暫養(yǎng)凈化及無水保活中太平洋牡蠣活力品質(zhì)與呈味物質(zhì)分析
2023-03-06 12:49:20林恒宗高加龍梁志源秦小明范秀萍林海生曹文紅黃艷平
食品科學(xué) 2023年4期
林恒宗,高加龍,梁志源,秦小明,范秀萍,林海生,曹文紅,黃艷平
(廣東海洋大學(xué)食品科技學(xué)院,國家貝類加工技術(shù)研發(fā)分中心(湛江),廣東省水產(chǎn)品加工與安全重點(diǎn)實(shí)驗(yàn)室,水產(chǎn)品深加工廣東普通高校重點(diǎn)實(shí)驗(yàn)室,廣東 湛江 524088)
太平洋牡蠣(Crassostrea gigas)生長速度快,產(chǎn)量高,肉質(zhì)鮮嫩,為我國北方沿海地區(qū)主要養(yǎng)殖品種。近年來,隨著消費(fèi)模式的改變,內(nèi)陸地區(qū)對鮮活牡蠣的需求越來越高,許多消費(fèi)者通過電商途徑購買鮮活牡蠣。活體牡蠣商業(yè)流通鏈主要包括采捕、凈化、運(yùn)輸、銷售4 個(gè)環(huán)節(jié)[1],各流通環(huán)節(jié)牡蠣會(huì)受到干露、溫度波動(dòng)、缺氧、饑餓、振蕩等逆境脅迫,機(jī)體臟器受到不同程度的損傷,存活質(zhì)量受到顯著影響[2]。但目前國內(nèi)外針對活體牡蠣的研究主要集中在采捕后保活運(yùn)輸階段,如:高加龍等[3]研究發(fā)現(xiàn),香港牡蠣(Crassostrea hongkongensis)在4 ℃條件下流通,其存活率最高、營養(yǎng)成分損失相對較低;Hu Xiaopei等[4]采用電子鼻評價(jià)不同溫度條件下運(yùn)輸?shù)幕铙w牡蠣新鮮度變化,發(fā)現(xiàn)4 ℃保活10 d仍保持良好的鮮活狀態(tài)。在這些研究中大多以存活、質(zhì)量損失、營養(yǎng)成分、微生物變化作為判斷標(biāo)準(zhǔn),難以全面反映牡蠣流通過程中活力及整體品質(zhì)。
脅迫條件下,為維持體內(nèi)環(huán)境的穩(wěn)定,需要有一定的物質(zhì)和能量消耗,因此能量代謝是鑒別貝類活力品質(zhì)變化的關(guān)鍵途徑。三磷酸腺苷(adenosine triphosphate,ATP)是一種高能磷酸化合物,可以為細(xì)胞各項(xiàng)生命活動(dòng)提供能量,而核苷酸能荷(adenylate energy charge,AEC)值可準(zhǔn)確反映貝類活力健康水平。大量研究已證實(shí),當(dāng)貝類AEC值為75%~90%時(shí),處于最優(yōu)生長、可繁殖階段;低于50%時(shí),處于不可逆、死亡階段[5-6]。