褐藻膠多糖分子修飾酶的研究進展
2023-03-09 04:27:10寧利敏朱本偉
食品與生物技術學報 2023年1期
關鍵詞:結構
李 麗,寧利敏,姚 忠,朱本偉*,徐 虹
(1.南京工業大學 食品與輕工學院,江蘇 南京 210009;2.南京中醫藥大學 醫學院·整合醫學學院,江蘇 南京 210023)
褐藻膠是一種由β-D-甘露糖醛酸(β-Dmannuronate,M)和α-L-古洛糖醛酸(α-Lguluronate,G)通過1→4 糖苷鍵連接的線性多聚體[1],根據其分子的長度、M/G 殘基比率和分布以及乙酰化程度等,使其具有凝膠強度、水合能力、黏度和生物活性等特性,在食品、醫藥和化妝品等領域有著巨大的應用價值。褐藻膠的這些特性主要通過褐藻膠修飾酶對褐藻膠的合成和修飾過程進行控制,為了能夠利用不同的褐藻膠修飾酶進行高效、安全和綠色地制備具有不同性質的褐藻膠分子,作者旨在介紹褐藻膠修飾酶合成和修飾褐藻膠的作用機理,總結目前幾種褐藻膠修飾酶的來源、分類、結構、作用方式和研究進展,為將來更好地應用褐藻膠修飾酶和開發褐藻膠的商業價值提供參考。
1 簡 介
褐藻膠根據殘基的排列組合不同分成3 種片段:聚甘露糖醛酸(poly-mannuronate,poly-M)片段、聚古洛糖醛酸(poly-guluronate,poly-G)片段和甘露糖醛酸-古洛糖醛酸(poly-MG)雜合段。poly-M 和poly-G 的鏈式結構非常相似,單糖組分的區別僅僅是C5 上羥基位置的不同(見圖1)[1]。

圖1 不同結構褐藻膠的結構示意圖Fig.1 Structural diagram of alginate with different structures
自然界中的褐藻膠主要是由褐藻、一些鈣質紅藻、假單胞菌屬和固氮菌屬的一些細菌產生的。目前的商業褐藻膠主要是從一些褐藻植物中提取,如泡葉藻(Ascophyllum)、公牛藻(Durvillaea)、昆 布(Ecklonia)、螺紋雷松藻(Lessonia trabeculata)、巨藻(Macrocystis)和馬尾藻(Sargassum)等[2]。不同來源褐藻膠的組成和結構不同,例如螺紋雷松藻和極北海帶(Laminaria hyperborea)中的褐藻膠古洛糖醛酸含量較高(M/G 殘基比率<1),海洋巨藻(Durvillaea potatorum)中的褐藻膠古洛糖醛酸含量較低,而細菌中的褐藻膠還會發生乙酰化反應[3]。此外一些褐藻中的褐藻膠結構還受到季節、生長環境、年齡及不同部位的影響[4]。
褐藻膠開始由GDP-甘露糖醛酸合成,然后被甘露聚糖C5 差向異構酶、褐藻膠裂解酶、褐藻膠乙酰化酶和褐藻膠脫乙酰化酶在聚合物水平上進行修飾改性(如圖2)。褐藻膠是以聚甘露糖的形式合成的(見圖2 第一行分子);甘露糖醛酸殘基(M)可以在O2 或O3 位置通過甘露糖乙酰化酶(A)乙酰化、甘露聚糖C5 差向異構酶(E)而異構化為古洛糖醛酸殘基(G)或被褐藻膠裂解酶(L)降解產生不飽和的4-脫氧-L-赤式-6-4-烯吡喃糖醛酸酯殘基(Δ);乙酰基可以通過褐藻膠脫乙酰酶(D)去除。
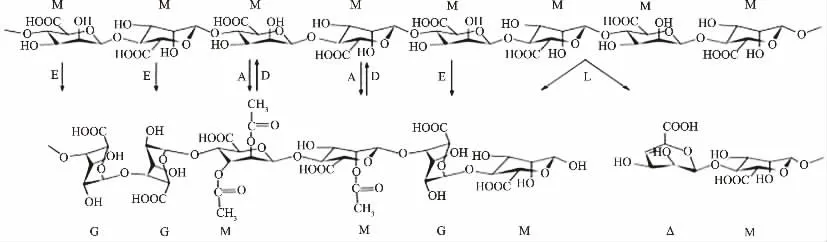
圖2 褐藻膠修飾酶的作用示意圖Fig.2 Schematic diagram of function of alginate-modifying enzymes
銅綠假單胞菌中合成褐藻膠所需的12 種蛋白質 由AlgD 操縱子編碼(AlgD、Alg8、Alg44、AlgK、AlgE、AlgG、AlgX、AlgL、AlgI、AlgJ、AlgF、AlgA),而另一種蛋白質AlgC 則在其他地方進行編碼[5]。褐藻膠合成復合體的模型如圖3 所示。首先,細胞質蛋白質AlgA、AlgC 和AlgD 通過一系列反應將D-果糖-6-磷酸轉化為GDP-甘露糖醛酸,這是合成褐藻膠的活化前體[6]。其次,GDP-甘露糖醛酸通過聚合成聚甘露糖醛酸鏈,部分Alg44[7]也參與了這個過程。Alg8 具有多個跨膜結構域和一個細胞質糖基轉移酶結構域,而Alg44 具有單個跨膜結構域,能將細胞質c-di-GMP 結合的PilZ 結構域與一個C 端周質結構域分開[8-9]。然后,在聚合時新生的聚甘露糖醛酸鏈被由AlgG、AlgL、AlgK、AlgX 和AlgE 組成的周質支架在周圍進行置換[9]。并且在周質中新生褐藻膠鏈的甘露糖醛酸殘基可以通過AlgI、AlgJ 和AlgF 的作用選擇性地在O2/O3 位置進行O-乙酰化[5]。周質支架中的AlgG 是一個差向異構酶,能夠將β-D-甘露糖醛酸轉化為α-L-古洛糖醛酸[10]。AlgL是一種雙功能周質蛋白質,不僅具有裂解褐藻膠的功能[11],同時作為修復系統存在于周質中,以清除這些合成異常的褐藻膠分子[12],保證周質支架的功能完整性。AlgK 位于周質中,通過脂質部分附著在外膜上,是一個廣泛參與蛋白質相互作用的結構域[13]。蛋白質AlgX 也在周質中與AlgK 和MucD 相互作用參與褐藻膠生物合成的翻譯后調節。AlgE 在外膜中形成一個β 桶孔,合成的褐藻膠最后通過AlgE從細胞中分泌出來[6]。

圖3 銅綠假單胞菌褐藻膠合成復合體模型[14]Fig.3 Alginate synthesis complex model of Pseudomonas aeruginosa[14]
此外如果產生褐藻膠的復合體不能正常工作的話,周質中的聚陰離子褐藻膠會吸引陽離子,從而產生滲透脅迫導致細胞溶解。
一般來講,經過修飾后的褐藻膠存在結構上的差異主要涉及以下幾點:
