生鮮肉持水性機理研究進展
2023-04-06 03:02:52余青青高瑞昌包玉龍
食品科學 2023年5期
關鍵詞:結構
余青青,劉 嬌,洪 惠,高瑞昌,包玉龍,*
(1.江蘇大學食品與生物工程學院,江蘇 鎮江 212013;2.中南民族大學生命科學學院,湖北 武漢 430074;3.中國農業大學食品科學與營養工程學院,北京 100083)
新鮮瘦肉大約含有75%的水分,持水性是肉品在外力作用下,保持自身水分或后續加工中外加水分的能力。貯藏加工過程中,肉中水分處在動態變化之中,水分可以因蒸發、滲出和烹飪等過程損失,也可以在氯化鈉、多聚磷酸鹽、酸堿腌制液作用下而增加[1]。持水性是生鮮肉最重要的品質之一,肉品持水性不僅與感官品質和營養價值相關,也影響肉品的貯藏特性。質地、多汁性、顏色、滋味、營養素的保留以及微生物的生長均與肉品中水分含量及水分結合能力有關[2]。因此,肌肉的持水性能一直是肉品行業關注的重點。
有關肉品持水性機理的研究開展較早,20世紀60年代,Hamm[3]提出以肌原纖維溶脹為核心的肉品持水性機制,認為靜電相互作用是肉品持水的基礎;此后Offer等[1]從肌肉結構的角度進一步發展了Hamm提出的肌肉持水理論,指出滲透壓的形成對于肌肉水分的保持有重要作用;Tornberg[4]認為除上述因素外,肌肉內部存在許多毛細結構,而毛細管作用力可以截留大量的水分。生鮮肉宰后成熟過程中,僵直橫橋的形成以及內源酶導致的蛋白降解對于持水性有重要的調控作用。Kristensen等[5]研究發現,宰后成熟過程中,豬肉的持水性先下降后上升,由此提出細胞骨架蛋白在宰后成熟過程中的降解可以導致肌纖維外汁液回吸到肌纖維內部的猜想;Huff-Lonergan等[6]系統闡述了宰后僵直過程中肌纖維間和肌束間空隙對生鮮肉汁液流失的影響。隨著相關學科的發展,持水性的理論也在不斷更新,如Puolanne等[7]引入了低密度水、高密度水等全新的概念,對肉品持水性的理論基礎進行了深入探討。近年來,蛋白質氧化等化學修飾對于肉品持水性的影響也引起了廣泛的研究興趣[8-10]。本文首先闡述肌肉微觀結構和其中水分分布,接著介紹肉制品持水的理論基礎,并簡介常見的持水性測定和表征方法,最后詳細討論影響肉制品持水的理化因素及其可能的機制,以期為肉類研究提供理論基礎,為肉類產品的加工提供參考。
1 肌肉微觀結構及水分分布
肌肉組織的微觀結構高度有序,按照從宏觀到微觀的層級,可依次分為肌束、肌纖維、肌原纖維、肌絲(圖1A)。一般而言,水分被緊密束縛在肌肉各個結構單元內,約占新鮮肌肉質量的75%,其中約85%的水分位于肌纖維內部,被束縛在由粗細肌絲組成的稠密的肌原纖維網絡之中,而另外約15%的水分則位于肌纖維外部。肌肉中的水分空間分布位置包括:肌原纖維內與肌原纖維之間、肌原纖維和肌纖維膜之間、肌纖維之間、肌束之間。肌肉中的水分根據流動性可分為結合水、不易流動水、自由水(圖1A)。場核磁共振技術通過檢測氫質子弛豫時間證明了這3 種狀態水分的分布[11-13]。