隨著脅迫過程中能量物質(zhì)被大量消耗,這必然會(huì)對牡蠣風(fēng)味品質(zhì)造成一定的影響,而牡蠣的風(fēng)味品質(zhì)取決于氨基酸、核苷酸、有機(jī)酸、有機(jī)堿、小分子肽、無機(jī)離子含量與組成。目前從物質(zhì)能量代謝及風(fēng)味變化途徑解析長時(shí)低溫?zé)o水保活流通對珍珠龍膽石斑魚(Epinephelus fuscogutatus♀×Epinephelus lanceolatus♂)[7]、羅非魚(Oreochromis niloticus)[8]、蝦夷扇貝(Patinopecten yessoensis)[9]、菲律賓蛤仔(Ruditapes philippinarum)[10]的影響已有相關(guān)報(bào)道,而鮮見關(guān)于太平洋牡蠣商業(yè)流通過程中活力品質(zhì)與呈味物質(zhì)劣變規(guī)律的研究。因此,本研究模擬當(dāng)前國內(nèi)活體太平洋牡蠣流通模式,通過ATP等能量物質(zhì)分析牡蠣在流通過程中活力品質(zhì)變化,通過呈味化合物含量變化考察牡蠣在流通過程中風(fēng)味品質(zhì)變化,探明牡蠣在凈化、降溫預(yù)冷及無水保活中的內(nèi)在聯(lián)系,旨在為優(yōu)化太平洋牡蠣商業(yè)流通過程中管理、提升其存活質(zhì)量提供技術(shù)和理論參考。
1 材料與方法
1.1 材料與試劑
鮮活太平洋牡蠣,平均殼長(127.62±2.07)mm,殼寬(64.11±3.57)mm,殼高(31.98±2.47)mm,濕質(zhì)量(129.51±2.81)g,于2021年10月購自山東威海燈塔水母海洋科技有限公司。牡蠣采捕后保留外殼污泥,每50 kg分裝于鋪有海水冰的泡沫箱中,經(jīng)專用牡蠣冷鏈運(yùn)輸車運(yùn)輸48 h后運(yùn)抵廣東海洋大學(xué)水產(chǎn)品保活流通實(shí)驗(yàn)室。立即用滅菌人工海水清洗表面泥污并挑選富有活力,外殼完整的個(gè)體進(jìn)行暫養(yǎng)凈化,以緩解和消除捕撈和運(yùn)輸操作時(shí)應(yīng)激脅迫。
海水晶 江西鹽通科技有限公司;三磷酸腺苷(adenosine triphosphate,ATP)、二磷酸腺苷(adenosine-diphosphate,ADP)、單磷酸腺苷(adenosine 5’-monophosphate,AMP)、肌苷5’-單磷酸(inosine 5’-monophosphate,IMP)及24 種氨基酸標(biāo)準(zhǔn)品美國Sigma公司;濃鹽酸、硼酸、氫氧化鈉、二水合磷酸二氫鈉、十二水合磷酸氫二鈉、高氯酸溶、氫氧化鉀(均為分析純) 廣州化學(xué)試劑廠。
1.2 儀器與設(shè)備