1)褐藻膠分子中M 殘基和G 殘基之間的比例和分布模式發生變化 如圖2 所示,先合成的部分甘露糖醛酸殘基(M 殘基)通過甘露聚糖C5 差向異構酶被異構化為古洛糖醛酸殘基(G 殘基),從而產生3 種褐藻膠多糖分子片段(poly-G、poly-M、poly-MG)。而poly-G 和poly-MG 的長度和數量影響著聚合物的幾個重要的物理特性,比如溶液的黏度及凝膠的穩定性、滲透性和水解性[14]。通常G 殘基含量比較少的聚合物不能形成陽離子凝膠,但是可以形成酸性凝膠。富含M 殘基的褐藻膠表現出免疫刺激性,而poly-G 則不具有免疫刺激性,即褐藻膠中M 和G 的相對含量直接關系到免疫活性強弱,且分子結構中各個聚合體的連接方式和連接順序對免疫活性也有直接的影響[15]。
2)不同生物體產生的褐藻膠多糖分子有不同程度的乙酰化修飾 細菌中產生的褐藻膠通常會在β-D-甘露糖醛酸的O2/O3 位置上選擇性地發生乙酰化修飾[16],而在海藻中產生的褐藻膠沒有這種變化。乙酰基能抑制聚合物與陽離子相互作用,因此褐藻膠的乙酰化可能會提高褐藻膠的黏度,降低其水溶性,從而增強褐藻膠形成凝膠的能力。去除O-乙酰殘基則會導致多糖與二價陽離子結合能力增強,多糖溶解度提高。另外,乙酰化的褐藻膠還可能與透明質酸類似,具有保留水和營養物質的能力[17]。
3)即使來源相同的褐藻膠其聚合度也有不同[12]在生物體合成褐藻膠的過程中,可能由于物理或者褐藻膠裂解酶的作用使褐藻膠多糖降解為褐藻膠寡糖,從而改變聚合物的聚合度。聚合度也會影響褐藻膠的黏度和凝膠強度,可以通過SEC-Malls 等方法測定[18]。
褐藻膠結構的多樣性影響了褐藻膠多糖的物理化學性質并使其在不同行業中有廣泛的應用前景。
在食品行業中,褐藻膠可以作為食品添加劑起到穩定、增稠和乳化等作用[19]。同時褐藻膠作為一種可食用纖維對預防結腸癌、心血管疾病、肥胖癥、體內重金屬的積累具有輔助治療的功效[20]。
在生物醫學材料的應用方面,褐藻膠作為支架材料其柔韌性可以完全彌補組織缺陷,并且光滑的表面可以避免對損傷部位的二次傷害[21]。Li 等使用褐藻膠和阿拉伯樹膠與鈣離子介導,通過交聯設計獲得了最佳黏性水凝膠支架,用于介導和加速一種細胞膜修復中的重要蛋白質MG53 的釋放,以改善傷口的再上皮化過程和持續釋放用于治療慢性傷口[22]。褐藻膠還是制藥工業中常見的賦形劑。有研究者在西咪替丁固定組合片劑中添加褐藻膠用于改善食管反流[23]。
在醫藥領域中,褐藻膠具有抗過敏、調節免疫、抗氧化和抗炎等生物活性,能夠激活人單核細胞產生在抗腫瘤、抗菌、抗病毒中發揮重要作用的免疫因子[24],故可用于治療外傷、修復皮膚損傷,具有良好的止血作用、可靠的治療效果和較低感染率等特性。Jiang 等證明褐藻膠寡糖顯示出有效地抗氧化活性和抗凋亡活性[25]。
2 褐藻膠裂解酶
褐藻膠裂解酶能夠將褐藻膠多糖降解為具有更高生物活性的褐藻膠寡糖[26]。此外褐藻膠裂解酶還可以用于褐藻資源的生物煉制[27]及褐藻膠多糖精細結構的表征[28]等方面。因此挖掘表征具有不同特性的褐藻膠裂解酶,對褐藻資源的利用具有十分重要的意義。
2.1 褐藻膠裂解酶的來源
在自然界中,褐藻膠裂解酶由許多利用褐藻膠作為碳源的細菌、動植物和病毒產生,目前細菌來源的褐藻膠裂解酶研究最為廣泛,已報道的產褐藻膠裂解酶的微生物主要包括弧菌、假單胞菌、黃桿菌、固氮菌、交替假單胞桿菌、鞘氨醇單胞菌以及克雷伯氏菌等[29]。其次,來源于病毒、真菌以及動植物的褐藻膠裂解酶也有一定的報道,Suda 等于1999年在小球藻病毒中也發現了褐藻膠裂解酶的基因[30]。Inoue 等首次在褐藻植物中提取出了一種褐藻膠裂解酶,進一步拓寬了褐藻膠裂解酶的來源[31],近年來不斷有人從鹽藻、紅藻和海帶等海洋藻類中分離出褐藻膠裂解酶[32-33]。在海洋軟體動物和棘皮動物體內也可以分離出褐藻膠裂解酶,如黑斑海兔(Aplysia kurodai)[34]、皺紋盤 鮑(Haliotis discushannai)和短濱螺(Littorina brevicula)等[35-36]。
2.2 褐藻膠裂解酶的分類
目前,褐藻膠裂解酶的分類方式主要有3 種:
1)根據氨基酸序列相似性多糖裂解酶被分成不同的家族,在CAZy 數據庫中,褐藻膠裂解酶可歸屬 到14 個多糖 裂解酶家族(PL):PL5、PL6、PL7、PL8、PL14、PL15、PL17、PL18、PL31、PL32、PL34、PL36、PL39 和PL41 家族[31],還有一些并不屬于以上家族的新型褐藻膠裂解酶,如Paenibacillus sp.LJ-23 中的Algpt[37]。其中來自細菌的褐藻膠裂解酶大多數屬于PL5、PL6、PL7、PL15 和PL17 家族,而來自真核細胞和病毒中的褐藻膠裂解酶主要屬于PL14 家族,不同的家族其在結構上有明顯的差異。
2)如圖1 所示,褐藻膠分子中存在4 種不同的化學鍵:M—M、G—G、M—G、G—M,大多數裂解酶對不同的化學鍵表現出不同的反應速率,根據對不同糖苷鍵的底物特異性,褐藻膠裂解酶可以分為:專一裂解聚甘露糖醛酸的裂解酶(EC4.2.2.3)、專一裂解聚古洛糖醛酸的裂解酶(EC4.2.2.11)以及對poly-M 和poly-G 均表現出高活性,可以更有效地降解褐藻膠的雙功能裂解酶(EC4.2.2.-)[38],并且不是所有的褐藻膠裂解酶都能裂解乙酰化的褐藻膠[39]。來自軟體動物的褐藻膠裂解酶和來自幾種海洋細菌的褐藻膠裂解酶多具有poly-M 底物特異性,那些發現于細菌培養液的胞外酶則大部分具有poly-G底物特異性。值得注意的是,某些poly-M 裂解酶/poly-G 裂解酶并不是只對poly-M/poly-G 有活性,也可以裂解poly-G/poly-M 和poly-MG,只是活性較低。雙功能裂解酶多集中在PL7、PL15 和PL17 家族中,許多雙功能裂解酶對不同的糖苷鍵活性不同,例如來自Persicobacter sp.CCB-QB2 的雙功能裂解酶AlyQ 對poly-G 和poly-M 的活性僅是對褐藻膠活性的60.7%和29.0%,表明AlyQ 更偏好裂解GM 或MG 之間的糖苷鍵[40]。而來自Wenyingzhuangia fucanilytica 的雙功能裂解酶Aly7B_Wf 對poly-M和poly-G 的活性分別為(27.03±1.69)U/mg 和(21.48±1.10)U/mg,表明該酶的首選底物為poly-M[41]。另外來自Sphingomonas sp.ZH0 的ZH0-I 對poly-G的活性則是對褐藻膠和poly-M 活性的120%[42]。
