干旱脅迫對不同抗旱類型蕪菁苗期生理特性的影響
2023-05-27 01:06:50高亞寧軒正英張凱浩阿依買木沙吾提馬國財
干旱地區農業研究 2023年3期
高亞寧,軒正英,張凱浩,李 玉,常 嬌,阿依買木·沙吾提,馬國財
(1.塔里木大學園藝與林學學院,新疆 阿拉爾 843300;2.塔里木大學南疆特色果樹高效優質栽培與深加工技術 國家地方聯合工程實驗室,新疆 阿拉爾 843300;3.塔里木大學分析測試中心,新疆 阿拉爾 843300)
蕪菁(Brassicarapa.L)別名蔓菁、圓根、盤菜等,由油用亞種演化而來,隸屬于十字花科蕓薹屬[1]。其維吾爾語又稱為恰瑪古,是生長在新疆天山南部、塔里木盆地西北,具有藥用、食用、飼用三大應用價值的2 a生草本植物[2-3]。新疆屬于極旱地區,土地沙漠化嚴重、日照時間長、水分蒸發量大,大多數土地為山脈、戈壁和沙漠[4]。蕪菁雖耐瘠薄、耐干旱,有較強的適應性,但長時間的干旱少雨仍會在一定程度上對蕪菁的葉片及肉質根生長發育造成嚴重影響。眾多研究表明,干旱脅迫可引起植物的生理生化過程損害,如細胞膜結構[5]、關鍵酶活性[6]、活性氧合成與積累[7]、氣孔調節和其他氣體交換特性[8]。植物細胞可以調控基因的表達、產生新的蛋白質,進而引起植物生理和代謝上的變化來應對干旱脅迫[9]。例如降低凈光合速率,改變植物代謝途徑,累積可溶性物質,合成脯氨酸、甜菜堿等滲透調節物質,分解體內原有的蛋白質,同時合成參與各種代謝調節相關的酶[10]。Jabeen等[11]研究表明干旱脅迫導致蕪菁的生長量(根和莖的干質量與鮮質量、根長與莖長)、色素含量(葉綠素a,b和總葉綠素)、總酚和抗壞血酸含量減少,顯著增強了蕪菁葉片與根部的游離脯氨酸、甜菜堿、丙二醛、過氧化氫的含量以及提高了其過氧化氫酶和超氧化物歧化酶活性。
當前在蕪菁的營養品質、遺傳多樣性和非生物脅迫等多方面都開展了大量的研究[1-3,11],但鮮有關于蕪菁抗旱性方面的研究。本試驗選取前期篩選出的1個抗旱型資源和1個干旱敏感型資源,采用盆栽自然干旱法,研究不同抗旱類型蕪菁在苗期干旱脅迫下的生理生化變化,旨為深入研究蕪菁抗旱生理機制奠定基礎,為蕪菁抗旱品種選育及抗旱栽培提供理論依據。
1 材料與方法
1.1 試驗設計
供試材料為本課題組前期獲得的干旱敏感型資源WJC106和抗旱型資源WJC129,采取盆栽自然干旱的方法進行試驗。于2021年8月在塔里木大學園藝試驗站智能溫室中進行育苗。分別將種子播種于裝有125 g混合基質(草炭、蛭石、園土按2∶2∶1體積混合)的營養缽中(每個營養缽播3粒),每份試材共播90個營養缽。育苗期間采取正常管理,當幼苗長出第2片真葉時進行定苗,每盆保留1株苗。待幼苗長至四葉一心時,采用自然干旱的方法進行干旱處理。共設置對照組(CK)和干旱組(T)兩個處理,各3次重復,每個重復15株。對照組:土壤相對含水量保持在70%~80%之間;干旱組:干旱處理前澆一次透水,之后不再澆水讓其自然干旱。直到幼苗葉片出現嚴重萎蔫時(干旱第10 d)試驗結束。