貝類暫養(yǎng)凈化系統(tǒng) 廣州創(chuàng)嶺水產(chǎn)有限公司;1100液相色譜儀、ZORBAX SB-A色譜柱 美國Agilent公司;YC-800低溫恒溫層析柜 北京亞星儀科公司;SPX-250B生化培養(yǎng)箱 上海博迅實(shí)業(yè)有限公司;PR224ZH精密天平 美國OHAUS公司;5810R冷凍離心機(jī) 德國Eppendorf公司;OSE手持式勻漿機(jī) 北京天根生化科技有限公司;PB-10 pH計(jì) 德國Sartorius公司。
1.3 方法
1.3.1 暫養(yǎng)凈化
凈化流程參照SC/T 3013—2002《貝類凈化技術(shù)規(guī)范》。按太平洋牡蠣養(yǎng)殖海域鹽度(31±0.5)‰,在貝類凈化系統(tǒng)中配制人工海水,開啟增氧設(shè)備持續(xù)曝氣24 h,以流速2.5 m3/h通過紫外燈管循環(huán)滅菌制冷至25 ℃,通臭氧至海水臭氧質(zhì)量濃度0.15 mg/L,將清洗后的牡蠣分裝于塑料筐(600 mm×420 mm×315 mm),置于預(yù)冷的人工海水中暫養(yǎng)凈化24 h。
1.3.2 誘導(dǎo)休眠及無水保活
參照郝爽等[11]方法并加以改進(jìn),凈化后的牡蠣采用聚氯乙烯熱收縮膜逐個(gè)包裹,包裝完成后將牡蠣分裝于塑料周轉(zhuǎn)筐(600 mm×420 mm×315 mm)中,置于恒溫層析柜,采用梯度降溫方式誘導(dǎo)休眠處理。處理方式為調(diào)節(jié)恒溫層析柜,從(25±0.5)℃開始以5 ℃/h的速率降至生態(tài)冰溫休眠溫度0 ℃,溫度每下降5 ℃停留90 min,避免溫度急劇下降造成應(yīng)激損傷。誘導(dǎo)休眠結(jié)束后,迅速將牡蠣分裝于套有內(nèi)膜袋的泡沫箱(450 mm×250 mm×200 mm)中,裝箱前在箱底平鋪一層無菌海水制備而成的冰袋,逐層覆蓋浸足滅菌海水的海綿保濕處理,加蓋后放置在溫度設(shè)定為0 ℃的恒溫層析柜中進(jìn)行無水保活。
1.3.3 樣品采集與制備
各實(shí)驗(yàn)組分別在凈化前、凈化后、休眠后、保活3 d、保活6 d、保活9 d取樣,每組隨機(jī)取牡蠣60 只,迅速開殼刨取肌肉組織,用預(yù)冷的0.86%生理鹽水漂洗,吸水紙拭干表面水分,混合勻漿成肉糜狀液氮速凍放于-80 ℃保存待測。
1.3.4 核苷酸及其關(guān)聯(lián)物測定
參照Pei Fei等[12]的提取方法并加以改進(jìn),準(zhǔn)確稱取4.0 g牡蠣勻漿肉糜,加入20 mL預(yù)冷的10 %高氯酸溶液,均質(zhì),4 ℃、8000 r/min冷凍離心15 min;用5 mol/L氫氧化鉀溶液調(diào)節(jié)pH值至6.8,沉淀物復(fù)提1 次,合并上清液;用超純水定容至100 mL,靜置30 min;取上清液用0.22 μm濾膜過濾后采用高效液相色譜儀測定。