3)從作用模式來看,褐藻膠裂解酶可以分為3類:(1)內切型褐藻膠裂解酶,目前報道的大多數的裂解酶均具有內切酶活性,這些酶在褐藻膠內部隨機切割糖鏈上的1→4 糖苷鍵,并產生不同聚合度(Dps)的不飽和褐藻膠寡糖[43];(2)外切型褐藻膠裂解酶,一些外切酶可以從多糖的末端切除單體或二聚體寡糖產生不飽和單糖,然后通過非酶反應轉化成DEH(4-deoxy-l-erythro-5-hexoseulose uronate),如來自Zobellia galactanivorans DsijT 的AlyA3[44],目前報道的大多數外切酶屬于PL7、PL15 和PL17 家族;(3)內切和外切型褐藻膠裂解酶,有一些裂解酶同時具有內切和外切兩種活性,例如Huang 等分離純化出的Alg17B 裂解酶對褐藻膠表現出內切和外切兩種活性,產生單糖和低相對分子質量寡聚糖[45]。目前只有PL6、PL7 和PL17 家族有過這種類型裂解酶的報道[38,46-49]。表1 中列舉了已經表征的各個家族的典型褐藻膠裂解酶。

表1 典型的褐藻膠裂解酶Table 1 Typical alginate lyase
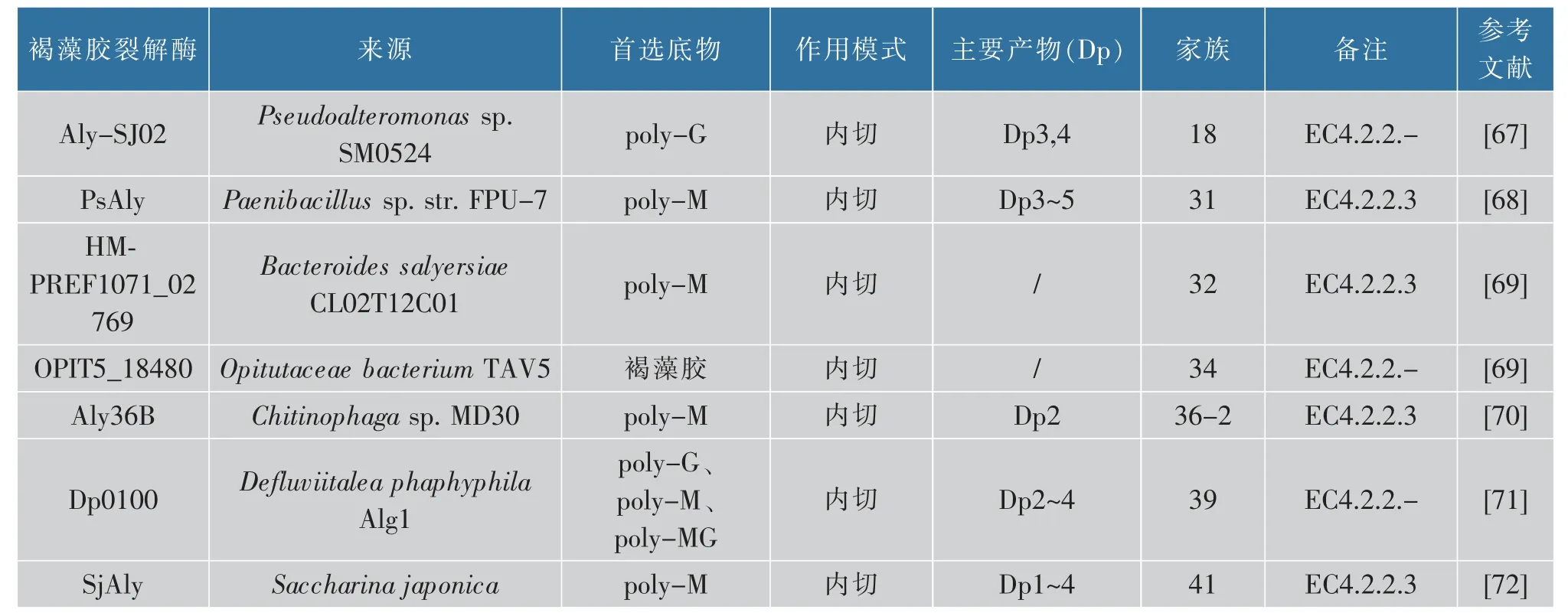
續表1
2.3 褐藻膠裂解酶的結構
作者所在課題組在褐藻膠裂解酶研究方面發表了多篇綜述,詳述了褐藻膠裂解酶的結構并闡釋了其與底物特異性之間的關系[29,73-74]。褐藻膠裂解酶的結構可以分成4 種類型[39]:
1)β-膠凍卷結構 這是褐藻膠裂解酶中最常見也是研究得最透徹的一種結構,PL7、PL14、PL18、PL36 家族的褐藻膠裂解酶多采用這種結構(見圖4)[75]。該結構可分為兩個相互連接的弧形反平行β 片:內凹片(SA)和外凹片(SB)。它們在中間進一步彎曲,形成近90°的球狀。內凹的薄片形成一個含有催化部位的裂隙并結合底物,在催化反應中起著不可或缺的作用。
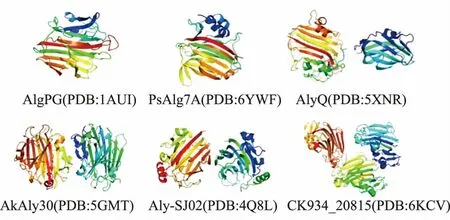
圖4 β-膠凍卷結構示意圖Fig.4 Schematic representation of β-jelly roll structure
2)β-螺旋結構 PL6 和PL31 家族的褐藻膠裂解酶多采用這種結構(見圖5),它由3 個β-折疊和3 個轉角(T)組成[76]。轉角(T)分別位于兩個β-折疊之間,一起形成了一個完整的β-螺旋(PB1-T1-PB2-T2-PB3-T3)。利用X 射線衍射儀等結構生物學技術對幾種具有β-螺旋結構的褐藻膠裂解酶進行了研究。褐藻膠裂解酶BcelPL6 在溶液中是一種單體,它由兩個結構域組成,N 末端結構域(NTD)和C 末端結構域(CTD)都采用右手平行的β-螺旋折疊。多肽的C 端(殘基432~468)為兩個平行于β-折疊PB3 平面的3 圈α-螺旋。β-螺旋的N 端部分在PB1 平面上有一個α-螺旋,C 端部分有一個α-螺旋和一個幾乎垂直于PB3 的β 鏈。生化分析表明,底物結合親和力主要由NTD 貢獻,而BcelPL6 的CTD 參與將底物固定到合適的構象。然而,CTD 具有較弱的褐藻膠裂解酶活性,可與PL6 結構域協同起到更有效的催化作用。此外,CTD 參與形成一個封閉的催化口袋,它的缺失使BcelPL6 對高度聚合底物的活性增加[77-78]。
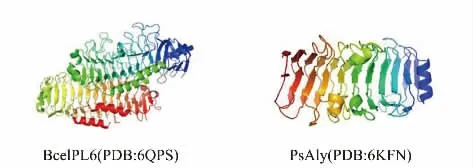
圖5 β-螺旋結構示意圖Fig.5 Schematic representation of β-helix structure
3)(α/α)n桶狀結構 PL5 家族褐藻膠裂解酶多采用這種結構(見圖6)。這種三維結構是由幾個反平行的α-螺旋構成的桶狀結構,從其頂部看螺旋環是逆時針方向的。例如,來自PL5 的A1-III 的活性中心由12 個α-螺旋形成,這些螺旋形成一個具有深隧道狀裂隙的α6/α5桶折疊。在這種模式下,底物穿透到隧道狀裂隙中,并進一步與催化位置相互作用[52]。

圖6 (α/α)n 桶狀結構示意圖Fig.6 Schematic representation of(α/α)n barrel structure
4)(α/α)n桶+反平行β-折疊結構的組合結構PL15、PL17 以及PL39 家族褐藻膠裂解酶多采用這種結構(見圖7)。PL15 家族中的Atu3025 被鑒定為該結構,其次還有一個口袋狀結構,口袋由兩個結構決定因素構成,即中心結構域和C 端結構域之間界面的構象變化和短α-螺旋H3 的存在,合理推測口袋狀結構是PL15 家族褐藻膠外切裂解酶的特征,對于釋放不飽和單糖作為唯一產物并識別底物非還原末端的外切模式至關重要[79]。PL39 中的Dp0100 褐藻膠裂解酶整體由3 個結構域組成,N 端結構域主要是螺旋結構,由不完整的(α/α)6環形成。中心結構域由16 條反平行的β 鏈排列在兩個β-折疊中,并帶有扭曲的α-螺旋。C 端結構域由兩個反平行β-折疊組成,形成一個典型的由16 條β 鏈組成的β-三明治。中心結構域的一面緊貼C 端結構域,形成一個4 層β-折疊層,另一面緊貼N 端結構域[80]。

圖7 (α/α)n 桶+反平行β-折疊結構結構示意圖Fig.7 Schematic representation of(α/α)n barrel +antiparallel β-fold structure
而PL8、PL32、PL34 以及PL41 家族褐藻膠裂解酶的結構尚未得到解析。
2.4 褐藻膠裂解酶的作用機制
