基于SLAF-seq技術(shù)的蘋果SNP位點開發(fā)及遺傳多樣性分析
2023-05-23 15:10:19劉萬達(dá)楊光焦奎寶王昆王天鶴王禹吳立仁
江蘇農(nóng)業(yè)科學(xué) 2023年8期
劉萬達(dá) 楊光 焦奎寶 王昆 王天鶴 王禹 吳立仁
摘要:為了解黑龍江蘋果種質(zhì)資源的遺傳關(guān)系,利用特異性位點擴(kuò)增片段測序技術(shù)(SLAF-seq)對在黑龍江地區(qū)收集的31份蘋果種質(zhì)資源進(jìn)行SNP 位點開發(fā)和遺傳多樣性分析。最終獲得了107.52 Mb讀長數(shù)據(jù),不同材料的SNP標(biāo)記數(shù)目在 309 012~540 030 間,樣品測序質(zhì)量值平均Q30為93.78%,平均GC含量為40.90%。共開發(fā)獲得 1 072 115 個SLAF標(biāo)簽,其中,多態(tài)性SLAF標(biāo)簽有275 389 個。通過序列分析,共得群體SNP位點121 352 個,基于開發(fā)SNP分子標(biāo)記,結(jié)果表明,31 份蘋果種質(zhì)資源分為3 個亞群,特異性的蘋果SNP位點開發(fā)和研究可為蘋果種質(zhì)資源鑒定和遺傳多樣性分析提供理論基礎(chǔ)。
關(guān)鍵詞:蘋果;SLAF-seq技術(shù);SNP;遺傳多樣性分析
中圖分類號:S661.102 文獻(xiàn)標(biāo)志碼:A
文章編號:1002-1302(2023)08-0061-06
基金項目:農(nóng)業(yè)農(nóng)村部園藝作物種質(zhì)資源利用重點實驗室開放基金(編號:NYZS201906);黑龍江省自然科學(xué)基金聯(lián)合引導(dǎo)項目(編號:LH2022C097)。
作者簡介:劉萬達(dá)(1982—),男,黑龍江齊齊哈爾人,碩士,副研究員,主要從事寒地蘋果育種及栽培技術(shù)研究。E-mail:haaslwd@163.com。
蘋果(Malus pumila MILL.)是我國主要栽培果樹樹種之一,分布廣、品種多,具有很高的經(jīng)濟(jì)價值,與柑橘、葡萄、香蕉并稱為世界四大水果[1]。黑龍江省中小型蘋果由于干物質(zhì)含量高,口味純正,品質(zhì)佳,深受廣大消費者喜愛與認(rèn)可。這些蘋果種質(zhì)資源中較大部分是由栽培大蘋果品種與野生資源或地方品種雜交獲得。兩者基因的整合,獲得的蘋果資源既具有果實商品性又能夠適應(yīng)黑龍江省的環(huán)境。但經(jīng)過幾代雜交,近年黑龍江省培育的蘋果新品種存在果實品質(zhì)提高幅度不大、抗寒能力下降明顯、適宜栽植區(qū)域小等問題,育種工作進(jìn)展緩慢。地方品種是非常重要的種質(zhì)資源,因此,研究分析其種質(zhì)資源群體結(jié)構(gòu)及遺傳多樣性對資源利用、種質(zhì)鑒定及現(xiàn)代遺傳育種具有十分重要的意義。