1.2 測定指標與方法
干旱處理結束后采用組內隨機取樣的方法,選取植株第4片長勢一致的葉片測定其生理指標,每個重復5片,每個處理3次重復共15片葉。參考李合生[12]的方法測定其相對電導率、總葉綠素、可溶性糖、可溶性蛋白、丙二醛和游離脯氨酸含量。采用蘇州科銘生物技術有限公司生產的試劑盒測定超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性和過氧化氫酶(CAT)活性。
采用發射掃描電子顯微鏡(APREO-S,Thermo Fisher Scientific公司,美國)對葉片的氣孔特征進行觀察,并用Image Pro Plus軟件進行指標測量。每個葉片隨機選取10個視野,主要測量氣孔張開長度、氣孔張開寬度、氣孔長度、氣孔寬度、氣孔開度、保衛細胞厚度、氣孔密度。氣孔開度=氣孔張開長度×氣孔開張寬度×π/4,氣孔密度=視野下氣孔數/視野下的面積[13]。
1.3 數據處理
采用Microsoft excel 2019軟件進行數據整理,用SPSS 25.0及 Origin 9.0軟件對試驗數據進行分析及作圖。
2 結果與分析
2.1 干旱脅迫對不同抗旱類型蕪菁苗期丙二醛含量及相對電導率的影響
丙二醛作為膜脂過氧化的最終分解產物,可反映膜系統的受傷害程度和抗旱性強弱,同時對細胞產生毒害作用[13];相對電導率是衡量細胞膜透性的重要指標[14-15]。如圖1所示,兩個蕪菁資源在干旱脅迫下,其苗期葉片丙二醛含量和相對電導率相對于對照組均呈現出顯著上升趨勢。圖1A顯示,與對照組相比,WJC106在干旱脅迫下的丙二醛增量顯著高于WJC129,增幅分別為59.71%和54.52%。圖1B顯示,WJC106和WJC129在干旱脅迫下相對電導率值與對照組相比,分別增加了27.05%和20.06%。干旱脅迫下,干旱敏感型的WJC106蕪菁苗期細胞膜受損程度大于抗旱型的WJC129。
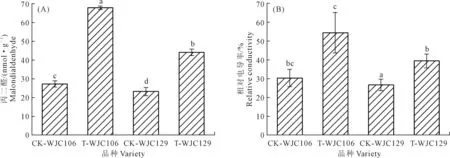
注:不同小寫字母表示處理間差異顯著(P<0.05),下同。Note:Different lowercase letters indicate significant differences between treatments at P<0.05,the same below.圖1 干旱脅迫對蕪菁苗期丙二醛含量及相對電導率的影響Fig.1 Effects of drought stress on MDA and relative electrical conductivity of turnips seedling
