甾醇類化合物降低高膽固醇HepG2細胞內膽固醇的作用機制
2023-04-06 03:02:38謝建華郭小妹袁蘭蘭申明月
食品科學 2023年5期
關鍵詞:水平
謝建華,郭小妹,袁蘭蘭,余 強,陳 奕,申明月*
(南昌大學 食品科學與技術國家重點實驗室,江西 南昌 330047)
甾醇類化合物是一類來自于植物或真菌的天然活性物質,其主要膳食來源是植物油、谷物、堅果、豆類和食用菌等。這些甾醇類化合物的結構與膽固醇相似,但僅在植物或真菌中合成。甾醇類化合物具有許多重要的生物活性,例如降膽固醇、抗炎、抗腫瘤、抗菌等[1-2],其中降膽固醇作用尤為明顯,被稱為“膽固醇的克星”,國內外關于甾醇類化合物降膽固醇的研究也越來越多。研究表明甾醇類化合物可以顯著降低血清總膽固醇(total cholesterol,TC)、血清甘油三酯(triglycerides,TG)、肝脂水平和致動脈粥樣硬化指數(即高密度脂蛋白膽固醇(high-density lipoprotein cholesterol,HDL-C)/低密度脂蛋白膽固醇(low-density lipoprotein cholesterol,LDL-C))[3-5],促進膽固醇排出,增加小鼠糞便膽固醇含量[6]。
近年來,由于不良的飲食習慣和生活方式,人群血脂水平明顯增加。血脂異常是以人體內脂蛋白代謝異常為特征,主要包括TC、LDL-C、TG水平的升高和/或HDL-C水平的降低等。血脂異常是心血管疾病的主要誘因和危險因素。人體內的血漿膽固醇穩態主要受肝臟內源性合成、腸道對膳食膽固醇的吸收以及膽汁的清除和排泄控制。在過去的幾十年里,科學家們已達成共識,營養和飲食管理是預防血脂異常,從而最大限度降低心血管疾病風險的關鍵策略[7]。增加富含甾醇類食物的攝入有助于改善血脂水平,降低膽固醇,預防心血管疾病。
目前,關于甾醇類化合物降膽固醇活性的研究主要集中于植物甾醇如β-谷甾醇、豆甾醇、菜油甾醇等,對于來源于食用菌類的甾醇研究很少,且大多都停留在表象指標研究階段,其作用機理仍不明確。本研究以麥角甾醇、麥角甾醇酯、(22E)-麥角甾-4,6,8(14),22-四烯-3-酮((22E)-ergosta-4,6,8(14),22-tetrean-3-one,(22E)-Ergosta)、星魚甾醇為研究對象,通過構建高膽固醇HepG2細胞模型,探究其對膽固醇水平的影響及其作用機制。
1 材料與方法
1.1 材料與試劑
人肝腫瘤細胞系HepG2來源于中國科學院細胞庫。