特異性位點擴(kuò)增片段測序(SLAF-seq)是一種基于高通量測序技術(shù)發(fā)展而來的簡化基因組測序技術(shù),具有成本低、通量高、深度高、準(zhǔn)確性高等突出優(yōu)勢,已被廣泛用于作物遺傳圖譜構(gòu)建、遺傳多樣性分析、進(jìn)化關(guān)系分析和基因定位[2-4]。當(dāng)前,基于SLAF-seq技術(shù)在紫蘇、甘薯和金花茶[5-7]等作物上已開發(fā)大量的SNP標(biāo)記及遺傳進(jìn)化分析,但在黑龍江省蘋果種質(zhì)上進(jìn)行遺傳多樣性分析及大量SNP分子標(biāo)記開發(fā)等尚未見報道。本研究利用SLAF-seq技術(shù)對來自黑龍江省的31份蘋果種質(zhì)資源進(jìn)行遺傳進(jìn)化分析,探討黑龍江省蘋果種質(zhì)資源的遺傳背景及親緣關(guān)系,為黑龍江省蘋果品種的分類及鑒定奠定基礎(chǔ),為蘋果種質(zhì)資源的收集保存、開發(fā)利用及雜交組合選配提供科學(xué)的參考依據(jù)。
1 材料與方法
1.1 材料
試驗于2021年6月從黑龍江省不同地區(qū)采集31份蘋果種質(zhì)資源嫩葉置于變色硅膠中保存?zhèn)溆茫瑯悠吩敿?xì)信息見表1。
1.2 DNA的提取和檢測
供試蘋果樣本基因組DNA提取采用CTAB法,利用分光光度計檢測DNA的純度和濃度,利用1%瓊脂糖凝膠電泳檢測DNA完整性。
1.3 酶切預(yù)測
根據(jù)蘋果基因組大小和GC含量等信息,本研究選用蘋果基因組作為參考基因組(https://iris.angers.inra.fr/gddh13/the-apple-genome-downloads.html)進(jìn)行酶切預(yù)測。組裝獲得的基因組709.56 Mb,GC含量為38.03%。最終確定選擇 RsaⅠ+HaeⅢ為限制性內(nèi)切酶切組合,最后選取目的片段回收,為確保酶切實驗準(zhǔn)確性,本研究選取粳稻品種日本晴為對照進(jìn)行測序。利用Illumina平臺對質(zhì)量檢測合格后的文庫進(jìn)行高通量測序,得到個體序列,最后對測序質(zhì)量值 Q30和GC 含量進(jìn)行評估和分析以保證測序質(zhì)量。
1.4 SLAF 標(biāo)簽的獲得及SNP 標(biāo)記的開發(fā)
