苦蕎蛋白源肽AFYRW對脂多糖誘導的人臍靜脈內皮細胞損傷的影響
2023-03-09 13:55:24張禮林王筑婷莫曉川許慶忠李紅梅
食品科學 2023年3期
關鍵詞:苦蕎
肖 怡,左 婕,鄧 艷,張禮林,王筑婷,莫曉川,許慶忠,李紅梅*
(貴州醫科大學基礎醫學院,生物化學與分子生物學教研室,貴州 貴陽 550025)
心腦血管疾病是全球性疾病死亡的主要原因之一,其發病率在中國呈逐年上升趨勢[1]。動脈粥樣硬化(atherosclerosis,AS)是導致心腦血管疾病的主要原因。血管炎癥在AS的發生發展中起重要作用。血管內皮的炎癥反應可上調多種細胞黏附分子的表達,包括血管細胞黏附分子-1(vascular cell adhesion molecule-1,VCAM-1)、細胞內黏附分子-1(intracellular adhesion molecule-1,ICAM-1)和E-選擇素,同時會導致多種促炎細胞因子的大量釋放,如白細胞介素(interleukin,IL)-6、IL-8和單核細胞趨化蛋白-1(monocyte chemoattractant protein-1,MCP-1)。黏附分子和促炎細胞因子的增多會促進白細胞黏附,增加血管通透性[2]。脂多糖(lipopolysaccharide,LPS)作為革蘭氏陰性菌細胞壁外壁的組成成分,能夠誘導全身性炎癥和敗血癥,被廣泛用于誘導人臍靜脈內皮細胞(human umbilical vein endothelial cells,HUVECs)損傷,是誘導細胞損傷的經典模型[3]。LPS誘導內皮細胞炎性損傷的相關機制較多,其中通過激活核因子κB(nuclear factor κB,NF-κB)下游信號通路,放大炎癥反應,被認為是最經典的機制之一[4]。因此,研究內皮細胞炎性損傷的作用機制,改善內皮細胞的功能,是防止心血管疾病發展的有效措施。
目前,從各種食源性動植物中尋找具有抑菌、抗氧化等多種重要生理功能的活性肽已成為研究的一個熱點。生物活性肽一般是由2~20 個氨基酸構成的短肽,分子質量大多在3 kDa以下,對生物機體的生命代謝活動有益或具有一定生物活性。生物活性肽常常隱藏于蛋白質一級結構序列中的一些特定區域內,必須釋放它們才能發揮作用[5]。已報道的生物活性肽具有多種生理功能,包括抗氧化、抗高血壓、抗病毒、抗腫瘤和抗菌等[6]。
苦蕎學名韃靼蕎麥,別名萬年蕎、野蘭蕎、蕎葉七,隸屬于蓼科(Polygonaceae)蕎麥屬(Fagopyrum),與何首烏、大黃等同屬蓼科,是我國藥食同源文化的典型體現。據《本草綱目》記載,苦蕎麥有益氣力、利耳目、降氣、寬腸、健胃等功效,能治療痢疾,咳嗽、水腫、喘息、燒傷等疾病。苦蕎營養豐富,富含蛋白質、維生素、膳食纖維、黃酮類化合物和多種微量礦物質元素等營養物質[7]。貴州威寧地區廣泛種植苦蕎,課題組前期采用堿性蛋白酶酶解苦蕎清蛋白,經多種色譜分離及質譜鑒定,得到一種新的具有抗氧化作用的活性五肽AFYRW(Ala-Phe-Tyr-Arg-Trp),分子質量為741.85 Da,其對羥自由基和1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-trinitrophenylhydrazine,DPPH)自由基的半數清除濃度(half maximal inhibitory concentration,IC50)分別為0.65 mmol/L和0.64 mmol/L。此外,AFYRW還具有較強的還原力和抑制脂質過氧化的作用[8]。本研究以HUVECs為研究對象,建立LPS誘導的內皮細胞損傷模型,觀察AFYRW對LPS誘導的內皮細胞炎性損傷的保護作用,為苦蕎的深加工及苦蕎活性肽的開發提供基礎實驗依據。