褐藻膠裂解酶通過β-消除反應裂解褐藻膠,在C4~C5 位形成不飽和雙鍵,并在新的非還原末端存在一個4-脫氧-L-赤式-6-4-烯吡喃糖醛酸酯殘基,這個雙鍵在235 nm 處有吸光度,可以用來定量測定褐藻膠裂解酶的活性[81]。
Gacesa 提出整個β-消除反應通過三步反應去完成[82]:首先,帶正電的氨基酸殘基或者金屬離子中和C5 位羧基上的負電荷從而降低H-5 質子的pKa;其次,催化堿從C5 位提取一個質子從而形成烯醇中間體,羧基上的電子轉移裂解4-O-糖苷鍵,導致在C4~C5 位形成雙鍵并最終使得糖苷鍵的斷裂;最后,在糖苷鍵斷裂的同時產生了含有4-脫氧-L-赤式-6-4-烯吡喃糖醛酸酯殘基作為不飽和非還原末端的寡糖。該反應過程中需要催化堿去提取質子,并且需要催化酸去提供質子。當C5 位提取質子的方向與C4 位橋接氧在糖醛酸環的方向在同側時為順式消除(從M 殘基處進行消除),當兩者方向相反時為反式消除(從G 殘基處進行消除)。
根據催化位點以及中和底物負電荷的方式不同可將絕大多數褐藻裂解酶的消除反應分為六大類:
1)金屬離子輔助催化 PL6 家族使用鈣離子去中和C5 位羧基上的負電荷,然后Lys 充當催化堿從C5 位提取一個質子,Tyr 充當催化酸提供一個質子[83]。
2)His/Tyr 型β-消除 PL7 和PL15 家族中的褐藻膠裂解酶采用Tyr 以及His 分別充當催化酸和催化堿[84-85]。
3)Tyr/Tyr 型β-消除 在催化過程中,兩個Tyr分別充當催化酸和催化堿,例如PL17 家族中的Alg17c,該酶分別采用Tyr450 殘基充當催化堿和Tyr258 作為催化酸[86]。
4)Tyr/Tyr 型β-消除 PL5、PL14 和PL18 家族中的褐藻膠裂解酶在催化過程中,同一個Tyr 既充當催化酸又充當催化堿[87-89]。
5)水催化型β-消除 在催化過程中水分子用于中和羧基上的負電荷,Arg 以及Lys 分別充當催化酸和催化堿。
6)Lys/Lys 型β-消除 在催化過程中同一個Lys 既充當催化酸又充當催化堿,例如PL36 家族中的Aly36B 在催化過程中Arg169、Tyr185 和Tyr187負責中和底物的負電荷,Lys143 既充當催化酸又充當催化堿[70]。
2.5 褐藻膠裂解酶的酶學特性
目前針對褐藻膠裂解酶酶學特性的研究報道較少,而某些酶學性質如嗜熱、嗜冷、溫度穩定性、pH 穩定性和鹽活化等的研究有利于酶適應生存環境和應用于工業化生產。因此,作者對已報道的典型褐藻膠裂解酶的酶學性質進行了整理。
熱穩定性是褐藻膠裂解酶應用中最重要的性質之一,在褐藻膠裂解酶催化過程中,由于熱穩定性的存在,催化反應可以在更高的溫度下進行,反應混合物黏度的降低和酶活性的提高從而促進底物的轉化[90]。當催化海帶粉等粗基質發生轉化時,高工藝溫度(高于45 ℃)還可以防止微生物污染。因此具有良好熱穩定性的褐藻膠裂解酶對于褐藻膠低聚糖的酶促生產有重要價值。目前已知的大多數褐藻膠裂解酶不具有熱穩定性,只在30~40 ℃表現出最佳催化活性[91],而關于具有熱穩定性的褐藻膠裂解酶報道較少,表2 中列舉了目前報道的一些熱穩定性良好的褐藻膠裂解酶。
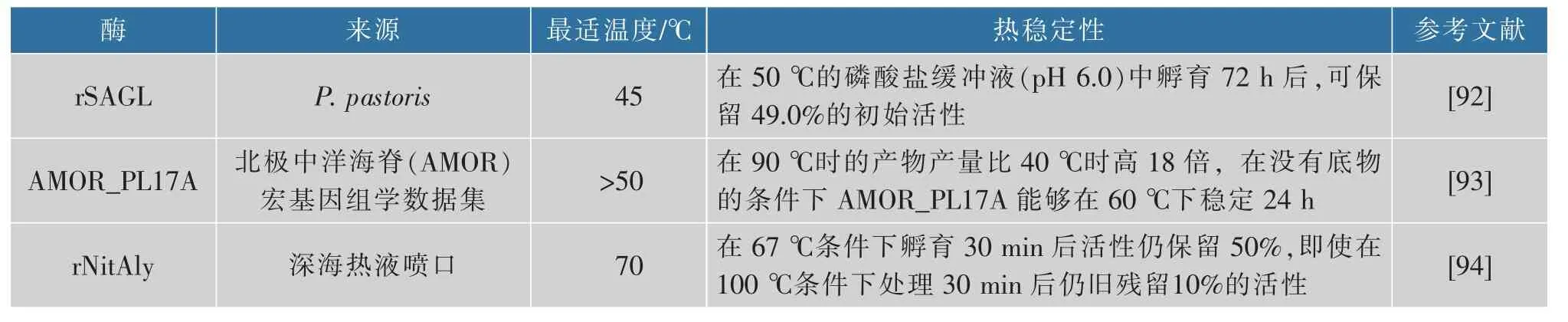
表2 典型的熱穩定性褐藻膠裂解酶Table 2 Typical thermostable alginate lyase
到目前為止有關褐藻膠裂解酶耐熱機制的相關研究還較少,根據已有報道,rNitAly 的熱穩定性與Cys80 和Cys232 之間形成的二硫鍵有關[94]。來自Cobetia sp.NAP1 的AlgC-PL7 的熱穩定性可能與α-螺旋有關[95]。Yang 等對AlyM 進行保守結構域重建,去除F5_F8_type_C 結構域后,在45 ℃下孵育1 h 的熱穩定性由保留30%~40%的酶活力提升到了保留大約70%的酶活力[96],經截斷后的突變體具有比全酶更緊湊的結構,能更好地抵抗熱變性的影響,證明酶的熱穩定性也可能與酶的結構緊湊程度有關。
在低溫條件下進行酶促反應可以減少能耗、降低微生物污染的風險和終止催化反應的難度,一般的耐冷性褐藻膠裂解酶在低于35 ℃時具有最高的催化活性,并通常在20 ℃時保留最高活性的50%[57]。表3 中列舉了3 種具有良好低溫穩定性的褐藻膠裂解酶。
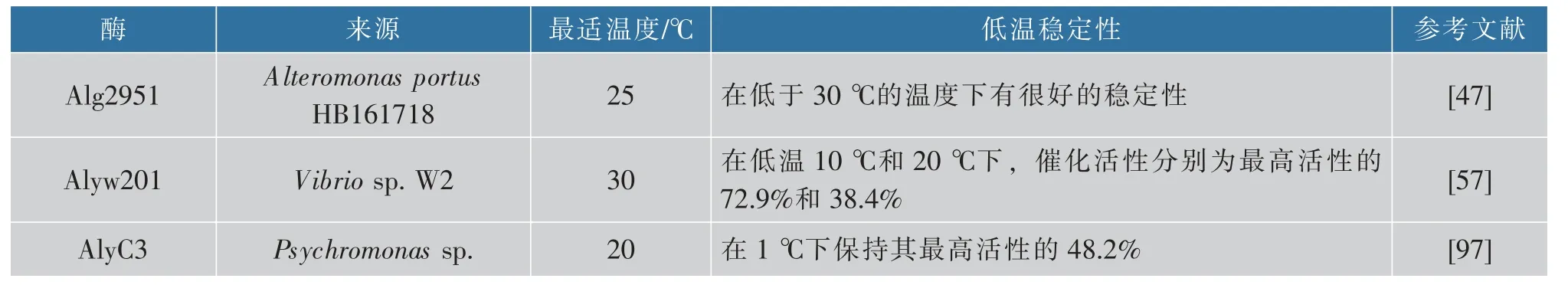
表3 典型的耐冷褐藻膠裂解酶Table 3 Typical cold-adapted alginate lyase
對于大多數褐藻膠裂解酶來說,催化的最適pH 接近中性,并且只在狹窄的pH 范圍內表現出較高活性,也有一些酶在狹窄的pH 堿性條件下表現出最高活性。只有少數酶在較寬的pH 范圍內表現出較高活性。例如來自Vibrio sp.NJ-04 的AlgNJ-04裂解酶在pH 4.0~10.0 的寬pH 范圍內保留了超過80%的活性,表現出優異的pH 穩定性[98]。而Alyw201 在pH 5.0~10.0 孵育12h后仍保留超過70%的活性。特別是在檢測到的整個pH范圍(pH 3.0~11.0)中,孵育12 h 后仍保留超過40%的活性[57]。另外,來自Pseudoalteromonas carrageenovora ASY5 的Alg823[99]、Vibrio sp.SY01 的Aly08[100]、Serratia marcescens NJ-07 的AlgNJ-07[101]、Paenibacillus sp.LJ-23 的Algpt[37]等均具有良好的pH 穩定性。