根據(jù)序列的相似性對31份蘋果種質(zhì)資源進(jìn)行聚類分析,一個 SLAF 標(biāo)簽如果在不同蘋果種質(zhì)間的序列有差異,即為多態(tài)性SLAF 標(biāo)簽。依據(jù) SLAF 標(biāo)簽開發(fā) SNP 位點,利用BWA[8]將測序結(jié)果和參考基因組序列比對,利用GATK[9]和SAM tools[10]對SNP標(biāo)記進(jìn)行開發(fā),最終選擇2種方法獲得的交集作為最可信的SNP標(biāo)記。根據(jù)缺失率<20%、次要基因型頻率(MAF)>5%的標(biāo)準(zhǔn)對所有的SNP位點進(jìn)行過濾篩選。
1.5 遺傳多樣性分析
利用Admixture軟件[11]對群體進(jìn)行群體結(jié)構(gòu)分析,利用MEGA5.0軟件[12],通過Neighbor-joining[13]算法,對群體進(jìn)行進(jìn)化樹和親緣關(guān)系分析,利用Eigensoft軟件[14]進(jìn)行主成分分析。
2 結(jié)果與分析
2.1 酶切方案與建庫評估
利用酶切預(yù)測軟件分析,選擇HaeⅢ+RsaⅠ為限制性內(nèi)切酶,酶切片段大小在364~464 bp的序列定義為SLAF標(biāo)簽,共預(yù)測到123 512個SLAF標(biāo)簽。利用SOAP軟件[15],將水稻日本晴的讀長與參考基因組進(jìn)行比對,結(jié)果(表2)表明,雙端比對效率為94.72%,測序數(shù)據(jù)酶切比例為83.40%,說明比對效率基本正常。根據(jù)獲得的SLAF標(biāo)簽的實際長度,繪制插入片段長度分布圖,結(jié)果(圖1)表明,讀長插入片段在預(yù)期范圍(364~464 bp)之內(nèi),表明SLAF建庫正常,可用于下步試驗。
由測序堿基分布情況(圖2)可知,堿基A與T、C與G分布較一致,符合堿基配對的原則,提示測序未出現(xiàn)錯誤,因此獲得SLAF標(biāo)簽可用于SNP標(biāo)記的開發(fā)和遺傳多樣性分析。
2.2 簡化基因組序列的產(chǎn)出和質(zhì)量評估
對31份蘋果種質(zhì)的測序數(shù)據(jù)統(tǒng)計結(jié)果,由圖3可知,共計獲得58.21個百萬讀長序列(Mreads)數(shù)據(jù),各樣品所獲得的讀長數(shù)為823 725~3 077 993,其中,來源黑龍江省哈爾濱市的品系“1962”樣品所獲得讀長數(shù)最大,為3 077 993個讀長,來源于黑龍江省雞西市的地方品種“一竄鈴”樣品所獲讀長數(shù)最小,為823 725個讀長(圖3-A)。