2.2 干旱脅迫對不同抗旱類型蕪菁苗期抗氧化酶活性的影響
如圖2所示,干旱脅迫使兩個不同抗旱類型蕪菁苗期的SOD、POD和CAT活性增加。圖2A、2B表明,與對照組相比,WJC129蕪菁幼苗葉片的SOD、POD活性受干旱脅迫的影響呈顯著增加趨勢,而WJC106葉片的SOD和POD活性與對照組無顯著差異。從圖2C可以看出,相比于對照組,兩個不同抗旱類型蕪菁在干旱脅迫下的CAT活性均顯著增加,且干旱下WJC129的CAT活性顯著大于WJC106。抗旱型的WJC129蕪菁苗期保護酶活性變化大于干旱敏感型的WJC106。
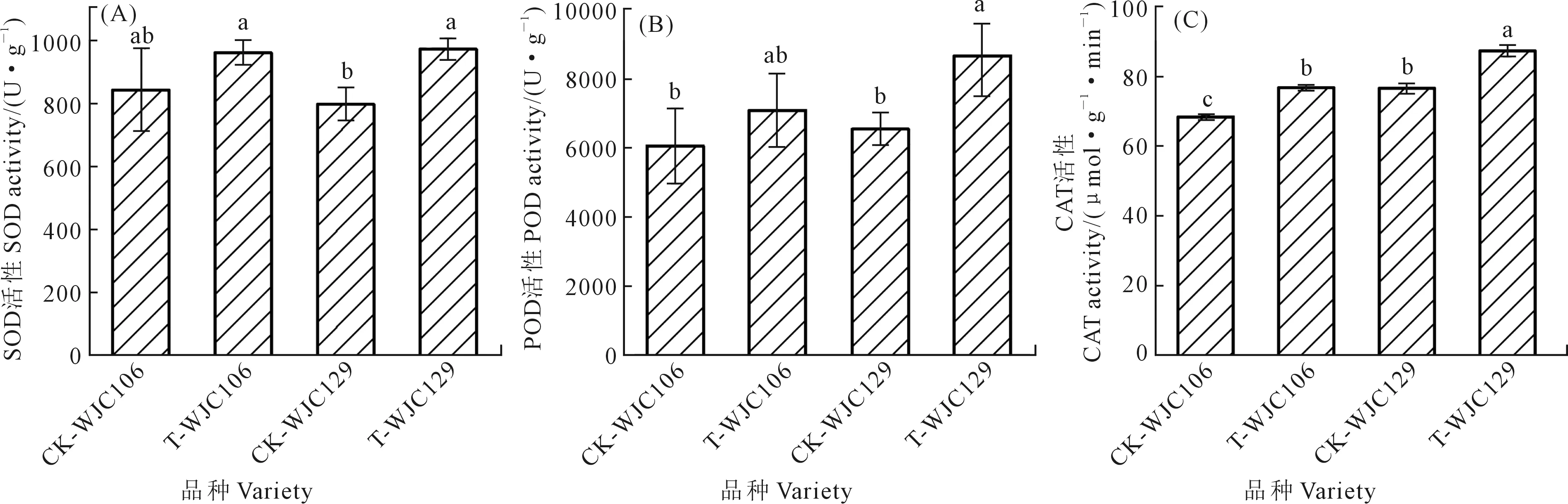
圖2 干旱脅迫對蕪菁苗期抗氧化酶活性的影響Fig.2 Effects of drought stress on antioxidant enzyme activity of turnips seedling
2.3 干旱脅迫對不同抗旱類型蕪菁苗期滲透調節的影響
圖3A、3B顯示,干旱脅迫下WJC106和WJC129兩個蕪菁的脯氨酸和可溶性蛋白含量均顯著高于對照組,其脯氨酸增量分別為85.22%和88.97%,可溶性蛋白增量為32.51%和58.26%。圖3C顯示,干旱條件下兩個不同抗旱類型蕪菁苗期葉片可溶性糖含量均增加,但無顯著差異。由此可知,干旱脅迫使蕪菁苗期滲透調節物質含量增加,且WJC129滲透調節物質變化量高于WJC106,抗旱型蕪菁苗期在受到干旱脅迫時的滲透調節能力強于干旱敏感型;同時脯氨酸和可溶性蛋白在干旱脅迫下較為敏感,起主要滲透調節作用。
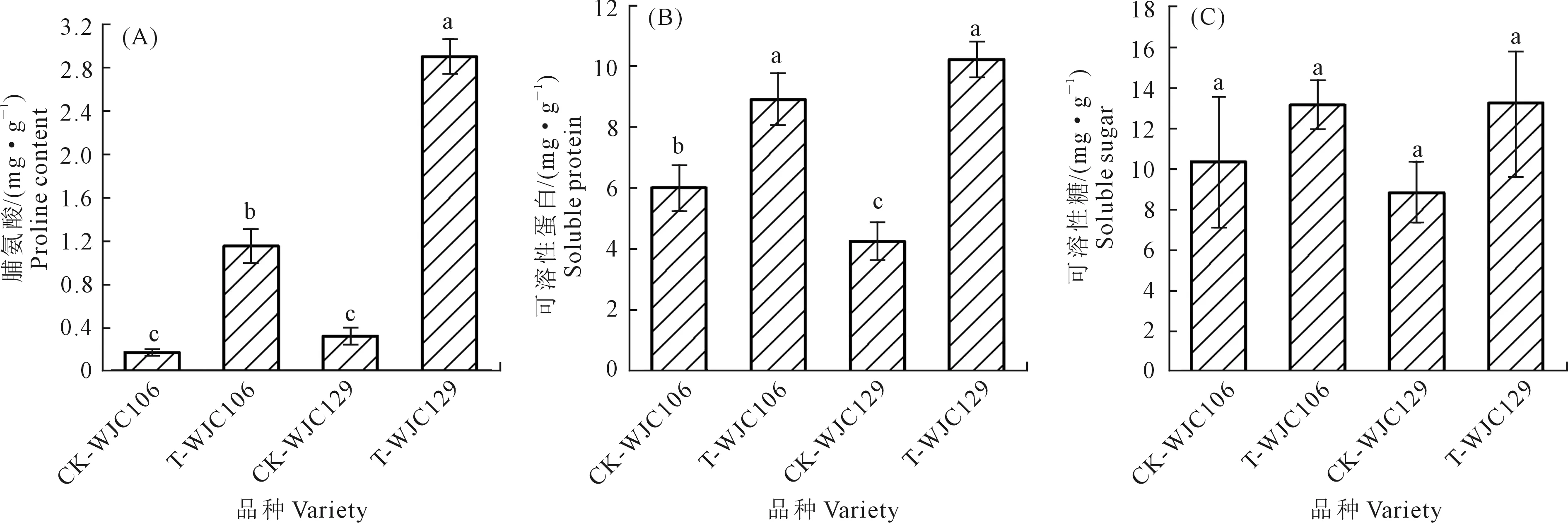
圖3 干旱脅迫對蕪菁苗期滲透調節物質含量的影響Fig.3 Effects of drought stress on contents of osmotic regulation substances of turnips seedling
2.4 干旱脅迫對不同抗旱類型蕪菁幼苗葉綠素含量及光合熒光參數的影響
如表1所示,干旱脅迫使兩個不同抗旱類型蕪菁苗期葉綠素含量顯著降低,且WJC129葉綠素含量高于WJC106。與對照組相比,兩個蕪菁處理組Pn和Tr呈現出顯著降低。干旱脅迫下WJC106的Gs值顯著低于對照組,而WJC129對照組與干旱組的Gs值無顯著差異。兩個不同抗旱類型蕪菁Ci變化量不盡相同,抗旱型WJC129蕪菁在干旱脅迫下呈現為下降趨勢,而干旱敏感型WJC106表現為顯著升高。兩個資源的水分利用效率變化量也有不同變化趨勢,在干旱脅迫的影響下,WJC106干旱敏感型蕪菁的WUE表現為顯著降低,而WJC129抗旱型資源出現顯著增加的趨勢。