色譜條件:ODS-BP-C18色譜柱(4.6 mm×250 mm,5 μm);流動(dòng)相A:20 mmol/L檸檬酸和40 mmol/L三乙胺混合溶液,并用冰乙酸調(diào)節(jié)pH值至4.8;流動(dòng)相B:純甲醇溶液;柱溫40 ℃;流速1.0 mL/min;樣品進(jìn)樣量10 μL;檢測波長254 nm;記錄峰面積,根據(jù)標(biāo)準(zhǔn)品的保留時(shí)間定性外標(biāo)法定量。AEC值按下式計(jì)算[6]:
1.3.5 游離氨基酸測定
參照GB 5009.124—2016《食品中氨基酸的測定》方法,牡蠣肌肉氨基酸水解處理:稱取100.0 mg牡蠣肉糜于玻璃水解管中,加入濃度為6 mol/L的鹽酸溶液10 mL,滴入3~4 滴苯酚;充氮?dú)? min并密封,110 ℃烘箱中水解22 h;水解完成后冷卻至室溫,加水稀釋定容至50 mL;吸取1 mL水解液95 ℃氮吹揮干,準(zhǔn)確加入濃度為0.01 mol/L鹽酸溶液1 mL;過膜,濾液經(jīng)柱前衍生后利用高效液相色譜儀測定。
色譜條件:ZORBAX Eclipse AAA色譜柱(4.6 mm×150 mm,3.5 μm);柱溫40 ℃;流速1.0 mL/min;檢測波長254 nm;流動(dòng)相A:40 mol/L磷酸二氫鈉(pH 7.8);流動(dòng)相B為乙腈-甲醇-水(45∶45∶10,V/V)。
1.3.6 有機(jī)酸測定
參照GB 5009.157—2016《食品中有機(jī)酸含量測定》方法,準(zhǔn)確稱牡蠣勻漿肉糜5.000 g,加入25 mL超純水均質(zhì),冷凍離心15 min(10000 r/min,4 ℃);取上清液用超純水定容至50 mL,經(jīng)0.22 μm濾膜過濾后采用高效液相色譜儀測定。
色譜條件:ZORBAXSB-AQ 色譜柱(4.6 mm×250 mm,5 μm);流動(dòng)相A為0.1%磷酸溶液;流動(dòng)相B為甲醇;檢測波長210 nm;柱溫40 ℃;流速1 mL/min;樣品進(jìn)樣量10 μL。
1.3.7 無機(jī)離子測定
陽離子:K+、Na+、Ca2+、Mg2+,參照GB 5009.268—2016《食品中多元素的測定》采用火焰原子吸收光譜法測定;陰離子:Cl-,參照GB 5009.44—2016《食品中氯化物的測定》方法進(jìn)行。
1.3.8 滋味物質(zhì)的呈味強(qiáng)度評價(jià)
采用呈味強(qiáng)度值(taste activity value,TAV)評價(jià)單個(gè)呈味物質(zhì)對整體的味覺特征貢獻(xiàn),參照Zhang Ninglong等[13]方法,計(jì)算太平洋牡蠣不同流通階段呈味核苷酸、呈味氨基酸、有機(jī)酸、無機(jī)離子的TAV。
1.4 數(shù)據(jù)處理