測序質(zhì)量值Q30為92.93%~94.67%,測序平均Q30為93.50%,其中,來源于黑龍江的地方品種“花紅”樣品Q30測序值最大,為94.67%,來源于內(nèi)蒙古自治區(qū)的育成品種“塞外紅”樣品的Q30測序值最小,為92.93%。所有樣品Q30值均在90%以上,說明測序數(shù)據(jù)合格。測序獲得GC比例為40.09%~42.14%,其中,來源于黑龍江哈爾濱市的品系“245”樣品的GC比例最大,為42.14%,來源于黑龍江綏棱的地方品種“綏棱黃果”樣品的GC比例最小,為40.09%,平均GC含量為40.80%,GC比例普遍不高,說明達(dá)到測序要求(圖3-B)。
2.3 SLAF標(biāo)簽和SNP分子標(biāo)記的開發(fā)
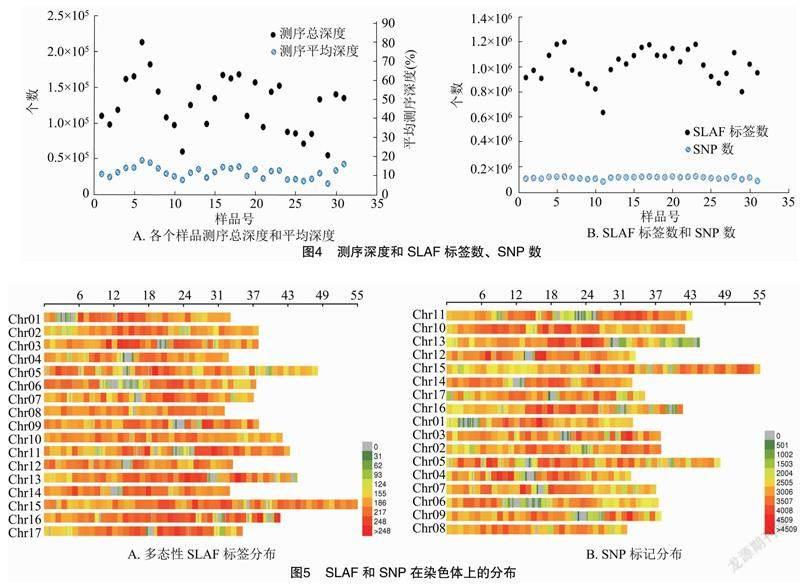
通過序列分析,共開發(fā)獲得了136 282個SLAF標(biāo)簽,其中多態(tài)性SLAF標(biāo)簽109 966個。由圖4可知,樣品的平均測序深度為11.29%(圖4-A),從多態(tài)性SLAF標(biāo)簽中開發(fā)獲得了2 039 575個SNP位點(圖4-B),對所有的SNP根據(jù)次要基因型頻率(MAF)>5%,缺失率<20%過濾,篩選高一致性的群體SNP。統(tǒng)計多態(tài)性SLAF標(biāo)簽和SNP標(biāo)記在蘋果不同染色體上的個數(shù),根據(jù)數(shù)據(jù)繪制染色體分布圖,由圖5可知開發(fā)的多態(tài)性SLAF標(biāo)簽和SNP位點在蘋果基因組17 條染色體上的分布情況,且在每條染色體上有比較均勻的分布,說明測序結(jié)果正常,可進(jìn)行下一步分析。
2.4 群體遺傳結(jié)構(gòu)分析
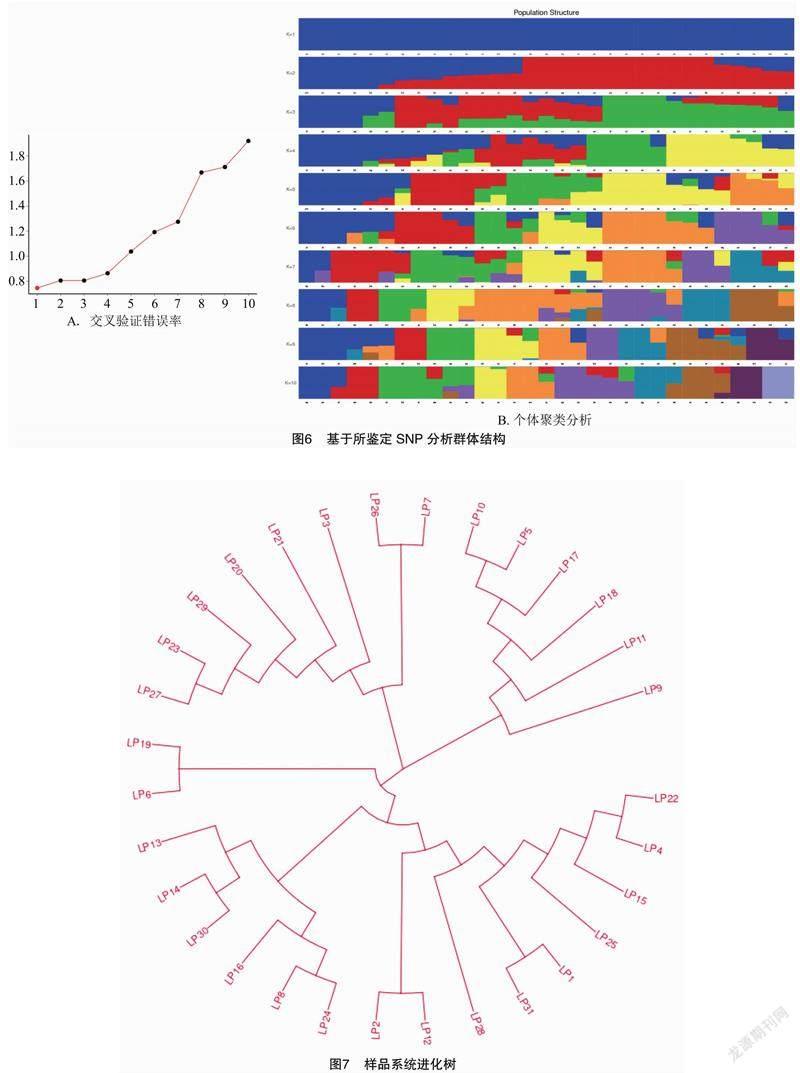
基于已開發(fā)的蘋果SNP位點,利用Admixture軟件分析了研究材料的群體結(jié)構(gòu)。根據(jù)交叉驗證錯誤率的谷值確定最優(yōu)分群數(shù)。由聚類分析結(jié)果(圖6)可知,K值為1時交叉驗證錯誤率最低,可見31份蘋果資源間遺傳結(jié)構(gòu)不明顯,說明不同來源的個體來源于同一祖先。