結合水由于與蛋白質表面有很強的相互作用而擁有非常穩定的性質,只有在煅燒的情況下才會解離,因此不易受到外界因素,如剪切力、重力、加熱或冷凍等的影響。結合水僅占為總水分含量的很小一部分,大約每克肌肉蛋白中含0.5 g結合水[14],相當于鮮肉質量的8%左右。也有學者對此持不同觀點,認為肌肉中只有大約0.5%的水分是真正的結合水[15]。結合水之外的水分根據所處位置分為不易流動水和自由水,不易流動水又稱截留水,位于肌原纖維之內,而自由水主要分布在肌原纖維之間以及肌纖維外部[16]。肌肉在宰后成熟過程中,橫橋連接的形成、骨架蛋白的降解、蛋白質變性等原因導致微觀結構變化,從而改變細胞內外空間形成的汁液流失通道,進而影響持水性[5,17-18]。
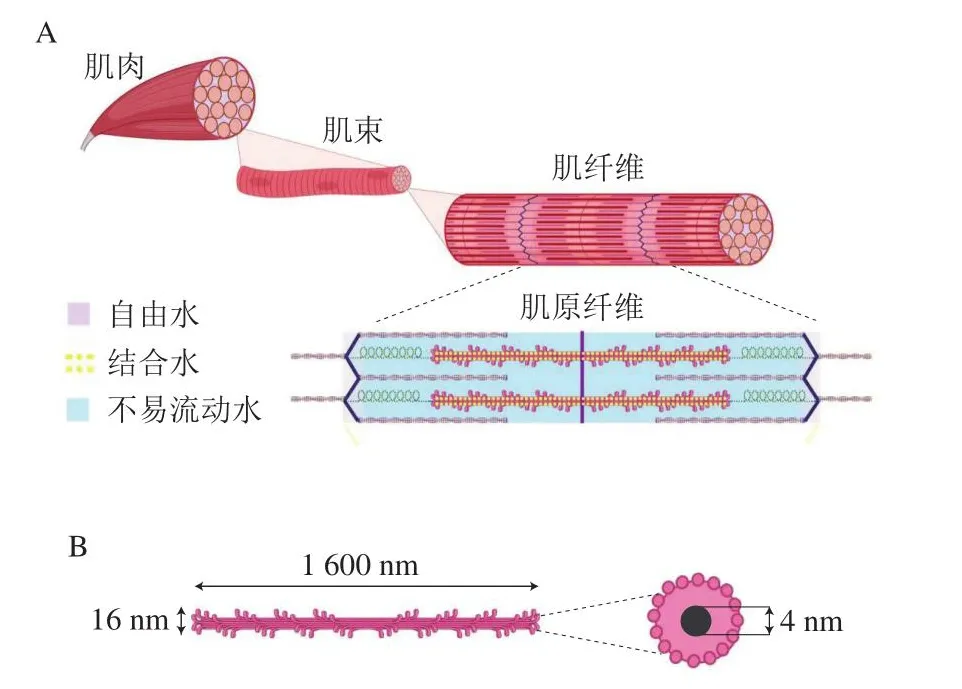
圖1 肌肉微觀結構及水分分布示意圖[7,16]Fig.1 Illustration of the microstructure of muscle and the distribution of water in muscle matrix[7,16]
盡管已有大量實驗研究了肌肉微觀結構及其對肉類品質的影響[19-20],但在更細微尺度上,如粗細肌絲結構與不易流動水間的關系仍然需要進一步的探明。粗肌絲(以肌球蛋白為主)大約含有200~300 個肌球蛋白分子,構成了約16 nm×1 600 nm的纖維[21]。有學者指出,粗肌絲中心含一個直徑約4 nm的中空部分[22](圖1B),雖然這一結構尚未得到證實,但粗肌絲中心的中空部分很可能與肉品持水性有很大關聯。隨著超分辨顯微鏡等技術的出現[23],肌肉微觀結構有望進一步探明。
隨著對于肌肉結構的認知不斷加深,科研人員對肌肉內水分的理化性質也有了新的發現,有助于更好地理解肉品的持水性。對于肌纖維內的水分,有學者認為由于排布高度結構化,水分的滲透壓和流動性都與普通的體相水不同,有時甚至表現為一種類玻璃態[24]。離子和其他一些物質根據其與水分子相互作用的特點,可以分為親液型(水結構增強)和離液型(水結構破壞)。親液基團(包括多價離子和半徑較小的離子)誘導水分子排列成有序結構,形成類似玻璃態的狀態,稱為低密度水。相反地,離液基團(大的單價離子)誘導水分子呈無序結構,形成高密度水(圖2)。Puolanne等[7]猜測高密度水具有更低的黏度和更好的流動性,與臨近肌肉結構的結合較為松散,因而其含量越高則持水性越低。已有研究表明肌肉中存在高密度水,如肌動蛋白絲附近的水化層[25]。但目前關于高密度水/低密度水對肉品持水性的影響還有待進一步的闡明。
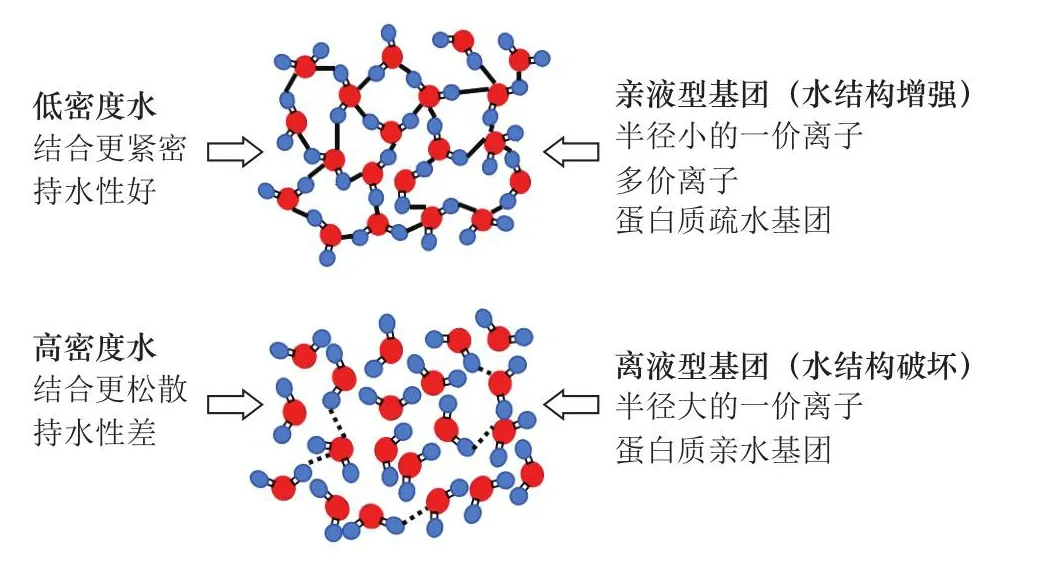
圖2 高密度水、低密度水的示意圖[7]Fig.2 Illustration of high-density and low-density water[7]
2 肉制品持水性理論