表1 干旱脅迫對蕪菁苗期葉綠素含量及光合參數的影響Table 1 Effects of drought stress on chlorophyll and photosynthetic parameters of turnips seedling
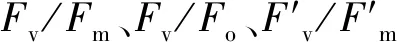
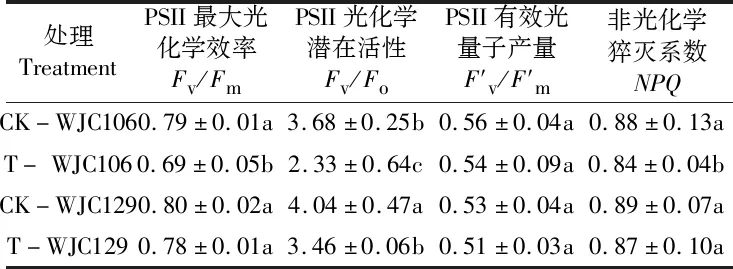
表2 干旱脅迫對蕪菁苗期熒光特性的影響Table 2 Effects of drought stress on fluorescence characteristics of turnips seedling
2.5 干旱脅迫對不同抗旱類型蕪菁苗期葉片氣孔形態的影響
表3為干旱脅迫對不同抗旱類型蕪菁苗期葉片氣孔特征值的影響,圖4為干旱脅迫對蕪菁苗期葉片氣孔形態的影響。由表3和圖4(A~D)可知,干旱脅迫使兩個不同抗旱類型的蕪菁苗期葉片氣孔張開長度、氣孔張開寬度、氣孔長度、氣孔寬度、保衛細胞厚度和氣孔開度極顯著降低。干旱脅迫下,WJC129抗旱型蕪菁苗期葉片氣孔開度的降幅顯著小于WJC106,降幅分別為65.90%和87.32%,并且氣孔張開長度、氣孔張開寬度、氣孔長度、氣孔寬度和保衛細胞厚度降幅也是WJC129明顯小于WJC106。表3與圖4(E~H)結合更直觀可知,干旱脅迫下WJC106蕪菁苗期葉片的氣孔張開率極顯著降低,而WJC129蕪菁無明顯差異。從氣孔密度上看,WJC106呈極顯著增加,WJC129無顯著差異。干旱敏感型WJC106蕪菁苗期葉片受干旱脅迫的損害較大。從圖4(C、D)和圖4(G、H)可以觀察到WJC129抗旱型蕪菁葉片表面有蠟質層覆蓋,蠟質晶體呈管狀形態。
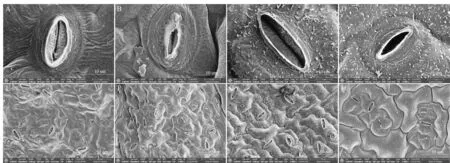
注:A為對照組WJC106蕪菁苗期葉片下表皮單個氣孔形態;B為干旱脅迫下WJC106蕪菁苗期葉片下表皮單個氣孔形態;C為對照組WJC129蕪菁苗期葉片下表皮單個氣孔形態;D為干旱脅迫下WJC129蕪菁苗期葉片下表皮單個氣孔形態;E為對照組WJC106蕪菁苗期葉片下表皮氣孔形態;F為干旱脅迫下WJC106蕪菁苗期葉片下表皮氣孔形態;G為對照組WJC129蕪菁苗期葉片下表皮氣孔形態;H為干旱脅迫下WJC129蕪菁苗期葉片下表皮氣孔形態。