基于已開發(fā)的蘋果SNP位點,利用MEGA5軟件繪制31份蘋果種質(zhì)的遺傳關(guān)系聚類圖,由圖7可知,31份蘋果資源可劃分出3個差異性分支,第一個分支由17份蘋果資源組成,其中9份育成品種,4份地方品種,2份品系,2份其他。第二個分支由8份蘋果資源組成,其中2份育成品種,4份地方品種,1份品系,1份其他。第三個分支由6份蘋果資源組成,其中4份育成品種,2份品系。說明種質(zhì)間存在較大變異,具有豐富的遺傳多樣性。聚類結(jié)果表明黑龍江省蘋果資源沒有明顯的區(qū)域特征。
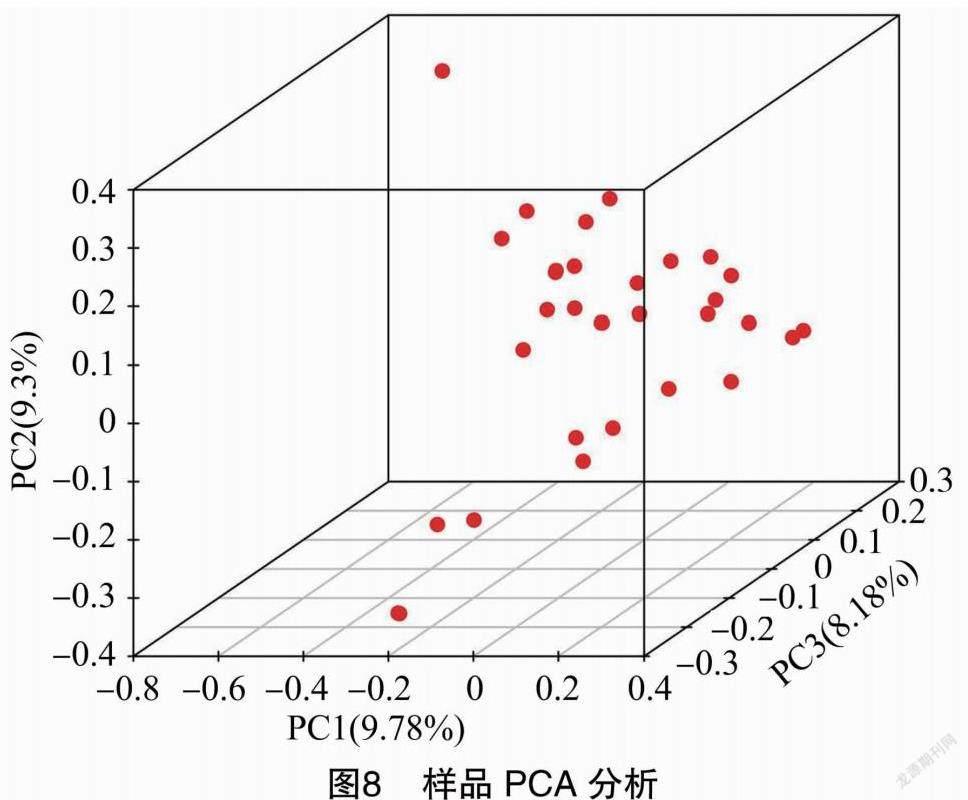
基于已開發(fā)的蘋果SNP位點,利用Eigensoft軟件,對31個蘋果種質(zhì)進(jìn)行主成分分析(PCA),得到樣品的聚類情況。由圖8可知,31份蘋果種質(zhì)聚為三維(PCA1、PCA2、PCA3),進(jìn)一步提示黑龍江蘋果31份種質(zhì)分化為3個亞群,黑龍江省蘋果資源品種之間存在遺傳多樣性。
3 討論與結(jié)論
蘋果是我國主要栽培果樹樹種之一,其面積和產(chǎn)量均居世界首位。目前,針對蘋果開展遺傳進(jìn)化分析研究,主要集中在生理生化水平和表型鑒定等方面。SLAF-seq技術(shù)具有快速、低成本、高通量開發(fā)SNP標(biāo)記的優(yōu)勢[16],國內(nèi)外研究者利用該技術(shù)已經(jīng)在廣金錢草、山西省地方梨、白皮松等作物上成功應(yīng)用[17-19],姜濤等運用SLAF-seq技術(shù)對39份連翹種質(zhì)資源進(jìn)行了SNP標(biāo)記的開發(fā)和遺傳多樣性分析,共獲得262 297個多態(tài)性SLAF標(biāo)簽,開發(fā)了1 809 741個SNP標(biāo)記[20]。陶紅霞等運用SLAF標(biāo)記,獲得125 497個多態(tài)性SLAF標(biāo)簽,構(gòu)建了蘋果遺傳連鎖圖譜[21]。李敏等通過SLAF-seq技術(shù)測序,獲得了99 526個多態(tài)性SLAF標(biāo)簽,并開發(fā)了9 488個高質(zhì)量的SNP標(biāo)記[22]。
植物的生長環(huán)境對物種的種群結(jié)構(gòu)和遺傳分化會產(chǎn)生直接的影響,通過蘋果樣品的系統(tǒng)進(jìn)化樹發(fā)現(xiàn),黑龍江蘋果種質(zhì)遺傳多樣性較為豐富,育成品種中“大秋”和“秋露”親緣關(guān)系比較近,地方品種中“凍果”和“山丁子”親緣關(guān)系較近,而地方品種“紅鈴鐺”和育成品種“七月鮮”親緣關(guān)系較近,上述品種之間親緣關(guān)系較近,可能是雜交育種產(chǎn)生的。