1 材料與方法
1.1 材料與試劑
肽AFYRW(純度≥98%)由上海淘普生物科技有限公司合成。HUVECs、內皮細胞培養基(endothelial cell medium,ECM) 美國Sciencell研究實驗室;BCA蛋白定量試劑盒、LPS、蛋白磷酸酶抑制劑、牛血清白蛋白、2’,7’-二-(2-羧乙基)-5(6)-羧基熒光素乙酰甲酯(2’,7’-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein acetoxymethyl ester,BCECF-AM) 北京索萊寶科技有限公司;細胞增殖及細胞毒性檢測試劑盒 大連美侖生物技術有限公司;p65、p-p65 抗體 美國Cell Signaling Technology公司;內皮型一氧化氮合酶(endothelial nitric oxide synthase,eNOS)、誘導型一氧化氮合酶(inducible nitricoxide synthase,iNOS)、ICAM-1、VCAM-1、IL-6、腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)抗體 武漢三鷹生物技術有限公司;NO試劑盒 南京建成生物工程研究所。
1.2 儀器與設備
iMark多功能酶標儀 伯樂生命醫學產品(上海)有限公司;TS100熒光倒置顯微鏡 日本Nikon公司;BB150二氧化碳培養箱 賽默飛世爾科技(中國)有限公司;H1650R臺式高速冷凍離心機 德國Hermle公司;TGL-16B臺式離心機 上海安亭科學儀器廠;SLIK-O3000-S數顯圓周搖床 美國賽洛捷克公司:DYY-6C型穩壓穩流電泳儀、Western blot制膠器及電轉移裝置 北京六一儀器廠;Tanon-4600SF凝膠成像系統上海天能公司。
1.3 方法
1.3.1 AFYRW的高效液相色譜和質譜鑒定
AFYRW經上海淘普生物科技有限公司合成,并采用高效液相色譜(high performance liquid chromatography,HPLC)和電噴霧離子化質譜(electrospray ionization mass spectrometry,ESI-MS)進行純度和成分分析。檢測條件為:色譜柱為kromasil C18-5(4.6 mm×150 mm),流動相A:乙腈(含0.1%的三氟乙酸),流動相B:水(含0.1%的三氟乙酸),采用梯度洗脫,上樣質量濃度為1 mg/mL,上樣體積為20 μL,流速1.0 mL/min,檢測波長214 nm。
1.3.2 細胞培養
細胞復蘇:HUVECs用ECM培養基培養(含5%胎牛血清、1%內皮生長因子、100 U/mL青霉素、100 U/mL鏈霉素)。將液氮罐中的凍存細胞取出,快速將其放入37 ℃水中反復搖晃約1 min,再將細胞從凍存管吸出轉移到裝有預熱培養基的新的培養瓶中,不用離心,搖晃均勻于培養箱中培養,次日換液及傳代培養。
細胞傳代:當細胞生長密度達80%~90%時,吸棄培養基,加入緩沖液清洗2 次,加入1 mL 0.25%胰蛋白酶消化1 min。倒置顯微鏡下觀察到細胞成塊脫落時,立即加入培養基終止消化。輕輕吹打培養瓶收集細胞,細胞懸液1 000 r/min離心5 min,棄上清液,加入1 mL培養基重懸細胞,根據需求加入不同的培養瓶。