在肉品持水性理論研究過程中,除了蛋白質與其鄰近的幾層水分子的關系,肌原纖維蛋白網絡及基質蛋白網絡體系如何維系其中的水分也十分重要。目前肉品持水作用力的基礎理論主要包括粗細肌絲之間的靜電作用力[3,26]、滲透壓[1]和毛細作用力[27]。各個理論側重點不同,尚無單一理論能完美解釋不同條件下的肉品持水現象。
2.1 靜電作用力
蛋白質在偏離自身等電點時,表面會攜帶有凈電荷。Hamm[3]指出,肌原纖維蛋白或肌絲之間的靜電斥力會造成肌原纖維的溶脹,而肌肉微觀結構中一些橫向連接(Z線、肌動球蛋白等)會限制肌原纖維的溶脹,與靜電斥力達到平衡。在肌纖維層面,肌原纖維與肌纖維膜之間的連接以及肌纖維外部的肌內膜也會限制肌纖維的溶脹。根據Hamm的觀點,肉品持水性由靜電斥力和橫向連接作用共同決定。水分子在肌絲蛋白之間受到吸引(極性基團)和排斥(疏水基團)的共同作用,在肌原纖維網絡之中形成一種類似冰結構的水分子簇。靜電作用力理論能夠很好地解釋諸多因素,如pH值、鹽含量等對于肉品持水性的影響。
2.2 滲透壓
與Hamm的觀點略有不同,Offer等[1]認為由于鄰近肌絲之間的間距大約在15~20 nm,這一間距太長而不足以產生足夠強的靜電斥力。他們各提出了基于滲透壓的肉品持水理論。Hamm在解釋氯化鈉對于肉品持水性的促進作用時提出帶負電的氯離子會選擇性地結合到肌絲上,增加肌絲的負電荷,增大靜電斥力;Offer等認同氯離子選擇性結合到肌絲,但由于肌絲不能自由移動,靜電相互作用會導致帶正電荷的離子(即鈉離子)向肌絲附近富集,在水相中形成不均勻的離子分布。這一不均勻的離子分布在肌絲之間形成滲透壓差異,從而導致水分子進入,引起肌原纖維溶脹。同樣地,肌肉結構中的橫向橋聯限制了肌原纖維的溶脹,與滲透作用達到平衡。
2.3 毛細作用力
肌絲之間的間距很小,這一距離理論上可以產生約300 m水柱的毛細作用力[28],因此,有學者指出毛細作用力是維系肉品持水性的重要因素[4]。但Puolanne等[7]指出肌肉宰后成熟過程中,肌絲間距的減小理論上會導致毛細作用增強,從而增強持水性,然而實際體系持水性卻隨著肌絲間距的減小而減小。他們指出毛細作用可能僅適用于肌纖維外的水分保持,而且在復雜的肌原纖維網絡之中,很難去界定毛細管孔徑。
3 持水性的表征方法
持水性的測定通常需要向肉品施加作用力,使水分從體系中釋放,這一作用力可以是重力,也可以借助擠壓、離心等外力,然后通過測定肌肉中自由水在外力作用下的損失量,表征肌肉的持水性[29]。不同的持水性測定及表征方法基于的原理略有不同,清晰地了解和區分這些不同的持水性測定表征方法對于理解實驗中觀測到的持水性現象十分重要。
3.1 滴水損失及汁液流失
滴水損失是指肉品在不同環境條件下汁液滴落造成的質量損失。最常見的測定方法是將肉塊(約30~100 g)懸掛在袋子中,并在冷藏條件下貯藏1~2 d,確保肉和袋子不接觸且方便汁液滴落。最終稱量貯藏前后的肉塊質量,從而計算得出滴水損失[30]。也可以將肉塊切成小塊放置在托盤,收集汁液后計算,但肉的表面積會對結果有影響。這一方法因其簡便性而被廣泛使用。汁液流失與滴水損失類似,指的是肉類在貯藏、零售期間在包裝容器中損失的水分。典型的汁液流失量大約占新鮮瘦肉質量的2%~6%,凍融循環、宰前應激(可能導致PSE(pale, soft, exudative)肉)等過程會增大水分流失[1]。