麥角甾醇(批號:E0018,純度>95%) 梯希愛(上海)化成工業發展有限公司;麥角甾醇酯(純度97%,批號:12-CF-135-3)、(22E)-Ergosta(純度98%,批號:4-EKP-158-1)、星魚甾醇(純度94%,批號:7-VHP-132-1) 加拿大Toronto Research Chemicals公司;DMEM培養基、油紅O染色液(細胞專用) 北京索萊寶科技有限公司;胎牛血清 以色列Biological Industries公司;膽固醇、25-羥基膽固醇、辛伐他汀上海阿拉丁生化科技股份有限公司;CCK8試劑盒日本同仁化學研究所;細胞裂解液、細胞總蛋白提取試劑盒、ECL化學發光試劑盒 上海碧云天生物技術研究所;兔單克隆尼曼-匹克C1型類似蛋白1(Niemann-Pick type C1-like 1,NPC1L1)抗體、兔單克隆ATP結合盒轉運體G5(ATP-binding cassette transporter G5,ABCG5)抗體、兔單克隆固醇調節元件結合蛋白2(sterolregulatory element binding protein 2,SREBP2)抗體、小鼠β-actin抗體 美國Abcam公司;兔單克隆ABCG8抗體、兔單克隆膽固醇酰基轉移酶2(acetyl-coenzyme A acetyltransferase,ACAT2)抗體、兔單克隆羥甲基戊二酸單酰輔酶A合成酶(3-hydroxy-3-methylglutarylcoenzyme A synthase,HMGCS1)抗體 美國Cell Signaling Technology公司;辣根過氧化物酶標記山羊抗鼠、山羊抗兔免疫球蛋白G(immunoglobulin G,IgG)二抗 北京中杉金橋生物技術有限公司;TG、總蛋白定量檢測試劑盒 南京建成生物工程研究所。
1.2 儀器與設備
MULTISKAN MK3酶標儀、HERACELL 150i二氧化碳細胞恒溫培養箱 美國Thermo Scientific公司;Research plus單道固定量程移液器 德國艾本德公司;CKX53倒置顯微鏡 日本Olympus公司;GelDoc XR Biorad凝膠成像系統、Mini-PROTEAN Tetra電泳儀、1704150全能型蛋白轉印系統 美國Bio-Rad公司;1260超高效液相色譜儀 美國Agilent Technologies公司。
1.3 方法
1.3.1 HepG2細胞的培養及實驗分組
HepG2細胞培養于含10%胎牛血清的DMEM培養基,每2~3 d按照1∶3比例傳代,取對數生長期細胞經胰蛋白酶消化,按2×105個/孔接種于6 孔板。實驗分為4 組:空白對照組:加入2 mL無血清培養基;模型組:加入2 mL無血清培養基配制的膽固醇(10 μg/mL)和25-羥膽固醇(1 μg/mL)混合溶液[8-9];陽性對照組:加入2 mL以無血清培養基配制的膽固醇(10 μg/mL)、25-羥膽固醇(1 μg/mL)和辛伐他汀(10 μmol/L)混合溶液;甾醇類化合物處理組:加入2 mL無血清培養基配制的膽固醇(10 μg/mL)、25-羥膽固醇(1 μg/mL)及不同濃度甾醇類化合物(25、50、100 μmol/L)的混合溶液。各組細胞培養24 h后用于后續實驗。