細胞計數:細胞重懸后吸取10 μL從蓋玻片邊緣加入,使細胞懸液全部充滿計數室。首先計算周邊4 個中方格內的細胞總數,再取平均數,根據實際需要對細胞懸液進行二次稀釋。
細胞分組和藥物干預:細胞狀態良好時即可分組,分為4 組:1)對照組;2)模型組:質量濃度0.125 μg/mL LPS;3)低劑量組(AFY5組):質量濃度0.125 μg/mL LPS+質量濃度5 μg/mL AFYRW;4)高劑量組(AFY50組):質量濃度0.125 μg/mL LPS+質量濃度50 μg/mL AFYRW。AFYRW干預保護24 h后,再用LPS作用1.5 h建立炎癥模型。
1.3.3 CCK8檢測細胞生存活力
以每孔4×103個的數量接種于96 孔板中,混勻后每孔加入100 μL細胞懸液,每組設置4 個平行復孔。等待細胞貼壁生長至融合狀態,分組給藥干預24 h后加入LPS作用1.5 h。造模結束后每孔加入10%的CCK8溶液,加液過程勿產生氣泡,混勻后放回培養箱中繼續培養0.5 h,450 nm波長處測定各孔吸光度。空白孔只含CCK8溶液,實驗重復3 次。細胞存活率按式(1)計算。
1.3.4 Western blot檢測相關蛋白的表達
冰上裂解提取細胞總蛋白,BCA法進行蛋白濃度測定。根據蛋白分子質量制備合適濃度的膠。10 μg總蛋白上樣、電泳,當溴酚藍電泳至膠邊緣時終止。制作轉膜體系,轉膜時間2 h。用5%牛血清封閉2 h。按一定比例稀釋抗體,一抗過夜孵育,TBST洗膜3 次,每次10 min。二抗室溫孵育2 h,TBST洗膜3 次,每次10 min。化學發光試劑顯色后凝膠成像系統拍照。收集圖像后用Image J軟件分析各條帶灰度值。
1.3.5 NO含量的檢測
調整HUVEC細胞密度為4×105個/mL,接種于6 孔培養板,于37 ℃、5% CO2培養箱中培養24 h,分別加入終質量濃度為5、50 μg/mL的AFYRW孵育24 h后,加入終質量濃度為0.125 μg/mL的LPS繼續孵育1.5 h,棄去LPS刺激的培養液后,用緩沖液清洗2 次,胰酶消化后收集細胞1 000 r/min離心5 min,超聲破碎制備細胞懸液,BCA定量蛋白濃度,根據試劑盒說明書進行測定,空白孔為雙蒸水和顯色液,標準品為20 μmol/L亞硝酸鈉,取上清液于酶標儀550 nm波長處測定各孔OD值,根據公式(2)計算NO含量。
式中:A標準為標準孔吸光度;c標準為標準品濃度;ρpr為細胞勻漿蛋白質量濃度/(g/L)。
1.3.6 單核細胞黏附實驗
細胞接種于24 孔板中等待貼壁,分為4 組:1)對照組;2)模型組:3)低劑量組(AFY10組):質量濃度0.125 μg/mL LPS+質量濃度10 μg/mL AFYRW;4)高劑量組(AFY50組):質量濃度0.125 μg/mL LPS+質量濃度50 μg/mL AFYRW。干預結束后將THP-1細胞1 000 r/min離心3 min,1640培養基重懸細胞后用計數板計數,使用BCECF-AM(5 μmol/L)標記重懸的THP-1細胞,混勻后培養箱中培養30 min,培養結束后用0.01 mol/L pH 7.2~7.4磷酸鹽緩沖液(phosphate buffered saline,PBS),清洗3 遍,洗掉多余的BCECF-AM。加入到HUVECs中共同培養1 h,注意避光,PBS沖洗未黏附的細胞,倒置熒光顯微鏡下拍照。
1.4 數據處理與分析
所有實驗均重復3 次。各組實驗數據以平均值±標準差表示,采用SPSS 19.0軟件進行統計分析,采用方差齊性檢驗和單因素方差分析對數據進行比較,以P<0.05為差異有統計學意義。