3.2 蒸煮損失
蒸煮損失是常見的肉類持水性指標,對肉塊蒸煮前后稱質量即可計算蒸煮損失。蒸煮損失受到肉類宰后成熟、蒸煮溫度、蒸煮條件等影響,一般不超過40%[1]。在所有表征持水性的方法中,蒸煮損失與肉類的多汁性相關度最高[29]。
3.3 濾紙擠壓法
該方法是將肉塊切碎,稱取固定質量(如300 mg)并放置在濾紙中心,利用重物(如200 g)持續擠壓約5 min,計算濾紙前后的質量變化,或者計算濾紙上汁液潤濕的面積和肉泥的面積之差,Wierbicki等[31]選取了部分肉類進行研究,得出可被擠出的水分占肉中總水分的30%~50%。該方法很早就已經被使用,近年來有研究人員將圖像采集系統和分析系統整合到該方法中,提出了持水性快速測定方法,并命名為WHC trend[32]。
3.4 離心法
離心法是借助離心力分離肉類中的部分水分,需要確保離心損失的水分可以被收集在離心管的底部而不與肉接觸,以避免重新被吸收。這一方法簡便,并且與汁液損失測定原理類似,同時測定結果相關度高,因此被廣泛采用。赫爾辛基大學的Ertbjerg教授研究團隊[19]擴展了此方法的應用,發現對提取的肌原纖維進行離心同樣可以用來表征不同情形下肉類體系的持水性,每克肌原纖維離心之后截留的水分大約在5~9 g[33-34]。
3.5 肌原纖維溶脹以及肌肉水合能力
3.1~3.4 節中介紹的方法均是通過測定水分損失表征肉品持水性,而也有學者采用肌原纖維的溶脹以及肌肉的水合能力來表征肉品持水性。肌原纖維的溶脹被廣泛用作肉品持水性的理論模型。利用相差顯微鏡可以直接觀測提取的肌原纖維的形態,肌原纖維的寬度反映了肌原纖維的橫向溶脹狀態。而將肉塊浸泡在溶液中,稱量浸泡前后的質量變化即可反映肉品持水的特性[8]。
3.6 無損檢測法
上述持水性測定或表征方法都需要破壞肉類結構,一些快速、無損的方法越來越受到研究人員和肉品行業的歡迎。研究表明低場核磁共振的橫向弛豫時間(T2)可以有效地測定肉類中的自由水[13]。Kamruzzaman等[35]采集了豬肉、牛肉、羊肉的高光譜圖像,成功建立了預測紅肉持水性的模型。Kaewthong等[36]成功地利用了電導率來評價雞胸肉的持水性。
4 影響肉品持水性的因素
蛋白質凈電荷以及肌肉的微觀結構是影響持水性的兩大重要內在因素[15],肉品生產過程中如宰前應激、胴體冷卻、冷藏、冷凍/解凍、腌制等外部因素均可以通過影響蛋白質凈電荷或肌肉微觀結構而影響其持水性。
4.1 蛋白質凈電荷