實(shí)驗(yàn)結(jié)果均平行測定3 次,數(shù)據(jù)均采用SPSS 26.0對實(shí)驗(yàn)結(jié)果進(jìn)行單因素方差分析(One-Way ANOVA)及組間差異采用Duncan多重比較,顯著性水平α設(shè)為0.05,結(jié)果以表示。
2 結(jié)果與分析
2.1 活力品質(zhì)變化
2.1.1 ATP及ADP測定
ATP提供維持正常生命活動(dòng)所必需的能量,其含量變化能較好地反映貝類的生命狀態(tài)[5]。活體牡蠣在不同流通階段ATP含量呈先上升后下降趨勢(圖1a),牡蠣初始ATP含量較低,為0.944 μmol/g,這可能是離水運(yùn)輸過程中受到低溫、饑餓、干露脅迫導(dǎo)致。經(jīng)24 h暫養(yǎng)凈化,ATP含量明顯高于凈化前水平(P<0.05),可見短期離水的活體牡蠣,在到達(dá)銷售地通過復(fù)水暫養(yǎng)可使機(jī)體達(dá)到新的活力水平。誘導(dǎo)休眠過程中牡蠣處于逆境脅迫狀態(tài),迅速啟動(dòng)自身的應(yīng)激反應(yīng)機(jī)制調(diào)動(dòng)能量物質(zhì)以維持機(jī)體平衡,因此ATP含量較凈化后明顯降低(P<0.05),之后隨保活時(shí)間的延長,ATP含量維持在一個(gè)穩(wěn)定的較低水平(P>0.05)。主要原因是牡蠣處于休眠狀態(tài),以無氧呼吸為主,而ATP供能需氧氣參與[14],因此在保活階段ATP含量處于較低的穩(wěn)定值。Liu Jinyang等[15]發(fā)現(xiàn),蝦夷扇貝在無水保活中ATP含量呈緩慢下降趨勢,與本研究結(jié)果相似。由圖1b可知,ADP變化趨勢與ATP較為相似,在流通中呈下降趨勢,這主要是因?yàn)榱姿峋彼峒っ缚蓪DP轉(zhuǎn)化為ATP為機(jī)體供能,使機(jī)體在逆境脅迫下保持相對穩(wěn)定的平衡狀態(tài)[16]。保活9 d后牡蠣體內(nèi)ADP含量仍維持在較高水平,可見牡蠣仍保持較高活力品質(zhì)。
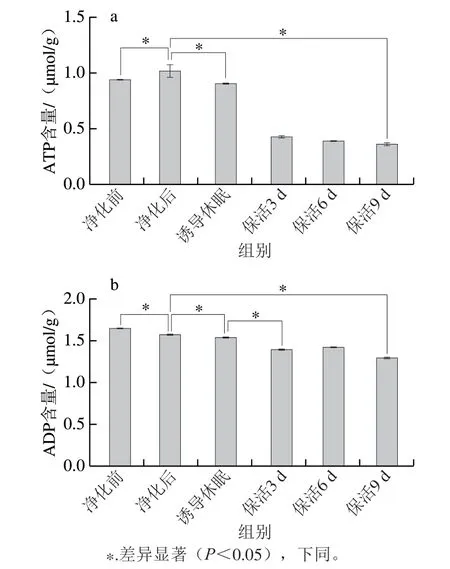
圖1 太平洋牡蠣流通過程中ATP(a)、ADP(b)含量變化Fig.1 Changes in ATP (a),and ADP (b) contents in C.gigas during circulation
2.1.2 AEC值