從海洋環境中分離出的褐藻膠裂解酶通常還具有鹽活化這一特性,即在一定濃度的氯化鈉溶液中,褐藻膠裂解酶的活性增加數倍,反映了它們對海水環境的適應能力。如在1 mol/L NaCl 條件下,AlgM4 的活性 增加了 約7 倍[102];AlyPM 其活性 在0.5~1.2 mol/L NaCl 條件下可以增加6 倍[103];而對于來自Vibrio sp.的A9mT 在0.4 mol/L NaCl 條件下其活性增加了24 倍[104]。盡管到目前為止已經發現了很多褐藻膠裂解酶可以被鹽激活,但是其中的激活機制并沒有被徹底揭示出來。根據已有報道,AlgNJ-04 的鹽活化特性是由于褐藻膠分子中結合水的去除或褐藻膠-酶復合物形成過程中的電荷效應[98]。AlgM4 的二級結構中α-螺旋和β-折疊的含量被NaCl 改變從而增強了其對底物的親和力和抵抗熱變性的能力[102]。AlyPM 的鹽活化特性也是由于在NaCl 存在時其對底物親和力增強,但是結構并不發生變化[103]。AlyC3 的鹽活化機制則是因為保留了二聚體四元結構[97]。
總之,目前對褐藻膠裂解酶的基礎研究依然主要集中在新型褐藻膠裂解酶的表征、酶學性質分析以及對酶的三級結構分析,而為了更好地利用褐藻膠裂解酶進行工業化生產,需要進行更多的研究來揭示褐藻膠裂解酶催化機制。
2.6 褐藻膠裂解酶的應用
褐藻膠裂解酶的應用包括褐藻膠寡糖的制備、原生質體分離、肺囊性纖維化病的治療以及褐藻膠測序等方面。
1)褐藻膠寡糖的制備 褐藻膠寡糖相對分子質量小,具有多種生物活性。褐藻膠寡糖的制備方式包括酸解、熱解和酶解等。Iwamoto 等比較了酸解和酶解制得的褐藻膠寡糖對腫瘤壞死因子TNF-α的誘導活性,結果證明酶解得到的褐藻膠寡糖具有更加廣泛的生物活性[105]。根據Belik 等報道通過褐藻膠裂解酶降解天然的聚甘露糖醛酸,產生的甘露糖醛酸寡糖可用于協同治療腫瘤[106]。
2)原生質體分離 褐藻膠是褐藻細胞壁中含量最豐富的成分,除此還有巖藻糖膠、纖維素、多酚和蛋白質等物質。研究表明多酚與褐藻膠網絡相連從而進一步加強細胞壁的強度[107]。采用纖維素酶和褐藻膠酶混合的方法可以降解褐藻細胞壁,釋放原生質體[108]。
3)肺囊性纖維化病的治療 褐藻膠裂解酶能夠配合一些抗生素降解肺囊性纖維化病人肺中病原菌產生的褐藻膠,使病原菌的細胞壁通透性增強,利于抗生素發揮抗生作用[109]。Patel 等開發了環丙沙星-褐藻膠裂解酶功能化殼聚糖納米顆粒(AgLase-CIPR-CH-NPs)用于有效治療肺囊性纖維化患者銅綠假單胞菌感染[110]。
4)褐藻膠測序 核磁共振是表征褐藻膠分子最常見的一種方法,然而其只能得出G 殘基和M殘基的統計分布,利用具有特殊底物特異性以及作用模式的酶來降解褐藻膠,再結合一些現代分析方法如質譜等對褐藻膠片段進行表征能夠進一步測量片段長度分布[28]。
5)生物燃料的制備 褐藻因其高生長速率和高糖分含量而被認為是生物燃料乙醇生產中的可再生生物質。由于缺乏木質素,褐藻生物質的糖化相對容易。但是傳統的工業微生物無法代謝褐藻膠,經代謝工程改造的微生物生產的外切型褐藻膠裂解酶可以有效利用褐藻膠,以此克服了該問題[111]。Wang 等優化了兩種重組褐藻膠裂解酶(內切型Alg7D 和外切型Alg17C)的酶促糖化過程,以便從褐藻膠中高效生產DEH,而DEH 是利用褐藻生物質生產生物燃料的關鍵底物[112]。
2.7 提高褐藻膠裂解酶的應用能力
目前針對褐藻膠裂解酶的應用能力,主要是通過酶的固定化和酶的分子改造提高酶活性或者改善某種特性,大大提高了酶的工業化應用效率。
為了發揮酶的最大應用價值,近年來研究人員證明了固定化酶在穩定性、可重復利用性和易于分離方面顯示出比游離酶更好的優勢,酶可以通過吸附、包埋和交聯進行固定化。目前對固體載體上固定褐藻膠裂解酶已經進行了一些研究,例如生物聚合物微球、超濾膜、殼聚糖納米顆粒、介孔氧化鈦顆粒等,為褐藻膠寡糖的生產、工業廢水的處理以及抗生物膜治療等方面提供了褐藻膠裂解酶應用的途徑[25,83]。作者所在課題組將褐藻膠裂解酶AlyPL6固定在介孔氧化鈦顆粒(MTOPs)上后,在重復使用10 次后,在45 ℃下保留>55.4%的活性[113]。Li 等將褐藻膠裂解酶Aly08 固定在低相對分子質量殼聚糖納米顆粒上,與游離的Aly08 相比,固定化的ALLMW-CS-NPs 在抑制銅綠假單胞菌生物膜的形成和阻斷成熟的生物膜方面表現出更高的效率,使得銅綠假單胞菌的生物量大大減少。該研究促進了褐藻膠裂解酶作為抗生物膜劑的進一步發展[114]。而Meshram 等將AlgL 固定在醋酸纖維素超濾膜上,成功地解決了多糖結垢問題對水凈化的影響[115]。
酶的分子改造主要包括合理設計、半理性設計、定向進化、保守結構域重組和非催化結構域截斷[116],經過分子改造后的突變體具有與初始酶不同的酶學特性。作者所在課題組系統總結了褐藻膠裂解酶分子改造的研究進展[117],目前常以引入二硫鍵、重建保守結構域以及定向進化來提高酶的活性和穩定性。例如,Yang 等在對cAlyM 的催化位點、二級結構以及空間構型進行綜合分析后,在分子中引入了二硫鍵,設計出了突變體D102C-A300C 和G103C-T113C,t1/2(45 ℃)分別增加了2.25、1.16 h[118]。作者所在課題組研究發現褐藻膠裂解酶Aly7B 在35 ℃下表現出較差的熱穩定性。然而,當截斷Aly7B-CDI(非催化結構域:R2~Y181)時,Aly7BCDII(催化結構域:W190~H477)在35 ℃下活性可以保持不變[119],熱穩定性大大提高。Xu 等通過在AlgL-CD 的活性中心引入堿性氨基酸來合理設計酶分子以增加酶活性,其中突變體E226K 的整體構象變得更加靈活,與底物的親和力增加,表現出比野生型AlgL-CD 更高的酶活力,但同樣由于鈣結合環的靈活性變高,與Ca2+的結合能力變弱,E226K 的熱穩定性變差[120]。而Su 等基于E226K 的作用模式,選擇loop 環1 上的I211 位點和底物入口處的E276、Y292 和R294 位點作為工程靶點,設計出的突變體E226K/I211T/R294V 催化裂隙周圍的環更靈活、底物入口更大,酶的催化效率提高了4.78 倍,半衰期t1/2(45 ℃)從89 min 增加到557 min[121]。此外,分子改造還可以用于改造酶的最終產物,Zhang等報道了借助AlyF 中間產物的裂解模式變化,對與糖結合的殘基Arg266 進行位點突變,改變亞位點對糖的親和力,結果證明F128T/W172R/R226H突變體的主要產物由三糖(三糖占比為87.0%)變為二糖和三糖(二糖占比提高到40.5%)[122]。