Note:B:A single stomatal morphology of the epidermis of the leaves at the seedling stage of turnip WJC106 in the control group;B:A single stomata morphology of the epidermis of the epidermis of the leaves at the seedling stage of turnip WJC106 under drought stress;C:A single stomatal morphology of the epidermis of the leaves at the seedling stage of turnip WJC129 in the control group;D:A single stomata morphology of the epidermis of the leaves at the seedling stage of turnip WJC129 under drought stress;E:The stomata morphology of the epidermis at the seedling stage of the control group WJC106 turnip;F:The stomata morphology of the epidermis of the leaves at the seedling stage of turnip WJC106 under drought stress;G:The stomata morphology of the epidermis of the leaves at the seedling stage of the control group WJC129 turnip;H:The stomata morphology of the epidermis of the leaves at the seedling stage of turnip WJC129 under drought stress.圖4 干旱脅迫對蕪菁苗期葉片氣孔特征的影響Fig 4 Effects of drought stress on stomatal eigenvalue of turnips seedling
3 討論與結論
環境在植物的生長發育過程中起著至關重要的作用,而水分因子更是占具主導地位[16]。本試驗表明,干旱脅迫使蕪菁苗期的丙二醛含量和相對電導率顯著增加,細胞膜結構受損,這與前人研究結果一致[11]。本試驗中WJC106干旱敏感型蕪菁苗期葉片的丙二醛含量和相對電導率的增量顯著高于WJC129抗旱型資源。干旱敏感型的蕪菁苗期細胞膜受損程度大于抗旱型,嚴重影響細胞中物質的進出,破壞細胞代謝的穩定性,同時表明蕪菁幼苗的抗旱性越強,電解質外滲量小,膜脂過氧化程度越低,其相對電導率和MDA積累量越低。