AEC值被認(rèn)為是描述細(xì)胞能量狀態(tài)的關(guān)鍵指標(biāo),其變化能準(zhǔn)確反映貝類活力變化[6]。圖2顯示,太平洋牡蠣暫養(yǎng)凈化后AEC值顯著高于暫養(yǎng)前水平(P<0.05),保活流通階段處于穩(wěn)定的較低值(P>0.05),這主要是由于ATP及ADP大量消耗所致,但在整個(gè)流通中牡蠣AEC值均大于50%,根據(jù)AEC值與動(dòng)物生理狀態(tài)的關(guān)系,處于應(yīng)激脅迫但可恢復(fù)狀態(tài)[5]。可見,牡蠣在長時(shí)冰溫保活流通過程中仍保持較高的活力品質(zhì)。在已有研究中,Jiménez-Ruiz等[17]發(fā)現(xiàn)長牡蠣在離水運(yùn)輸過程中AEC值呈下降趨勢;劉慧慧等[18]研究表明,菲律賓蛤仔在干露脅迫過程中AEC值有所下降,復(fù)水處置后其活力明顯提升,與本研究結(jié)果相似。
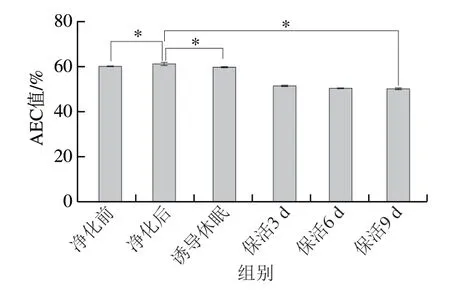
圖2 太平洋牡蠣流通過程中AEC值變化Fig.2 Changes in AEC value in C.gigas during circulation
2.1.3 游離氨基酸組成與含量變化
③融資成本高。一般中小企業(yè)貸款利率高于市場平均利率水平。為彌補(bǔ)和防范貸款風(fēng)險(xiǎn),很少選用信用貸款的方式。最常見的做法是把貸款利率在基準(zhǔn)利率基礎(chǔ)上上浮一定比例,用額外的利息收入去平衡無法收回的貸款。以及,在融資過程中,中間環(huán)節(jié)繁瑣,費(fèi)用過高,這些因素都增加了中小企業(yè)經(jīng)營成本,影響了中小企業(yè)的利潤。
游離氨基酸在貝類應(yīng)對逆境脅迫的能量代謝響應(yīng)中起關(guān)鍵作用[19],太平洋牡蠣不同流通階段游離氨基酸含量變化見表1。活體牡蠣肌肉組織共檢測出24 種游離氨基酸,其中含量最高的為Ala,其次是Trp、Glu、Arg、Asp、Gly。由圖3可以看出,凈化前與凈化后的牡蠣在同一分類分支上,表明其氨基酸無顯著差異;誘導(dǎo)休眠后至保活3、6、9 d分別在3 個(gè)不同分類分支上,表明牡蠣在流通階段氨基酸含量差異明顯。結(jié)合表1分析,暫養(yǎng)凈化期間,牡蠣體內(nèi)12 種氨基酸(Asp、Glu、Ser、Arg、Ala、Gly、Cit、Arg、Ala、Lys、Hyp、Pro)較凈化前增加,提示牡蠣活力恢復(fù),低溫誘導(dǎo)休眠至保活終期,氨基酸含量隨保活時(shí)間的延長緩慢下降,這一現(xiàn)象在菲律賓蛤仔[10]、蝦夷扇貝[20]無水流通中也有報(bào)道,表明游離氨基酸可作為能源,以響應(yīng)機(jī)體在無水保活流通中復(fù)雜的環(huán)境變化,如:溫度波動(dòng)、饑餓、缺氧等。
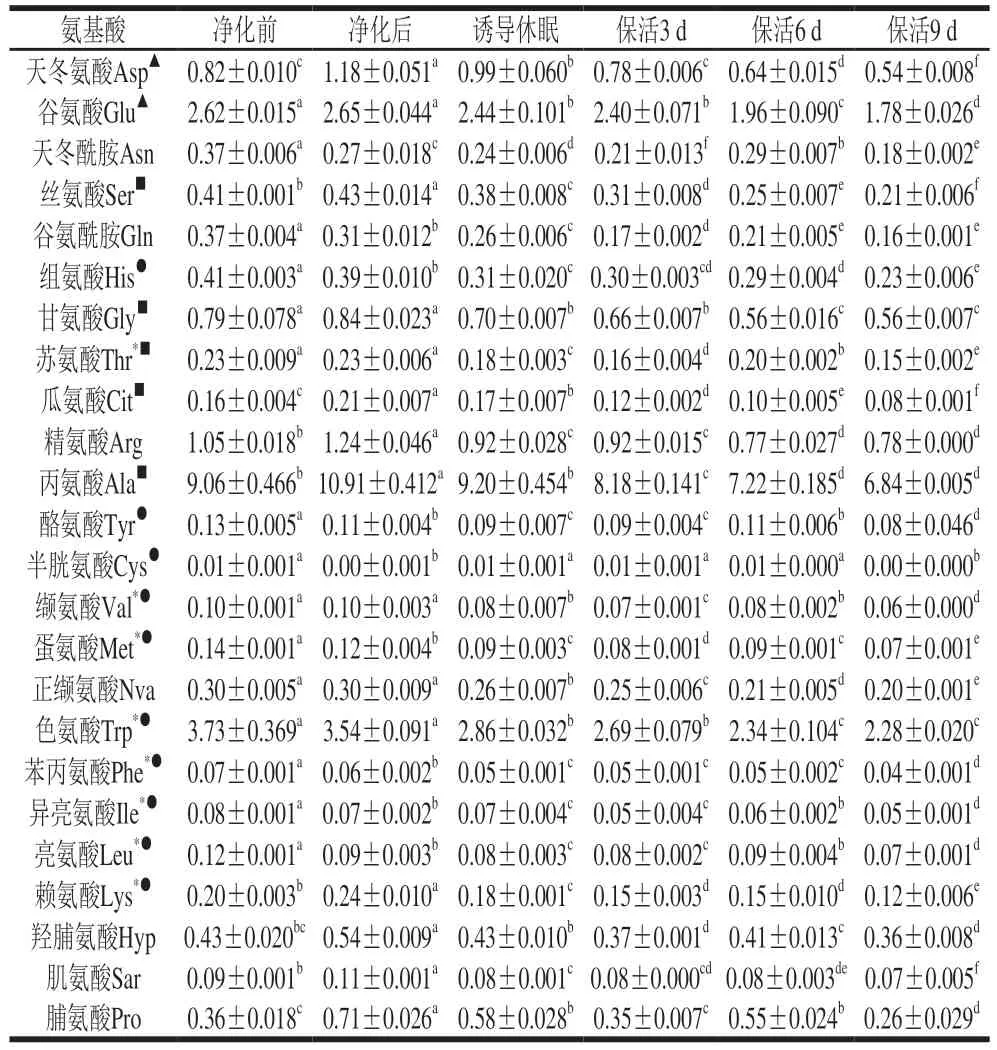
表1 太平洋牡蠣流通過程中游離氨基酸含量變化Table 1 Changes in free amino acid contents of C.gigas during circulation mg/g