蛋白質上部分氨基酸殘基(Lys、Arg、His、Asp、Glu等)以及碳末端羧基和氮末端的氨基可以發生解離從而攜帶電荷(圖3)。肉類在宰后成熟過程中,隨著組織內乳酸的積累,pH值會下降。一般豬肉、牛肉的最終pH值會略高于肌原纖維蛋白整體的等電點(pH 5.0左右)[1,37],因此肌原纖維蛋白整體攜帶凈的負電荷。研究表明,肌絲凈電荷的多少影響肌絲間距并最終影響肉品的持水性。圖3展示了不同pH值條件對肌絲電荷以及肌原纖維蛋白體系持水性的影響[9]。在pH值接近等電點時,肌絲因含有大致相同量的正負電荷而凈電荷接近0。通過加酸加減都可以使pH值偏離等電點而增加肌絲的凈電荷(表現為圖3中肌原纖維可以結合更多的帶電荷染料)。其中pH值增大導致組氨酸的解離平衡主要以不帶電荷的形式存在,相反地,pH值減小導致谷氨酸、天冬氨酸等解離平衡時主要以不帶電荷的形式存在,兩種情形下蛋白質的整體凈電荷都會增大,從而增大了肌絲間的靜電互斥[26]或者造成反離子的不均勻分布導致滲透壓差[1],使得肌絲間距變大,最終增強了持水性。當pH值進一步增大或者降低時,由于體系酸性或堿性過大,導致肌原纖維網絡體系崩塌而造成持水性下降。NaCl是肉類加工中常用的添加物,當添加NaCl時,由于氯離子選擇性與粗肌絲結合,當肌絲本身攜帶凈負電荷時,氯離子的加入會增加總凈負電荷,從而增強靜電排斥,肌絲間距增大,持水性升高;相反地,當肌絲攜帶凈正電荷時,氯離子的加入會減少總凈電荷,造成持水性下降。由于肌絲凈電荷的載體為肌原纖維蛋白,蛋白質理化性質的變化也可能影響凈電荷,比如Bao Yulong等[33]在研究模擬氧化體系下肌原纖維的持水性時發現氧化增大了持水性,推測可能是蛋白質組氨酸殘基的氧化生成羰基化合物2-oxo-histidine,從而失去攜帶的正電荷,導致凈的負電荷增加。主要肌原纖維蛋白凈電荷的增加也在等電點電泳結果中得到證明[33];而肌漿蛋白的變性導致溶解度下降,可能會沉積到肌原纖維蛋白表面從而遮蔽部分電荷[16]。
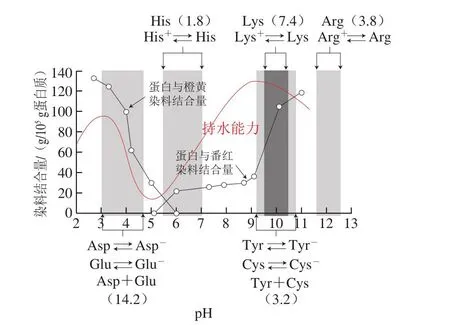
圖3 pH值對肌絲電荷以及肌原纖維蛋白體系持水性的影響[9]Fig.3 Effect of pH on myofilament charge and the water-holding capacity of myofibrillar protein[9]
由于pH值和氨基酸基團的pKa值共同決定側鏈基團的解離情況,因此pH值是影響肌肉蛋白凈電荷從而影響其持水性的重要原因。動物的宰前應激影響肌肉內糖原含量以及糖酵解的速率從而影響最終pH值,如持水性良好DFD肉的形成與宰前糖原過度消耗導致肉類最終pH值偏高有關。而胴體的冷卻過慢雖然不改變肌肉最終pH值,卻影響pH值下降的速率,如持水性差的PSE肉的形成與胴體pH值下降過快有關[1]。在腌制過程中,一些可食用酸堿[38]、磷酸鹽[39]的加入都有可能改變體系的pH值從而改變持水性。在肉品冷凍過程中,也涉及pH值的變化,伴隨著肌肉中水分的凍結,溶質濃縮效應導致各類離子(包括H+)濃度增加,從而改變未凍結水分的pH值。Zhang Yuemei等[34]等猜測在冷凍時,H+可能會被部分地束縛在冰晶中從而影響體系局部的pH值,相較于慢速冷凍,快速冷凍可能導致H+更多地被截留在冰晶中,從而未凍結部分pH值更高。此外,冷凍電勢可能也是影響冷凍肉類未凍結部分液體pH值變化的原因,冷凍產生的電勢會被體系中移動性很強的H+或OH-中和,最終導致局部pH值發生變化[40](圖4)。