2 結果與分析
2.1 AFYRW的HPLC和質譜鑒定結果
經HPLC測定,AFRYW的純度為95%(圖1A)。經ESI-MS測定其分子質量為741.85 Da(圖1B)。

圖1 合成AFYRW的HPLC(A)和質譜(B)圖Fig.1 High performance liquid chromatogram (A) and mass spectrum (B) of synthetic AFYRW
2.2 AFYRW對LPS誘導的HUVECs細胞活力的影響
如圖2所示,與對照組相比,質量濃度0.125 μg/mL LPS 作用內皮細胞1.5 h 后,細胞存活率顯著降低(P<0.05);與模型組相較,不同質量濃度的AFYRW預處理24 h,再加入LPS刺激1.5 h,細胞活力均顯著提高(P<0.05),AFY50組尤為顯著。以上結果說明AFYRW能降低LPS誘導的細胞死亡。
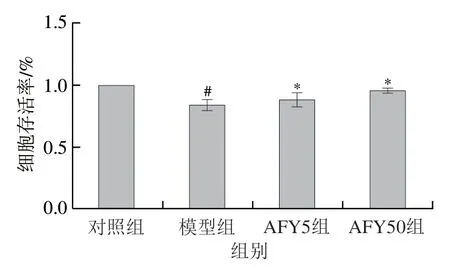
圖2 AFYRW對LPS誘導的HUVECs細胞活力的變化影響Fig.2 Effect of AFYRW on cell viability of HUVECs induced by LPS
2.3 AFYRW對LPS誘導的HUVECs細胞中NO含量的影響
由于NO在內皮功能障礙中起著重要作用,因此檢測了AFYRW對LPS誘導的HUVECs中NO水平的影響,結果如圖3所示。與對照組相比,LPS刺激后,NO含量顯著增多(P<0.05);與模型組相比,低質量濃度AFYRW預處理24 h后NO含量明顯降低,高質量濃度AFYRW可顯著抑制NO的生成(P<0.05)。由于iNOS和eNOS對NO生成的調節的影響,因此檢測了LPS及AFYRW+LPS作用下HUVECs中iNOS和eNOS的相對表達量。如圖4所示,與對照組相比,LPS刺激下,HUVECs的iNOS相對表達量顯著增加(P<0.05)、eNOS相對表達量無顯著變化(P>0.05),從而導致NO生成過量。結果表明AFYRW可通過降低NO的生成進而減輕細胞損傷。

圖3 AFYRW對LPS刺激下HUVECs的NO含量的影響Fig.3 Effect of AFYRW on the content of NO in HUVECs under LPS stimulation

圖4 AFYRW對LPS刺激下HUVECs的iNOS、eNOS蛋白相對表達量的變化影響Fig.4 Effect of AFYRW on iNOS and eNOS protein expression under LPS stimulation
2.4 AFYRW對LPS刺激下單核細胞黏附的影響
單核細胞黏附、聚集在內皮上是動脈硬化發生早期的表現。單核-內皮黏附實驗被用來觀察ARYRW是否能抑制LPS刺激下單核細胞的黏附。如圖5、6所示,與對照組相比,LPS刺激能明顯誘導單核細胞THP-1黏附到HUVECs;加入AFYRW的實驗組中熒光標記的THP-1細胞明顯低于LPS組,并且AFYRW質量濃度越高,標記的THP-1細胞數量越少。結果表明AFYRW能夠抑制LPS誘導單核細胞THP-1對HUVECs細胞的黏附。

圖5 單核細胞對內皮細胞的黏附性檢測結果(10×)Fig.5 Determination of monocyte adhesion to endothelial cells (10×)