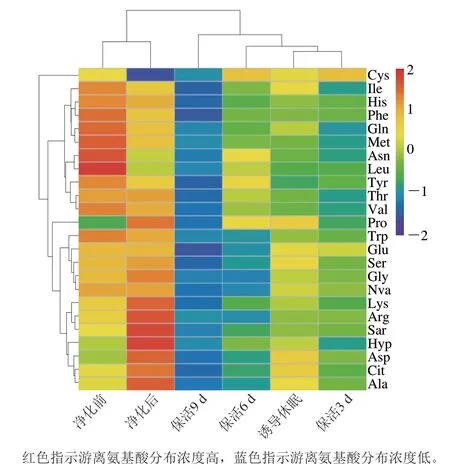
圖3 太平洋牡蠣流通過程中游離氨基酸含量變化熱圖Fig.3 Heat map of changes in free amino acid contents of C.gigas during circulation
如圖4所示,與凈化前相比,牡蠣在凈化后總游離氨基酸(total free amino acids,TFAA)、必需氨基酸(essential amino acids,EAA)、非必需氨基酸(nonessential amino acids,NEAA)均呈明顯上升趨勢,這可能是由于牡蠣離水后長時(shí)處于干露狀態(tài),需消耗大量能量物質(zhì)以維持機(jī)體平衡,復(fù)水凈化后,其脅迫解除,活力恢復(fù),游離氨基酸總量明顯提升;誘導(dǎo)休眠后其總量低于凈化后水平,之后隨著保活時(shí)間的延長,總含量變化與保活時(shí)間呈負(fù)相關(guān),主要原因是牡蠣長時(shí)處于低溫應(yīng)激脅迫狀態(tài),能量代謝從而導(dǎo)致游離氨基酸總量進(jìn)一步降低。對比TFAA、EAA、NEAA含量變化可知,NEAA在流通過程中變化趨勢明顯快于EAA,與Fan Xiuping等[19]對珍珠龍膽石斑魚在冷脅迫過程中NEAA變化趨勢相似,推測NEAA在活體牡蠣流通過程中能量消耗起著關(guān)鍵作用,而EAA則可能用于合成應(yīng)激脅迫下所必需的化合物。
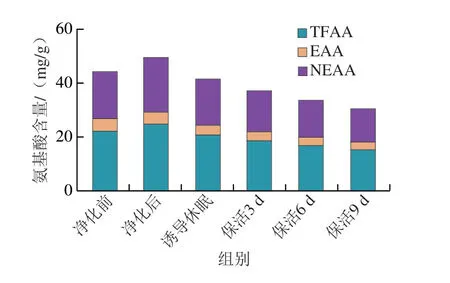
圖4 太平洋牡蠣流通過程中TFAA、EAA、NEAA含量變化Fig.4 Changes in TFAA,EAA and NEAA contents in C.gigas during circulation
2.2 呈味物質(zhì)含量變化
2.2.1 呈味氨基酸含量變化
將表1中游離氨基酸按呈味特性劃分為鮮味氨基酸(delicious amino acid,DAA)、甜味氨基酸(sweet amino acid,SAA)和苦味氨基酸(bitter amino acid,BAA)。由圖5可知,牡蠣在流通過程中SAA>DAA和BAA,SAA為活體牡蠣中主要呈味物質(zhì),含量占60%以上。結(jié)合圖4氨基酸組成變化分析,DAA、SAA和BAA含量變化與TFAA變化基本一致,牡蠣通過暫養(yǎng)凈化DAA(Asp、Glu)、SAA(Ser、Gly、Thr、Ala、Cit)愉悅性呈味物質(zhì)明顯提升,同時(shí)BAA(His、Tyr、Cys、Met、Trp、Phe、Ile、Leu、Lys)非愉悅性呈味物質(zhì)顯著降低。采捕后的牡蠣腸道中含有大量未排出的糞便、食物殘?jiān)⒛嗌车任镔|(zhì),這些殘留物通常被認(rèn)為是牡蠣不良口感及土腥味的來源,可見通過暫養(yǎng)凈化牡蠣鮮甜滋味得到明顯改善[21-22]。