 潘燕林(1986- ),博士,副研究員,主要從事特色經濟植物重要性狀及功能基因研究,(E-mail)panyanlinll@163.com。
通信作者:? 郭倫發,副研究員,主要從事特色經濟植物的研究與利用研究,(E-mail)37623630@qq.com。