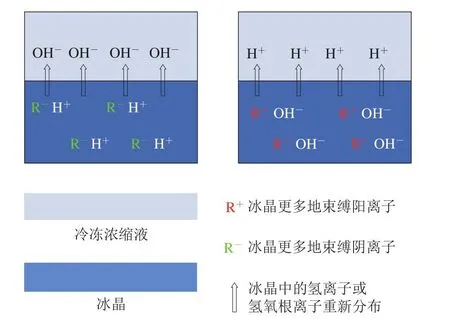
圖4 冷凍過程中冰水兩相電勢介導的pH值變化機理示意圖[40]Fig.4 Schematic illustration of pH changes in frozen electrolyte solutions mediated by freezing potential[40]
4.2 微觀結構
肌纖維的結構變化是凈電荷之外又一影響肌肉持水性的重要因素。肌纖維結構決定了肌肉中水分分布,也限制了肌原纖維/肌纖維的過度溶脹。肌小節是肉類的基本結構單元,宰后肌動球蛋白橫橋的形成減少了能容納的水分,多聚磷酸鹽的加入可以破壞肌動球蛋白的連接從而增強持水性[41]。除此之外,肌小節的橫向收縮也會導致肌原纖維的體積減小,擠出部分水分[18]。肌原纖維通過肌間線蛋白、紐帶蛋白等與細胞膜相連接,在加工或貯藏過程中,如果連接蛋白降解充分,肌原纖維失去與細胞膜的連接,則肌原纖維的收縮將不會聯動肌纖維整體收縮,從而減少了水分的流失;如果連接蛋白的降解不充分,則這些連接仍然存在,肌原纖維的收縮可以借助這些連接傳遞到整個肌纖維,從而使肌細胞直徑減小將水分排出細胞外。由于細胞外的水分更容易流失,因此肉品持水性下降(圖5)。肉的宰后成熟過程往往伴隨著細胞骨架蛋白在內源酶的作用下降解,而連接蛋白的降解減少了收縮的肌原纖維對于肌纖維整體的牽制,肌纖維重新溶脹,肌細胞直徑增大使肉品持水性在宰后成熟之后變好[5-6]。Zeng Zhen等[42]研究發現,肌原纖維Z線附近的輔肌動蛋白可以被鈣蛋白酶降解,從而導致Z線弱化,減少對肌原纖維橫向溶脹的限制,肌原纖維的持水性得以提升(圖5)。

圖5 肉品空間位阻效應對持水性影響的機理示意圖[6,42-43]Fig.5 Illustration of the mechanism of changes in water-holding capacity of muscle due to steric effect[6,42-43]
生鮮肉在貯藏加工過程中(如含氧包裝、輻照、冷凍等),其蛋白質容易發生氧化,而已有的研究表明蛋白的氧化修飾也可以通過影響肉品微觀結構從而影響持水性[44]。蛋白質氧化影響肉的持水性主要有兩種機制:1)肉品中主要的蛋白水解酶——μ-鈣蛋白酶在宰后早期可能因發生氧化而鈍化,減弱了其對一些細胞骨架蛋白的降解能力;2)氧化促使蛋白分子間形成二硫鍵或羰基介導的交聯從而強化了粗絲的結構完整性,限制了肌原纖維粗絲的橫向溶脹(圖5),造成持水性下降[45]。