3 甘露聚糖C5 差向異構酶
甘露聚糖C5 差向異構酶(mannuronan C5-epimerase,MC5E)是一種褐藻膠修飾酶,催化褐藻膠中的β-D-甘露糖醛酸(M)轉化為C5 同分異構體α-L-古洛糖醛酸(G)。研究表明純化后的一些MC5Es 能在體外引入G 片段,并進一步增加褐藻膠中的G 殘基含量,經驗證AlgE2 在特定的褐藻膠中將G 殘基相對含量從初始值(0~45%)提高到約70%[123]。而AlgE6 和AcAlgE1 甚至可以將G 殘基相對含量分別提高到78%和87%[124]。考慮到褐藻膠的理化性質和生物活性與G/M 的含量有關,MC5E 作為一種褐藻膠修飾酶是生產具有特殊性質的褐藻膠的重要工具酶。
3.1 甘露聚糖C5 差向異構酶的來源及分類
MC5Es 有兩個主要的來源:真核生物來源,包括褐藻、海帶和昆布等;細菌來源,主要包括假單胞菌屬(如熒光假單胞菌、銅綠假單胞菌、丁香假單胞菌和門多薩假單胞菌)和固氮菌屬(如圓褐固氮菌和棕色固氮菌)[124]。不同物種中的MC5Es 具有不同的數量、類型和特征。
MC5Es 可分為Ca2+依賴亞型和Ca2+非依賴性亞型。例如丁香假單胞菌中的PsmE、圓褐固氮菌中的AcAlgE1 以及棕色固氮菌中AlgE1~7 的酶活性取決于Ca2+的存在,而丁香假單胞菌、棕色固氮菌、銅綠假單胞菌中的周質AlgG 可以在無Ca2+條件下高效地將甘露糖醛酸轉化為古洛糖醛酸[125]。對于依賴Ca2+的MC5Es 來說,Ca2+參與中和異構化反應中甘露糖醛酸的電荷,而在丁香假單胞菌中的Ca2+非依賴型周質AlgG 中一些特殊的氨基酸,如Arg345,也可以執行同樣的功能[126]。
MC5Es 也可以分為單功能和雙功能MC5Es。例如棕色固氮菌中的AlgE2 和AlgE7 都具有差向異構酶活性和裂解酶活性,在存在Ca2+(3.3 mmol/L)的情況下,AlgE2 可以修飾褐藻膠并使其鏈中的糖苷鍵發生斷裂,具體表現為催化后的褐藻膠平均相對分子質量下降和整個褐藻膠溶液的黏度下降[123]。同樣,AlgE7 也具有與AlgE2 相同的雙功能且其裂解酶活性高于AlgE2[127]。Gawin 等也在2020 年從棕色固氮菌中鑒定出3 種褐藻膠修飾酶,其中AcAlgE2和AcAlgE3 具有裂解酶和異構酶活性,且在體外條件下只顯示出裂解酶活性[128]。來自銅綠假單胞菌的AlgG 不僅具有異構酶活性,而且還能保護褐藻膠不被降解[129]。來自丁香假單胞菌中Ca2+依賴的PsmE會催化褐藻膠中的M 殘基的異構化和O-乙酰水解[125]。
3.2 甘露聚糖C5 差向異構酶的結構及催化機制
棕色固氮菌產生的7 種分泌性差向異構酶(AlgE1~7,AlgEs)結構特征已經研究清楚,AlgEs 由兩個不同的蛋白質模塊組成,包括一個或兩個A 模塊(約385 個氨基酸)和1~7 個R 模塊(約150 個氨基酸)[123]。每個AlgE 都有一個獨特的A 和R 模塊的序列、數量和分布(見圖8),每個胞外AlgE 的最后一個R 模塊通常都有一個參與酶分泌的非結構化多肽[130]。含有與褐藻膠和Ca2+結合位點的A 模塊可以進行異構化反應和確定最終的異構化反應模式[131]。而R 模塊不具有異構化活性,主要通過降低催化反應對Ca2+濃度的要求,使A 模塊的催化活性提高了約10 倍[132]。除此之外,具有9 個氨基酸序列的4~7 個重復的R 模塊參與了酶的分泌和Ca2+的結合[130]。丁香假單胞菌的PsmE 具有與AlgEs 相似的A 和R 模塊,同時還具有額外的M 區、N 區和RTX 區。M 區和RTX 區都是參與Ca2+的結合,而N區是一種乙酰化酶[125]。此外,圓褐固氮菌中的AcAlgE1、AcAlgE2 和AcAlgE3 具有與AlgEs 相 同的模塊結構[35]。
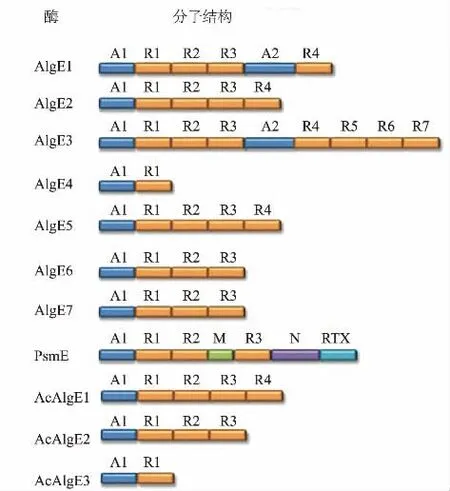
圖8 MC5Es 的模塊結構Fig.8 Module structure of MC5Es
研究人員通過核磁共振方法確定了AlgE4 的結構,AlgE4 的A 模塊折疊成一個右手平行β-螺旋,由4 個平行的β-折疊組成,包括12 個完整的匝數[133]。突變體實驗證明,Tyr149、Asp152、His154 和Asp178 是AlgE4 活性的關鍵殘基,甘露糖醛酸的質子化羧基可以與Asp152(或Asp178)形成氫鍵,Tyr149 作為催化堿提取與C5 結合的質子,質子化的His154 作為催化酸貢獻一個質子對C5 進行親核攻擊,最后形成古洛糖醛酸[133]。R 模塊形成了一個右手平行的β-膠凍卷。根據計算,A 模塊可以結合11 個糖醛酸殘基,R 模塊可以結合5 個殘基[134]。AlgE4 的結構模型可以用于更好地理解其他AlgE1~7 結構。
Wolfram 等解釋了來自丁香假單胞菌的AlgG的分子結構,它折疊成右手平行β-螺旋,有11 個完整的線圈和一個不完整的線圈,其中線圈4~10 形成一個碳水化合物結合域或糖類水解結構域[126]。
研究表明,褐藻膠裂解酶和甘露聚糖C5 差向異構酶的催化機制是相似的[133],裂解反應和異構化反應都涉及3 個步驟,兩者均需要中和羧基上的負電荷,并催化提取C5 位上的質子,兩者的反應機理差異發生在最后一步,對于裂解酶來說,最后一步是糖苷鍵的斷裂,同時在C4 和C5 之間形成雙鍵,離開的基團發生質子化形成一個新的還原端。而甘露聚糖C5 差向異構酶需要一個氨基酸殘基,該殘基能夠從吡喃糖環的另一側向C5 提供一個質子,形成C5 差向異構體。