圖6 單核細胞對內皮細胞的黏附性檢測結果Fig.6 Determination of monocyte adhesion to endothelial cells
2.5 AFYRW對LPS刺激下HUVECS黏附分子和炎癥因子表達的影響
血管內皮細胞損傷后,可分泌多種黏附分子和促炎細胞因子,趨化單核細胞黏附于血管內皮。與對照組比較,LPS刺激后,細胞中的VCAM-1、ICAM-1、IL-6、TNF-α蛋白的相對表達量顯著增加(P<0.05),說明LPS誘導了HUVECs的炎性損傷;與模型組比較,低、高質量濃度的AFYRW均可降低VCAM-1、ICAM-1、IL-6、TNF-α蛋白的相對表達量,高劑量組效果尤其極顯著或顯著(P<0.01、P<0.05),炎癥有所緩解(圖7)。說明AFYRW可減輕LPS誘導HUVECs的炎性損傷。NF-κB信號通路在LPS誘導的HUVECs的黏附分子及炎癥因子表達中起著關鍵作用。與對照組比較,LPS的刺激使p-p65蛋白相對表達量顯著增加(P<0.05);與模型組相比,低劑量AFYRW可顯著降低該蛋白表達(P<0.05),高劑量AFYRW可極顯著降低p-p65的表達(P<0.01)(圖8),說明AFYRW可通過抑制NF-κB信號減輕黏附分子及炎癥因子的表達,從而減緩LPS誘導的炎癥反應。
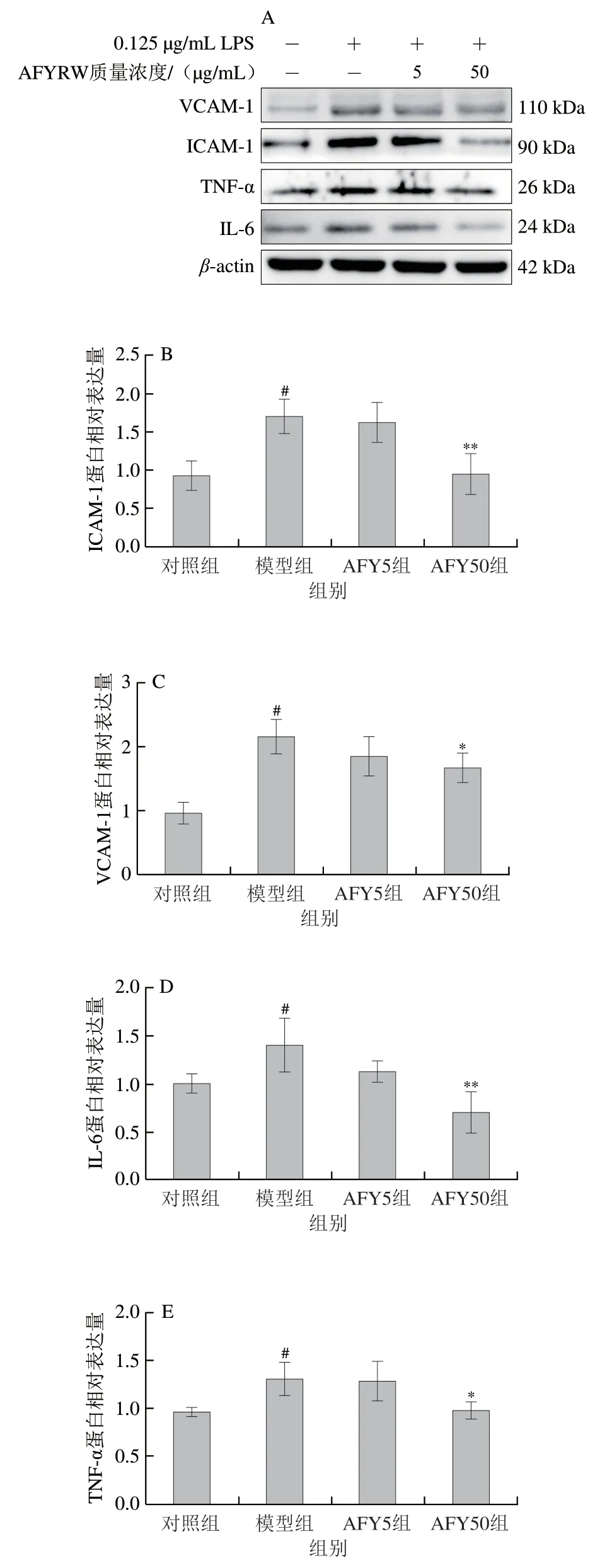
圖7 AFYRW對LPS刺激下ICAM-1、VCAM-1、IL-6和TNF-α表達的影響Fig.7 Effect of AFYRW on the protein expression of ICAM-1,VCAM-1,IL-6 and TNF-α in HUVECs under LPS stimulation

圖8 AFYRW對LPS刺激下p-p65表達的影響Fig.8 Effect of AFYRW on the protein expression of p-p65 under LPS stimulation
3 討論
飲食因素在維持正常免疫系統功能方面發揮著至關重要的基礎和核心作用[9]。生物活性肽作為蛋白質分解的產物,不僅能像蛋白質一樣改善機體的營養狀況,還具有更強的抗氧化和調節免疫的作用[10]。研究認為生物活性肽的免疫效應與肽序列中的氨基酸的物理化學特性有關,如帶有正電荷的精氨酸的生物活性肽可結合并激活免疫細胞的趨化因子受體,色氨酸可被免疫細胞表面的阿片受體識別[11]。來源于苦蕎蛋白的抗氧化肽AFYRW序列中含有精氨酸和色氨酸,同時考慮氧化應激與炎癥之間的關系,因此,本實驗主要研究AFYRW對LPS誘導的血管內皮細胞的炎癥反應的改善作用。
慢性血管炎癥和隨后單核細胞與內皮細胞的黏附是AS發生和發展的基本環節[12]。因此,抑制炎癥反應和單核細胞與內皮細胞的黏附可能是預防AS的有效策略[13]。血管內皮細胞損傷后,可分泌VCAM-1和ICAM-1等多種黏附分子,這些物質可趨化單核細胞及T淋巴細胞黏附于血管內皮。動脈硬化早期,白細胞黏附、聚集是一個重要環節,正常的血管內皮不與白細胞發生黏附,但在血管硬化部位白細胞黏附于內膜表面,并穿透內皮細胞間連接進入到內膜,內皮細胞表面表達的VCAM-1和ICAM-1可促進白細胞和血管內皮細胞黏附,加速白細胞向血管內皮游走,并促進平滑肌細胞增殖[14-17]。在促炎細胞因子中,IL-6是AS發展過程中的關鍵炎癥分子[18],IL-6激活內皮細胞參與白細胞募集并誘導IL-8、MCP-1和黏附分子的釋放,因此,抑制IL-6可以預防炎癥的發展[19-22]。在本研究中,用AFYRW干預后,可顯著抑制LPS誘導HUVECs上的黏附分子VCAM-1和ICAM-1、炎癥因子IL-6和TNF-α的表達,并減少LPS誘導的THP-1細胞與HUVECs的黏附。以上結果表明AFYRW對血管炎癥具有有益的作用。