1.3.2 細胞存活率測定
將對數生長期HepG2細胞(1×104個/孔)接種于96 孔板。細胞貼壁后,無血清培養基同化處理12 h,然后分別加入200 μL濃度為25、50、100 μmol/L的各甾醇溶液處理24 h,按照CCK8試劑盒說明書測定光密度值(OD450nm)。以不加甾醇溶液為空白對照組,以不接種細胞、不加甾醇溶液為空白,根據下式計算細胞存活率。根據細胞存活率結果確定甾醇作用濃度。
1.3.3 細胞內TG相對含量測定
將對數生長期HepG2細胞以2×105個/孔接種于6 孔板中,按照分組處理各組細胞。吸去細胞上清液,磷酸鹽緩沖液(phosphate buffered saline,PBS)清洗3 次,加入100 μL細胞裂解液,輕晃使裂解液充分作用,將細胞收集于離心管中,2 000 r/min、4 ℃離心5 min,取上清液。按照試劑盒說明書測定細胞內TG含量。模型組、陽性對照組和甾醇類化合物處理組TG相對含量均以其相對空白對照組的含量表示,單位為%。
1.3.4 細胞內TC相對含量測定
細胞按照分組處理完成后,4 ℃、1 000 r/min離心10 min收集細胞。加入100 μL細胞裂解液,振蕩渦旋30 s,靜置5 min,反復處理6 次,離心后收集上清液,采用總蛋白定量檢測試劑盒測定上清蛋白質量濃度。取細胞上清于EP管中,加入等體積質量分數15% KOH-乙醇溶液,渦旋5 min,40 ℃水浴超聲皂化1 h,加入數滴質量分數6%三氯乙酸溶液去除蛋白,渦旋5 min,加入等體積正己烷-異丙醇(3∶2,V/V)充分混勻,室溫12 500 r/min離心15 min,收集上層有機相,剩余溶液反復抽提4 次。合并有機相,氮吹至近干,加入1 mL流動相復溶,室溫12 500 r/min離心15 min,取上清液,濾膜過濾后進樣,進行超高效液相色譜分析。
檢測條件:ZORBAX SB-C18色譜柱(4.6 mm×150.0 mm,5 μm),流動相為乙腈-異丙醇(67∶33,V/V),檢測器為二極管陣列檢測器,檢測波長206 nm、柱溫20 ℃、進樣量20 μL、流速1 mL/min,等度洗脫。模型組、陽性對照組和甾醇類化合物處理組的TC相對含量均以其相對空白對照組的含量表示,單位為%。
1.3.5 油紅O和Filipin III染色并觀察細胞結構
將對數生長期細胞以2.5×105個/孔接種于6 孔板,按前述分組處理48 h。根據油紅O染色液(細胞專用)試劑盒說明書對細胞染色,顯微鏡獲取圖像。