當植物受到干旱脅迫時,干旱信號會促進脯氨酸和可溶性糖等代謝物質的產生,激發抗氧化系統以維持氧化還原穩態,并通過過氧化物酶防止細胞損傷和膜完整性的破壞[17]。SOD、POD和CAT是保護酶系統中最重要的三種酶,其中SOD催化兩個超氧自由基發生歧化反應形成O2和H2O2,產生的H2O2再由POD和CAT分解除去[18]。三種酶協同作用有助于抵抗氧化損傷,維持細胞膜的穩定性和完整性。本研究中,干旱脅迫使兩個不同抗旱類型蕪菁苗期的SOD、POD和CAT活性增加,并且抗旱型蕪菁抗氧化酶活性的增幅顯著大于干旱敏感型。可溶性蛋白、可溶性糖和脯氨酸是植物主要的滲透調節物質,干旱脅迫下可通過滲透調節使植物維持一定的膨壓,從而維持細胞生長、氣孔開放和光合作用等生理過程、增強抗旱性[19]。本研究中兩個不同抗旱類型蕪菁苗期的脯氨酸、可溶性糖和可溶性蛋白含量在干旱條件下均表現出增加趨勢,但可溶性糖增幅差異不顯著,且抗旱型WJC129滲透調節物質含量高于干旱敏感型WJC106,表明蕪菁苗期可通過增加其滲透調節物質提高其抗旱性,且脯氨酸和可溶性蛋白在干旱脅迫下較為敏感,起主要的滲透調節作用。
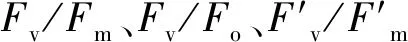
氣孔是植物進行氣體交換的重要器官,是水汽蒸騰和CO2進出的主要通道,受干旱影響,氣孔可能會關閉[24],以減少水分損失。本研究中干旱脅迫使兩個不同抗旱類型的蕪菁苗期葉片氣孔變小,保衛細胞變薄,氣孔開度顯著減小。抗旱型蕪菁氣孔張開率顯著大于干旱敏感型的,仍能維持較高的光合作用。干旱狀態下干旱敏感型WJC106蕪菁葉片表皮細胞出現重疊褶皺和起伏,影響氣孔開度和植物蒸騰,而抗旱型WJC129表皮細胞無明顯起伏,并且其表面附有蠟質層,蠟質晶體呈管狀形態。前人研究表明,蠟質是通過防止非氣孔性水分的流失,使得植物體蒸騰作用得以下降,從而能夠更好地維持植物體的水分來抵御干旱脅迫[25]。故抗旱型WJC129通過表面附有的蠟質層來增加其抗旱性。
綜上所述,干旱敏感型蕪菁苗期細胞膜受損程度大于抗旱型,且抗旱型蕪菁有較強的自我恢復與協調能力,通過增加抗氧化酶活性和滲透調節物質含量來抵御干旱脅迫。滲透調節物質中,脯氨酸和可溶性蛋白在干旱脅迫下較為敏感,起主要的調節作用。干旱脅迫降低蕪菁的光合熒光參數,氣孔特征值下降,光合機構活性降低;抗旱型蕪菁的各光合特性指標降幅均低于干旱敏感型,氣孔特征值也顯著大于干旱敏感型,故仍能維持較高的光合作用。抗旱型蕪菁表皮附有蠟質層,從而增加其抗旱性。