AlgG 和AlgE4 的A 模塊的整體結構與一些β-螺旋結構的褐藻膠裂解酶(PL6、PL31 家族)在結構上有相似之處,然而使用Dail 進行蛋白質結構比較結果表明,它們之間沒有任何可能表明共同進化起源的序列同源性,并且這些褐藻膠裂解酶的催化殘基在AlgE4 中的A 模塊中并不保守[133]。異構酶AlgE4 中的A 模塊的催化位點更類似于褐藻膠裂解酶A1-III(PL5 家族)和ALY-1(PL7 家族),ALY-1和A1-III 的底物結合裂縫中心的4 個催化殘基(Gln/Asn、His、Arg 和Tyr)在反應機理中起著關鍵作用。AlgE4 中的A 模塊的活性位點Asp152、His154、Lys117 和Tyr149 也顯示出相同的空間排列,并且甘露糖醛酸三糖以與A1-III 中結合的三糖非常相似的方向和位置結合。盡管4 個活性位點氨基酸殘基的位置相似,但是其他重要殘基(尤其是Asp178)和亞位點-2 和-3 的環境非常不同。
AlgG 與AlgE4 的活性位點結構非常相似,活性位點的主要區別在于AlgE4 不包含與AlgG 中發現的Arg345 相當的殘基,而是包含參與形成AlgE4中Ca2+結合位點的幾個酸性殘基。正如在大多數β-螺旋褐藻膠裂解酶中所發現的那樣,這種Ca2+在反應過程中能夠中和糖醛酸的羧酸基團,從而發揮與Arg345 相當的作用[126]。依賴Ca2+的褐藻膠裂解酶和AlgE4 通過Ca2+中和糖醛酸上的負電荷,而不依賴Ca2+的褐藻膠裂解酶和AlgG 則使用一個精氨酸殘基來完成這個反應[135]。在第二步催化中,精氨酸或賴氨酸通常作為裂解酶的催化堿基,而去質子化的His319 能夠從甘露糖醛酸的C5 中提取質子,His319 很可能在AlgG 的催化反應中起催化堿基的作用。那么,水分子在AlgG 的催化過程中充當催化酸,這也與前文中提到的一些具有β-螺旋結構的褐藻膠裂解酶也有水分子作為催化酸的結論一致[126]。
對于雙功能酶AlgE7,Gaardlos 等解釋了兩種活性如何相互調節的結構基礎,首先異構酶和裂解酶活性具有相同的催化位點,這兩種活性通過不斷改變褐藻膠鏈而相互影響,并且產物的形成高度依賴于酶作用于哪種底物。當使用明確的底物poly-M和poly-MG 時,裂解酶優先的裂解位點為M↓XM和G↓XM(X 為G 或M)。其中H154 在反應中充當催化堿,Y149 充當催化酸,而突變體R148G 幾乎失去了所有裂解酶活性,但保留了異構酶活性,因此推測R148 會影響Y149 將質子提供給糖苷鍵或糖環的另一側[136]。
3.3 甘露聚糖C5 差向異構酶的作用方式
關于MC5E 作用方式的報道較少,相關報道主要集中在棕色固氮菌中的AlgE 家族,目前已經提出兩種反應模式,即優先攻擊模式和漸進作用模式[137]。在優先攻擊模式中,酶優先攻擊鄰近G 殘基中的M 殘基,并在每次差向異構化反應后脫離底物[138]。而在漸進作用模式中,由于甘露糖醛酸之間通過β-1,4 糖苷鍵連接,每個殘基相對于其相鄰殘基旋轉近180°,因此酶和多糖鏈相對移動,使得下一次異構化反應可以在酶與底物不解離的情況下完成[138]。有研究報道了AlgE4 的漸進作用模式,酶沿著底物鏈進行滑動,每隔一個殘基進行一次異構化,不需要發生旋轉。即底物封閉在一個相對較大的凹槽中,并且在活性位點的每一側都有一個夾子,該夾子封閉了聚合物并幫助其滑動[137]。而丁香假單胞菌中的AlgG 也遵循漸進作用模式,這也是為什么假單胞菌中的褐藻膠出現M 和G 交替的原因[126]。此外,AlgE2 和AcAlgE1 遵循優先攻擊模式進行反應,并且與底物的組成和鈣離子的濃度有關[126,128]。而其余的AlgEs 都是根據優先攻擊模式或者兩種機制的組合進行反應[132]。
由于反應模式的不同,不同的AlgEs 異構酶所產生的產物也不同。根據報道,AlgE1 具有兩種不同的催化結構域,一種產生poly-G,一種產生poly-MG[139],AlgE2 和AlgE5 主要產生短的poly-G[140],PsmE、AlgE6 和AcAlgE1 主要產生長的poly-G[140],AlgE4 主要產生poly-MG[141]。人們可以利用該產物特點獲得不同類型的褐藻膠低聚糖。
3.4 甘露聚糖C5 差向異構酶的酶學特性
目前已經表征的大多數異構酶的最適pH 在6.5~8.0,最適溫度范圍為30~40 ℃[124]。據報道,來自Pseudomonas mendocina sp.DICP-70 的PmC5A 作為裂解酶和異構酶時的最適pH 分別為8.0 和9.0,是目前最適pH 最高的酶[142]。金屬離子影響酶活力,當Na+濃度為100~200 mmol/L 時,AlgE1 活性最高[124]。對于AlgE2,Ca2+濃度為0.58 mmol/L 時,初始反應速率隨Na+濃度的增加而增加,當Ca2+濃度達到3.3 mmol/L 時,Na+對異構化反應沒有促進作用,而當Na+濃度達到約20 mmol/L 時,對異構化反應有抑制作用[143]。對于雙功能酶AlgE7,較高的Na+濃度可降低裂解酶活性并增加poly-G 的形成,而高Ca2+濃度則提高裂解酶活性并減少poly-G 的形成[136],因此,可以通過調節Na+與Ca2+濃度來調節AlgE7 的裂解酶活性與異構酶活性,獲得目標產物。研究人員還證明在某些情況下Sr2+可以代替Ca2+,但酶的活性明顯降低[144]。
3.5 甘露聚糖C5 差向異構酶的應用
MC5E 作為一種褐藻膠修飾酶,能夠增加底物的G 殘基含量,將褐藻膠或褐藻膠寡糖升級為具有長poly-G 或poly-MG 的產品,該類型的產品表現出優良的機械性能和獨特的生物活性,可以應用于制備藥物釋放材料、用作抗菌劑、促進角質細胞增殖等方面。