將對數生長期細胞接種在13 mm蓋玻片上,用膽固醇合成抑制劑辛伐他汀處理24 h,質量分數4%多聚甲醛溶液室溫固定30 min,PBS沖洗后,細胞用50 μg/mL的Filipin III避光染色45 min,PBS漂洗,并用顯微鏡觀察拍照。
1.3.6 蛋白免疫印跡法檢測HepG2細胞膽固醇調控相關蛋白的表達
細胞按照上述分組分別處理后,用預冷的PBS洗3 遍,收集各組細胞,每管加入150 μL細胞裂解液,冰上裂解5 min。隨后4 ℃、12 000 r/min離心10 min,取上清液,部分用于蛋白定量,剩余上清液加入Loading Buffer變性后保存于-80 ℃備用。蛋白樣品經十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳后轉移至聚偏二氟乙烯膜,5%牛血清白蛋白封閉1 h,加入相應抗體,4 ℃過夜,用TBST洗膜3 次,每次15 min,然后加入相應的辣根過氧化物酶標記二抗,室溫孵育1 h。用TBST洗膜3 次后在聚偏二氟乙烯膜上均勻加入ECL化學發光試劑,使用凝膠成像儀顯影。以β-actin條帶灰度作為參照,計算靶蛋白的相對表達水平。
1.4 數據處理與分析
所有實驗均重復3 次,結果用平均值±標準偏差表示。使用Origin 2021軟件進行圖形繪制,使用SPSS Statistics 21軟件進行方差分析,P<0.05表示差異顯著。
2 結果與分析
2.1 甾醇類化合物對HepG2細胞存活率的影響
為確定甾醇類化合物是否對細胞活力和增殖有影響,使用CCK8試劑盒檢測不同濃度甾醇類化合物對HepG2細胞的毒性作用。如圖1A所示,不同濃度各甾醇類化合物處理組與空白對照組的細胞存活率均無顯著差異,說明甾醇化合物的加入對細胞活力沒有影響,所選擇的濃度可用于后續實驗。

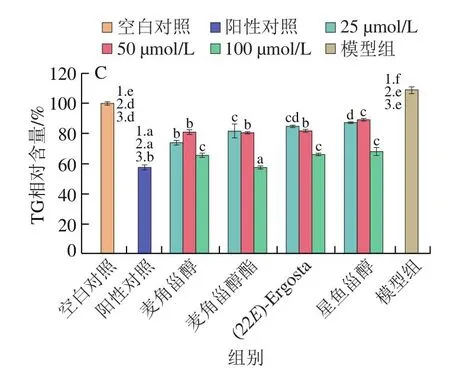
圖1 甾醇類化合物對HepG2細胞存活率(A)、細胞內TC(B)和TG(C)水平的影響(n=3)Fig.1 Effect of sterol compounds on the viability (A) and intracellular TC (B) and TG (C) levels of HepG2 cells (n = 3)
2.2 甾醇類化合物對細胞TC水平的影響
如圖1B所示,膽固醇和25-羥膽固醇處理細胞后,模型組細胞內TC水平相較于空白對照組顯著升高(P<0.05),與模型組相比,甾醇類化合物處理組TC水平發生不同程度的下降。濃度為25 μmol/L時,麥角甾醇組TC水平最低,其次為(22E)-Ergosta和麥角甾醇酯組,星魚甾醇組TC水平最高;濃度為50 μmol/L時,麥角甾醇、麥角甾醇酯和(22E)-Ergosta組之間無顯著差異(P>0.05),TC水平均顯著低于星魚甾醇組;濃度為100 μmol/L時,麥角甾醇酯和(22E)-Ergosta組TC水平較低,其次為麥角甾醇組,星魚甾醇組TC水平最高。