本研究利用特異性位點擴(kuò)增片段測序技術(shù),通過對31份蘋果種質(zhì)資源進(jìn)行SNP 位點開發(fā)和遺傳多樣性分析,共得到 107.52 Mb Clean Reads 數(shù)據(jù),每個樣品的 SNP 標(biāo)記數(shù)目介于 309 012~540 030 之間,樣本平均測序質(zhì)量值 Q30 為93.78%,樣品平均 GC 含量為 40.90%。共得到 1 072 115 個 SLAF 標(biāo)簽,其中275 389 個多態(tài)性 SLAF 標(biāo)簽。通過序列分析,獲得 121 352 個有效的單核苷酸多態(tài)性標(biāo)記(SNP),利用開發(fā)的SNP分子標(biāo)記,將31份蘋果種質(zhì)資源分為3個亞群。本研究平均測序深度較高,獲得的SNP位點數(shù)量多,為今后結(jié)合蘋果果實性狀、葉片性狀等數(shù)量性狀進(jìn)行蘋果屬資源的評價利用,如親本選擇、關(guān)聯(lián)定位分析、雜種優(yōu)勢利用及進(jìn)化[23]等研究提供了重要的分子標(biāo)記輔助育種標(biāo)記,為蘋果種質(zhì)資源保護(hù)和品種創(chuàng)制等研究提供了有力證據(jù)。
參考文獻(xiàn):
[1]宋春暉,陳曉菲,王枚閣,等. 基于SLAF-seq技術(shù)鑒定蘋果砧木耐澇候選基因[J]. 中國農(nóng)業(yè)科學(xué),2021,54(18):3932-3944.
[2]Dong Z M,Chen L,Li Z,et al. Identification and molecular mapping of the semi-dwarf locus (sdf-1) in soybean by SLAF-seq method[J]. Euphytica,2020,216(6):103.
[3]Wei Q Z,Wang W H,Hu T H,et al. Construction of a SNP-based genetic map using SLAF-seq and QTL analysis of morphological traits in eggplant[J]. Frontiers in Genetics,2020,11:178.
[4]Zhang S Z,Hu X H,Miao H R,et al. QTL identification for seed weight and size based on a high-density SLAF-seq genetic map in peanut (Arachis hypogaea L.)[J]. BMC Plant Biology,2019,19(1):537.