NO是血管穩態的關鍵調節因子,NOS催化L-精氨酸產生NO。NO促進動脈血管擴張,抑制血小板聚集、單核細胞黏附至血管內皮上,并抑制平滑肌細胞的增殖。大量研究表明,內皮細胞中NO生成的增加可抑制NF-κB活性并減輕AS[23]。在內皮細胞中,NO通過eNOS催化生成,因此,促進eNOS活化可能是控制AS的有效途徑。然而,研究報道iNOS催化產生的過量NO會對血管功能產生有害的影響[24]。iNOS是炎癥標志物,在炎癥和免疫刺激下,iNOS被活化,催化NO過多產生,使機體發生氧脅迫,引起細胞損傷[25],如過多的NO會引起血管擴張,使血流減慢[26]。在炎癥早期NO可推動炎癥細胞遷移,推動炎癥進程。因此,NO可間接反映血管內皮細胞的受損程度[27]。在本研究中,發現用LPS處理HUVECs后,NO含量明顯升高;AFYRW干預后,NO含量顯著降低,并呈劑量依賴性。通過Western blot檢測eNOS和iNOS的蛋白表達情況,發現eNOS在各組中的表達量沒有明顯變化,而在LPS處理后,iNOS表達明顯增高,經過AFYRW干預后,與模型組相比,iNOS相對表達量顯著下調,說明AFYRW可逆轉LPS誘導的iNOS表達,減少NO的生成,從而減輕細胞損傷。
NF-κB幾乎存在于所有炎癥介質蛋白質基因的啟動子和增強子部位上,NF-κB信號激活可通過調節包括細胞因子、趨化因子和黏附分子在內的各種炎癥介質的表達,在機體免疫中尤為重要[28]。通常情況下,NF-κB以二聚體的形式存在,與抑制蛋白-κB(inhibitors of κB,IκB)結合,存在于胞質中。在刺激因子作用下(如LPS),IκB復合物先發生磷酸化,隨后泛素化降解,NF-κB p65的核定位序列暴露,活化的NF-κB p65可以進入細胞核并與特定的DNA區域結合,促進靶基因的序列表達,如VCAM-1,從而參與細胞各病理生理過程[26],并誘導相關因子的表達,其中包括iNOS,從而增強炎癥反應[29-30]。因此,若能有效地抑制NF-κB的活性,可能有助于阻斷AS的發生發展[31-32]。本研究發現LPS作用內皮細胞后,細胞內NF-κB p65磷酸化水平明顯升高,表明LPS可能促進p65磷酸化,激活NF-κB轉移進入細胞核,產生炎癥反應,AFYRW干預后,p65磷酸化水平明顯下調,減輕炎癥反應。
綜上所述,苦蕎活性肽AFYRW 對LPS 誘導的HUVECs的炎性損傷具有早期預防作用,其作用機制可能與抑制NF-κB的活性有關。
猜你喜歡
農產品加工(2023年14期)2023-08-17 10:32:28
農業科技與信息(2021年8期)2021-12-06 04:24:43
詩潮(2021年11期)2021-11-24 19:55:50
四川農業與農機(2021年1期)2021-01-05 19:15:18
青年歌聲(2018年5期)2018-10-29 03:18:40
現代食品(2018年21期)2018-02-14 20:51:24
食品安全導刊(2018年30期)2018-01-17 05:47:48
廣東飼料(2016年3期)2016-12-01 03:43:12
家庭百事通·健康一點通(2016年4期)2016-04-22 09:16:57
林業與生態(2016年2期)2016-02-27 14:24:01