根據Hamm[26]的觀點,肌原纖維蛋白對肉品持水性的貢獻約占50%,肌漿蛋白僅占3%左右。最近研究表明肌漿蛋白在特定條件下也參與肉品中微觀結構的形成,從而影響持水性[46]。在類PSE肉中,肌漿蛋白的沉積與肌絲間距的減小同時發生,變性的肌漿蛋白在細胞內肌原纖維外以及肌纖維外發生沉積,形成了網絡結構,從而部分抵消了肌原纖維蛋白變性對持水性的下降作用[17]。最新的研究也進一步證實了在類PSE肉中,肌漿蛋白的變性提升了肌原纖維的持水性[46]。肌肉中結締組織的存在也是限制肌肉溶脹的因素,比如完整的肌內膜會限制肌纖維的過度溶脹。目前,肌肉中研究比較成熟的是肌原纖維蛋白以及肌漿蛋白,關于膜蛋白以及其他低豐度的骨架蛋白對于持水性影響的研究比較有限。隨著蛋白組學技術在食品科學領域的應用愈加廣泛,通過蛋白組學技術可以發現高持水性的生物標志物,從而為研究持水性的調控機制提供更多思路[43,47-51]。Liu Yueyue等[48]研究發現冷凍鳙魚肉持水性的下降伴隨著蛋白質分子的氧化,而最主要的氧化修飾包括與脂肪氧化產物丙二醛和4-羥基壬烯醛(4-hydroxynonenal,HNE)的加成。Zhang Muhan等[50]發現高持水性鵝肉與低持水性鵝肉相比,差異蛋白主要集中在結構蛋白、代謝酶、抗氧化酶、應激蛋白;Zuo Huixin等[51]指出肌球蛋白輕鏈、熱休克蛋白HSP27、磷酸丙糖異構酶可以作為預測牦牛肉高持水性的生物標志物。然而,差異蛋白只能體現含量上的差異,對于蛋白是否變性、酶解程度等指標則很難做出區分,而這些又是影響持水性的重要因素。隨著組學技術的進步,上述蛋白變性等問題也將迎來更加全面且合理的解決方案。如已有研究人員開發出限制性酶解-液相色譜-質譜聯用分析技術,從而實現對蛋白質結構變化的高通量篩選[52]。
除了上述提及的蛋白酶解、氧化、變性等,其他一些物理性的影響因素也會導致肌肉微觀結構發生變化。生鮮肉類的汁液主要從切口處流失,肉塊的大小、切口與肌纖維方向的關系、支撐方式(懸掛、或放置在臺面)都可能通過改變汁液流失通道的結構從而影響持水性[1]。冷凍/解凍過程冰晶的物理破壞作用也是影響肉品持水性的一大因素。對于慢速冷凍的肉品,冰晶大且主要分布在細胞外,凍藏過程中溫度波動造成的重結晶使冰晶體積進一步增加,大冰晶會破壞肉的微觀結構,造成解凍時更多的汁液流失[53]。
5 結 語
學術界關于肉品持水性變化機理方面的研究開展較早,各個模型體系下肉品持水的理論基礎已經得到了很好地闡明。然而,實際肉品體系中所涉及到的眾多理化因素(pH值、離子強度及類型、蛋白酶解、蛋白變性等)大多處于一個動態平衡的過程。肉品體系的一大特點是很難預測任何單一因素對肉品品質產生的確切影響。因此,肉品持水性機理仍然有待進一步闡明。隨著蛋白質化學、水的功能特性研究的進步和定量蛋白組學、超分辨顯微鏡等高新技術的發展,肉制品持水性的理論研究將會取得新的成果。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