[5]姜 濤,劉靈娣,田 偉,等. 紫蘇SNP分子標(biāo)記開發(fā)及遺傳多樣性分析[J]. 分子植物育種,2021,19(4):1243-1249.
[6]蘇文瑾,趙 寧,雷 劍,等. 基于SLAF-seq技術(shù)的甘薯SNP位點開發(fā)[J]. 中國農(nóng)業(yè)科學(xué),2016,49(1):27-47.
[7]劉 凱,李開祥,韋曉娟,等. 基于SLAF-seq技術(shù)的金花茶SNP標(biāo)記開發(fā)及遺傳分析[J]. 經(jīng)濟(jì)林研究,2019,37(3):79-83.
[8]Li H,Durbin R. Fast and accurate short read alignment with Burrows-Wheeler transform[J]. Bioinformatics,2009,25(14):1754-1760.
[9]McKenna A,Hanna M,Banks E,et al. The Genome Analysis Toolkit:a MapReduce framework for analyzing next-generation DNA sequencing data[J]. Genome Research,2010,20(9):1297-1303.
[10]Li H,Handsaker B,Wysoker A,et al. The sequence alignment/map format and SAMtools[J]. Bioinformatics,2009,25(16):2078-2079.
[11]Alexander D H,Novembre J,Lange K. Fast model-based estimation of ancestry in unrelated individuals[J]. Genome Research,2009,19(9):1655-1664.
[12]Tamura K,Peterson D,Peterson N,et al. MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J]. Molecular Biology and Evolution,2011,28(10):2731-2739.
[13]Saitou N,Nei M. The neighbor-joining method:a new method for reconstructing phylogenetic trees[J]. Molecular Biology and Evolution,1987,4(4):406-25.
[14]de Hoon M J L,Imoto S,Nolan J,et al. Open source clustering software[J]. Bioinformatics,2004,20(9):1453-1454.
[15]Li R Q,Yu C,Li Y R,et al. SOAP2:an improved ultrafast tool for short read alignment[J]. Bioinformatics,2009,25(15):1966-1967.
[16]段義忠,王建武,杜忠毓,等. 基于SLAF-seq簡化基因組技術(shù)的沙冬青SNP位點開發(fā)及遺傳分析[J]. 植物研究,2018,38(1):141-147.
[17]唐曉敏,張春榮,周良云,等. 基于SLAF-Seq技術(shù)的廣金錢草SNP位點開發(fā)及遺傳分析[J]. 分子植物育種,2020,18(18):6101-6107.
[18]白牡丹,郝國偉,張曉偉,等. 基于SLAF-seq技術(shù)的山西省地方梨品種的SNP分析[J]. 西北農(nóng)業(yè)學(xué)報,2020,29(7):1020-1027.
[19]田 倩,劉雙委,鈕世輝,等. 基于SLAF-seq技術(shù)的白皮松SNP分子標(biāo)記開發(fā)[J]. 北京林業(yè)大學(xué)學(xué)報,2021,43(8):1-8. [HJ2mm]
[20]姜 濤,溫春秀,田 偉,等. 基于SLAF-seq技術(shù)連翹SNP分子標(biāo)記開發(fā)及遺傳多樣性分析[J]. 分子植物育種,2021,19(16):5405-5413.
[21]陶紅霞. 基于SLAF標(biāo)記的蘋果遺傳連鎖圖譜構(gòu)建[D]. 楊凌:西北農(nóng)林科技大學(xué),2015.
[22]李 敏,郭 聰,王 瑩,等. 基于SLAF-seq技術(shù)的喬木柳SNP位點開發(fā)[J]. 西南農(nóng)業(yè)學(xué)報,2018,31(5):891-895.
[23]李曉穎,鄭少泉,徐紅霞,等. 基于SLAF-seq技術(shù)枇杷SNP位點開發(fā)[J]. 分子植物育種,2021,19(15):5038-5045.