2.3 甾醇類化合物對細胞TG水平的影響
TG是重要的脂質組分,如圖1C所示,與模型組相比,4 種甾醇均顯著降低了高TC細胞內TG水平(P<0.05)。高濃度(100 μmol/L)時麥角甾醇酯降TG效果最佳,與陽性對照組辛伐他汀相當,其余3 組間無顯著差異;低濃度(25 μmol/L)時麥角甾醇的降TG效果最好;濃度為50 μmol/L時,麥角甾醇、麥角甾醇酯和(22E)-Ergosta組間TG水平沒有顯著差異,降TG效果均優于星魚甾醇(P<0.05)。
2.4 甾醇類化合物對HepG2細胞脂質蓄積的影響
油紅O可以特異性使細胞脂質染紅,通過顏色分布和程度可以觀察細胞內脂質分布情況。圖2顯示,膽固醇和25-羥膽固醇處理細胞后顏色較正常組更紅,說明脂質含量明顯增加,陽性藥物和甾醇類化合物處理后的細胞紅色分布減少,說明它們不同程度減少了HepG2細胞內的脂質蓄積,并且甾醇類化合物對細胞內脂質的減少作用呈濃度依賴性,其中(22E)-Ergosta作用最為明顯。結果表明甾醇類化合物可以降低膽固醇和25-羥膽固醇誘導的細胞內脂質積聚。
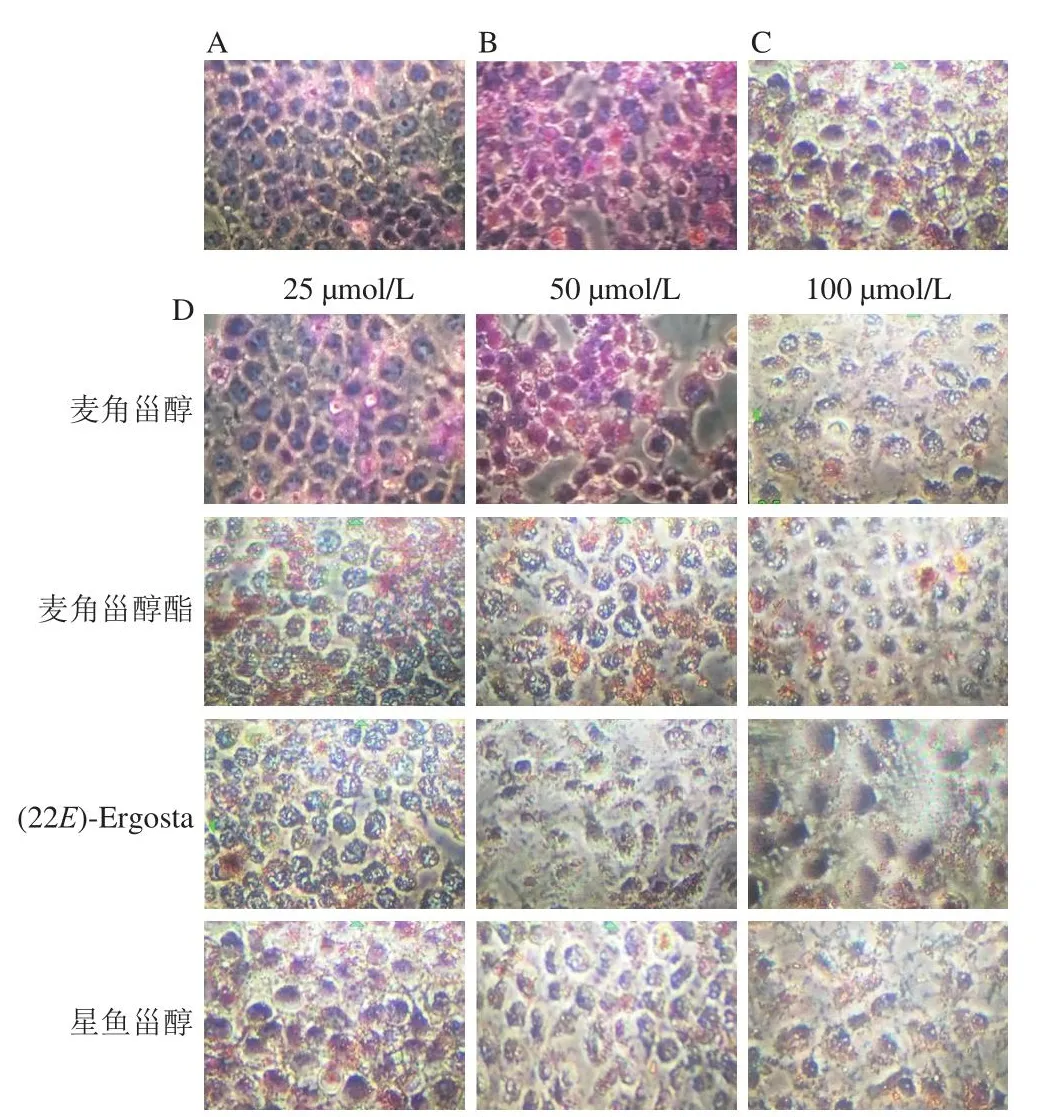
圖2 甾醇類化合物對HepG2細胞脂質蓄積的影響(×200)Fig.2 Effect of sterol compounds on lipid accumulation in HepG2 cells (× 200)
2.5 甾醇類化合物對HepG2細胞膽固醇代謝相關蛋白表達的影響
上述結果表明甾醇類化合物可以降低膽固醇和25-羥膽固醇誘導HepG2細胞內TC、TG水平并抑制脂質積聚。為進一步探究甾醇類化合物降低細胞內膽固醇的作用機制,分析膽固醇代謝相關蛋白表達水平變化。如圖3、4所示,相較于模型組,4 種甾醇類化合物均抑制了NPC1L1的表達,且抑制作用呈現劑量依賴性。其中高濃度麥角甾醇酯下調NPC1L1表達的效果與陽性對照辛伐他汀相近。與模型組相比,甾醇類化合物不同程度上調了ABCG5/8的表達,其中麥角甾醇的上調作用最為顯著,這與陽性對照辛伐他汀的作用效果相反。隨著作用濃度的上升,麥角甾醇酯、(22E)-Ergosta及星魚甾醇上調ABCG8蛋白表達的能力增強。HMGCS1和SREBP2是調控膽固醇合成的關鍵蛋白[10-11],ACAT2可將膽固醇酯化,隨后組裝成乳糜微粒進入淋巴循環[12],本實驗發現4 種甾醇類化合物均可有效下調HMGCS1、SREBP2和ACAT2蛋白的表達。