1)制備具有優異機械性能的材料 富含G 的褐藻膠分子可以通過與一些二價離子結合而形成熱穩定的三維凝膠網絡,形成的凝膠[145]具備一定的剛性和機械強度,具有低收縮、高孔隙率[146]、高穩定性和高交聯密度等特點。用MC5E 調節褐藻膠分子中的G 殘基含量從而制備具有優異機械性能的材料是一種較為簡單、可控和節能的方法。
2)制備應用于藥物釋放的褐藻膠 褐藻膠是輸送藥物的理想材料。藥物擴散過程的速度主要與褐藻膠分子的孔隙大小有關,研究表明,poly-G 含量較高的褐藻膠分子具有更開放的孔隙結構,表現出較高的擴散能力。而富含M 的褐藻膠分子是軟的,多孔性較差,并且隨著時間的推移容易脫膠[147]。因此,富含G 的褐藻膠能夠更好地保護藥物不被破壞,使其達到特定的位置并最大限度地發揮藥效。
3)增強褐藻膠寡糖的抗菌活性 富含G 的褐藻膠寡糖可以通過與細胞表面的結合來分解生物膜,影響表面電荷,導致微生物的聚集,并通過阻止微生物移動來影響致病微生物的活性并增強抗生素的效果,因此M/G 殘基比率低的褐藻膠寡糖表現出突出的抗菌活性并可以用來制備抗菌劑[148]。
4)制備促進角質細胞增殖的褐藻膠寡糖 褐藻膠和褐藻膠寡糖也可以應用于治療脫發癥,因為它們不僅可以作為凝膠基質嵌入和裝載活性成分,還可以促進角質細胞增殖。其中,具有由末端還原的poly-MG 組成的褐藻膠寡糖組分是角質形成細胞生長最有效的激活劑[149]。
4 褐藻膠乙酰化
細菌中的褐藻膠在O2/O3 位上發生乙酰化,增加了自身的水合能力和黏度,有助于聚合物作為保護膜靠近細菌并阻礙免疫細胞的運動,同時乙酰化還使褐藻膠裂解酶無法作用于M 殘基,從而保護褐藻膠不被裂解成褐藻膠寡糖。然而到目前為止還沒有發表過詳細的褐藻膠乙酰化機制。
4.1 褐藻膠乙酰化機制初步分析
目前對銅綠假單胞菌中乙酰化褐藻膠的生物合成途徑及相關基因已經基本了解,研究表明AlgI、AlgJ 和AlgF 形成一個多蛋白質復合體,并借助Alg8 和AlgX 形成的分泌多蛋白質復合體與包膜上的褐藻膠發生聚合作用,進行褐藻膠的O-乙酰化反應過程。雖然AlgI、AlgJ 和AlgF 不是褐藻膠合成蛋白質復合體的一部分,但它們是褐藻膠乙酰化所必需的,Chanasit 等也通過實驗證明了這一點[150]。AlgI 是一種膜結合型O-乙酰基轉移酶,其從未知的細胞質乙酰基供體通過細胞質膜轉移乙酰基,而AlgJ 附著在內膜上,將乙酰基轉移到AlgX,AlgX 是甘露聚糖O-乙酰化酶,包含一個碳水化合物結合模塊,能夠將M 殘基乙酰化[151]。
研究表明乙酰化程度高的褐藻膠的異構化程度較低[152],原因可能是乙酰化酶AlgX 和周質異構酶AlgG 均是修飾酶復合體的一部分,可以看做是多糖底物的固定化酶,兩者均能夠結合M 殘基并進行修飾反應,但目前尚不清楚兩種酶在與褐藻膠的結合上是否存在競爭關系[126]。
4.2 研究乙酰化機制的意義
高乙酰化褐藻膠具有的黏度高、水合能力強、G殘基含量低等特點使其在組織工程開發和再生醫學中的水凝膠和支架以及含有生物活性膠囊化合物的傷口敷料等眾多材料方面具有巨大潛力[153]。
銅綠假單胞菌形成以乙酰化褐藻膠為主要成分的生物被膜,可以提高生物被膜的穩定性,從而有助于細菌抵抗惡劣的生存環境。同時相比于浮游細菌,生物被膜內的銅綠假單胞菌對抗生素的耐藥性提高了10~1 000 倍,乙酰化褐藻膠是導致細菌耐藥性和細菌免疫逃逸的主要原因[154]。因此研究褐藻膠的乙酰化機制對于治療銅綠假單胞菌感染也具有重要意義。
5 褐藻膠脫乙酰化酶
乙酰化的褐藻膠可以自發產生脫乙酰化,但在正常條件下反應緩慢,前文中提到,胞外甘露聚糖C5 差向異構酶可以催化產生G,而這些酶不能催化已經乙酰化的殘基,根據對AlgE4 的晶體結構的分析表明這是由于空間位阻的限制,因此,為了獲得高G 含量的褐藻膠,需要對已經乙酰化的殘基進行去乙酰化。
目前還沒有發現假單胞菌能夠產生連續G 殘基的褐藻膠,但是人們在丁香假單胞菌中發現了一種分泌性甘露聚糖C5 差向異構酶PsmE,該酶能夠在體外催化生成連續的G 殘基聚合物,PsmE 能夠催化乙酰化底物進行差向異構化反應,是一種具有脫乙酰基和C5 差向異構功能的雙功能酶,使得菌株產生的褐藻膠具有較低乙酰基數量和較高G 殘基含量的特性[125]。
6 展 望
盡管目前已經表征了大量的褐藻膠裂解酶,特別是對于來自海洋細菌中的酶,但對極端環境中的裂解酶知之甚少,一些新發現的褐藻膠裂解酶表現出的新穎的酶學性質和作用機制也沒有得到清晰地解釋。縱觀近年來發表的關于褐藻膠裂解酶分子改造的相關文章,對酶的改造仍舊需要建立在對酶結構和作用機制分析的基礎上。因此現下仍舊迫切需要篩選、克隆和表征新型褐藻膠裂解酶并解析其酶學性質,了解褐藻膠裂解酶的調控機制。同時隨著褐藻膠裂解酶在生物工業上應用的深入,迫切需要利用固定化和分子改造等手段以改善酶的催化效率、特殊性能和產量,以滿足其日益增加的商業需求。
MC5E 作為一種生物工具酶在生產褐藻膠及具有特殊M/G 殘基比率和序列分布的褐藻膠寡糖方面具有重要意義,至今人們已經分離和表征了幾種具有不同性質的MC5Es 并根據其晶體結構初步闡明了其催化機制,但對于MC5Es 的結構和反應機制還需要進一步探索與研究。由于已知天然MC5Es 的熱穩定性和催化效率并不理想,因此,需要通過計算機輔助設計、定點突變以及DNA 修飾等方式提高酶的催化活性和酶學性質,推動MC5Es 的工業化應用。
褐藻膠修飾酶作為一種生物技術工具表現出了重要的生物活性和應用價值。目前人們在褐藻膠裂解酶和甘露聚糖C5 差向異構酶上的研究較深,而對于褐藻膠乙酰化和脫乙酰化的了解尚未深入。隨著結構分析技術不斷發展,對褐藻膠修飾酶作用機制和條件的不斷深入了解,可以利用各種酶的產物特性進行配合生產目標褐藻膠分子,滿足不同的商業功能需求。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