圖3 甾醇類化合物作用下HepG2細胞膽固醇代謝相關蛋白表達免疫印跡圖Fig.3 Western blot analysis of cholesterol metabolism-related protein expression in HepG2 cells treated with sterols
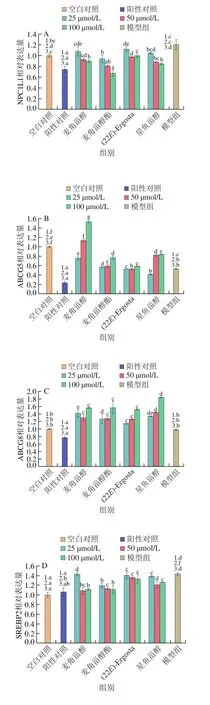
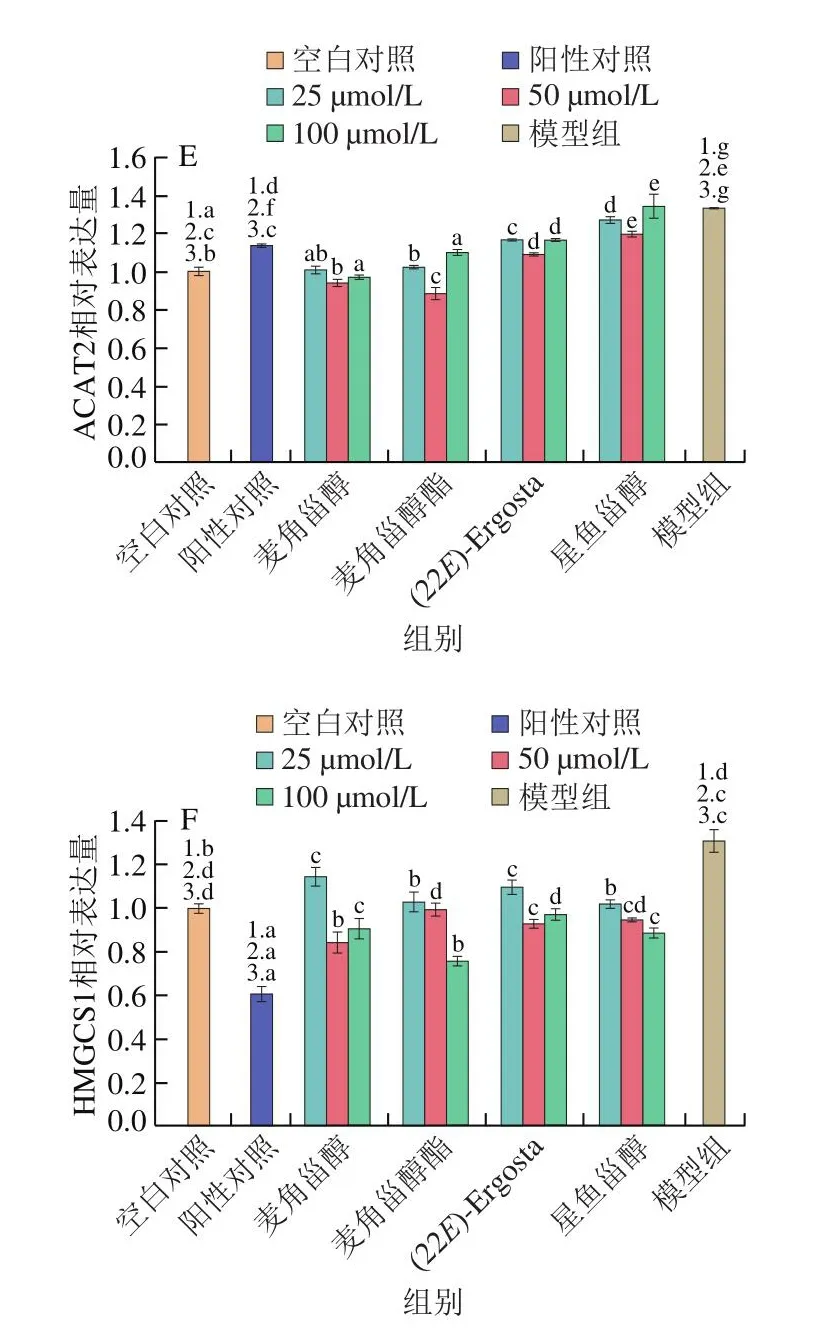
圖4 甾醇類化合物對HepG2細胞膽固醇代謝相關蛋白表達的影響(n=3)Fig.4 Effects of sterols on the expression of cholesterol metabolismrelated proteins in HepG2 cells (n = 3)
以上結果提示4 種甾醇類化合物可能通過降低NPC1L1、HMGCS1、SREBP2和ACAT2的表達,從而減少膽固醇的吸收、合成和酯化,進而降低細胞內TC水平。同時,甾醇類化合物上調了膽固醇轉運蛋白ABCG5/8的表達,說明甾醇類化合物促進了膽固醇的轉運。
3 討 論
甾醇的主要食物來源為植物油、面包、谷物和蔬菜等,隨著物質的豐富,不健康的飲食習慣使人們容易攝入過多的膽固醇,從而導致心血管疾病。現有研究證實甾醇具有顯著的降低膽固醇作用,補充甾醇類化合物可以改善血脂代謝情況,但是多數研究僅通過測定血清脂質、血清TC水平等指標[7,10,13-15]說明甾醇降膽固醇的作用,對于其降膽固醇的內在機制并未深入探索。本研究通過建立高膽固醇HepG2細胞模型,以辛伐他汀作為陽性對照,考察4 種甾醇類化合物(麥角甾醇、麥角甾醇酯、(22E)-Ergosta、星魚甾醇)對HepG2細胞TC、TG水平的影響,并通過其對膽固醇代謝相關蛋白表達的影響探究甾醇類化合物可能的降膽固醇機制。
TC、TG水平是反映機體脂質代謝水平的主要生化指標。研究發現甾醇類化合物可以有效降低TC、TG、LDL-C含量[16-18],在本實驗中,4 種甾醇類化合物降低了高固醇導致的細胞內TC及TG水平的升高,油紅O染色結果顯示甾醇類化合物改善了高膽固醇細胞的脂質蓄積。為進一步研究4 種甾醇類化合物對HepG2細胞內膽固醇水平調節作用的機制,使用Western blot測定膽固醇合成代謝相關蛋白的表達。人體內血漿膽固醇的穩態主要受到肝臟內源性合成、腸道對膳食膽固醇的吸收以及膽汁的清除和排泄控制。NPC1L1是膳食和膽汁膽固醇腸道吸收的關鍵蛋白[19]。NPC1L1在HepG2細胞中高表達,并定位于富含小GTPase Rab5蛋白的亞細胞泡狀室。Davies等[20]建立NPC1L1敲除的小鼠模型,與野生型小鼠的肝細胞相比,敲除NPC1L1的小鼠肝細胞表現出異常的質膜吸收和脂質(包括膽固醇和鞘脂)轉運。NPC1L1缺失可以完全抵消飲食誘導的小鼠高膽固醇血癥,其血漿脂蛋白和肝膽固醇譜與使用膽固醇吸收抑制劑依麥哲布治療的野生型小鼠相似[21]。本實驗發現4 種甾醇類化合物相較模型組顯著下調了NPC1L1蛋白的表達(P<0.05),表明甾醇類化合物可能通過減少膽固醇的吸收降低細胞內膽固醇含量。ACAT2在膽固醇酯化、腸道膽固醇吸收和載脂蛋白釋放中具有重要作用,研究表明植物甾醇可以下調ACAT2表達,從而有效抑制膽固醇吸收,降低脂肪水平[22-23]。在本研究中,模型組ACAT2表達量較空白對照組顯著升高(P<0.05),膽固醇可以激活ACAT2的表達,促進膽固醇酯化,4 種甾醇類化合物可以下調ACAT2表達,從而減少膽固醇的酯化,影響膽固醇的吸收。
ABCG5和ABCG8位于腸上皮細胞和肝細胞的頂膜上,ABCG5和ABCG8形成專性異源二聚體,限制腸道吸收膽固醇,促進膽固醇排泄到膽汁[24-25]。小鼠中過表達ABCG5和ABCG8可減少肝臟膽固醇循環從而延緩飲食誘導的動脈粥樣硬化[24]。本實驗結果表明4 種甾醇類化合物可以有效上調ABCG5/8蛋白的表達,且對ABCG8的上調較ABCG5顯著,說明4 種甾醇類化合物可能通過促進肝細胞內膽固醇向膽汁分泌,從而降低細胞內膽固醇水平。
HMGCS包括HMGCS1和HMGCS2,是促進膽固醇合成的重要酶[26],Yao Weilong等[27]研究表明抑制HMGCS1蛋白表達后細胞內膽固醇水平顯著下降。SREBPs能夠調控脊椎動物細胞中的脂質穩態,其激活后調節下游基因的表達[28],SREBP途徑的發現使控制膽固醇合成和攝取分子機制的研究取得重大進展[29-31]。本實驗中4 種甾醇類化合物顯著降低了HMGCS1及SREBP2蛋白的表達,說明4 種甾醇類化合物可能通過減少膽固醇的合成,從而降低膽固醇的水平。
4 結 論
通過添加膽固醇和25-羥基膽固醇構建高膽固醇HepG2細胞模型,在培養體系中添加不同濃度的4 種甾醇類化合物,對細胞進行油紅O染色,測定細胞中的TC、TG水平,發現4 種甾醇類化合物可以抑制膽固醇和25-羥基膽固醇引起的HepG2細胞內TC、TG水平升高,改善細胞內脂質積累情況。為進一步探究甾醇類化合物在高膽固醇HepG2細胞模型中的作用機制,對涉及膽固醇合成、轉運和代謝的相關蛋白表達水平進行測定發現,4 種甾醇類化合物可以降低NPC1L1、HMGCS1、SREBP2和ACAT2的表達,上調膽固醇轉運蛋白ABCG5和ABCG8表達。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