保活流通階段,牡蠣整體呈味氨基酸下降明顯,表明在長時(shí)低溫流通過程中對牡蠣風(fēng)味造成了一定影響。步營等[7]在對珍珠龍膽石斑魚在無水保活實(shí)驗(yàn)中發(fā)現(xiàn),苦味氨基酸隨著流通時(shí)間的延長而不斷升高,這與本研究結(jié)果相反,可能是物種間差異、保活方式及流通時(shí)間不同有關(guān)。
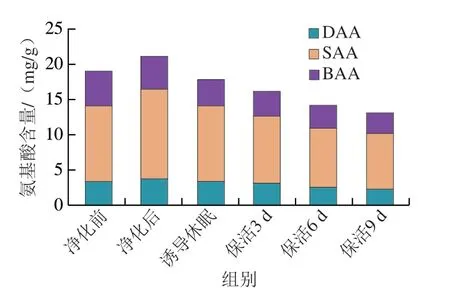
圖5 太平洋牡蠣流通過程中DAA、SAA、BAA氨基酸含量變化Fig.5 Changes in DAA,SAA and BAA amino acid contents in C.gigas during circulation
2.2.2 呈味核苷酸含量變化
呈味核苷酸是評價(jià)貝類滋味的重要指標(biāo),牡蠣在低溫脅迫下機(jī)體由耗氧呼吸轉(zhuǎn)變成無氧呼吸,ATP迅速被分解為ADP→AMP→IMP→HxR→Hx,而這些分解產(chǎn)物影響牡蠣的口感和風(fēng)味。其中IMP具有重要的呈鮮味作用,與天冬氨酸、谷氨酸之間存在協(xié)同效應(yīng),而AMP有抑制苦味的特性,與IMP在誘導(dǎo)鮮甜味方面存在協(xié)同作用,當(dāng)IMP含量較低時(shí),會(huì)賦予其鮮美味道并能增加甜味[23]。太平洋牡蠣在不同流通階段AMP及IMP均能檢出,且AMP、IMP隨著流通時(shí)間的延長呈緩慢積累特征(圖6a、b)。結(jié)合圖1分析,這可能是長時(shí)饑餓及低溫脅迫導(dǎo)致牡蠣機(jī)體代謝發(fā)生顯著變化,ATP、ADP發(fā)生降解,最終導(dǎo)致AMP與IMP的積聚,從而提升牡蠣的鮮甜滋味。大量研究已證實(shí),無水保活有利于機(jī)體鮮甜味核苷酸的積累,如:蝦夷扇貝[20]在無水保活24 h后,肌肉中的IMP含量顯著增加;大菱鲆(Scophthalmus maximus)[23]在無水保活18 h后,AMP與IMP仍維持在較高水平。
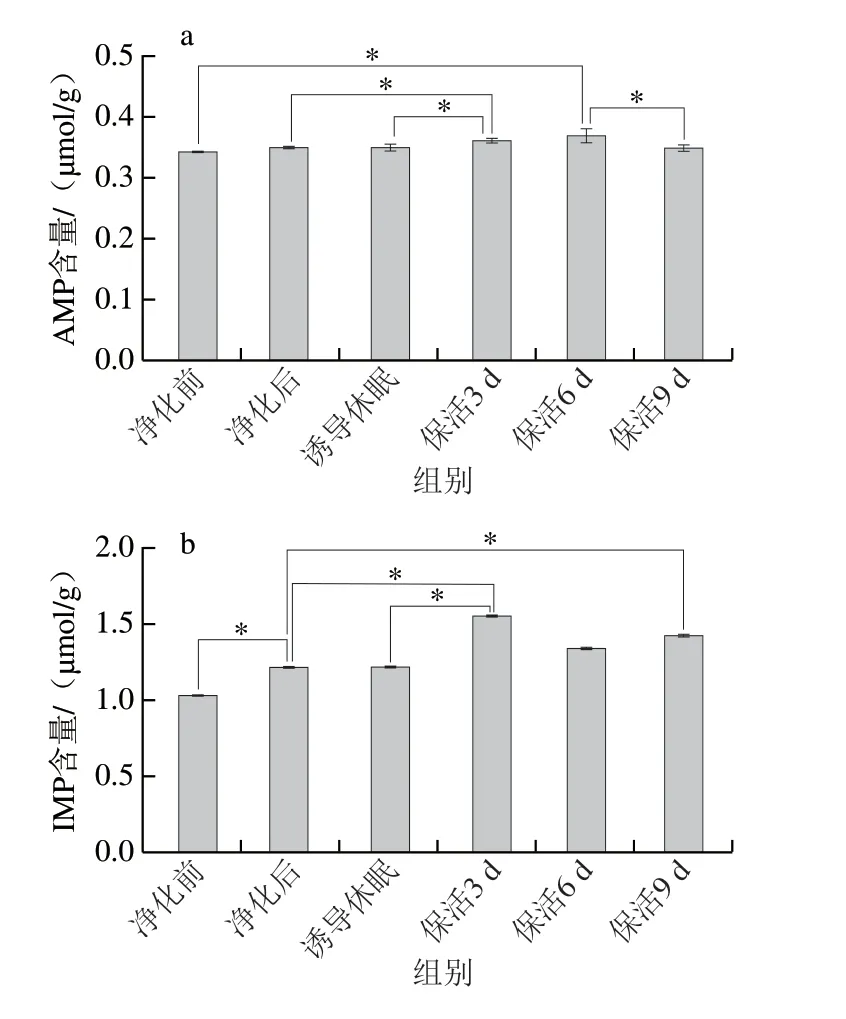
圖6 太平洋牡蠣流通過程中AMP(a)、IMP(b)含量變化Fig.6 Changes in AMP (a) and IMP (b) contents in C.gigas during circulation
有機(jī)酸是水產(chǎn)貝類滋味的重要組成部分,其中乳酸、琥珀酸、蘋果酸已被鑒定存在于活體牡蠣中[24],乳酸是反映機(jī)體能量代謝的關(guān)鍵指標(biāo),也對鮮味有增強(qiáng)作用;蘋果酸呈略帶果香的酸味,而琥珀酸是貝類中主要呈鮮味的有機(jī)酸,來源于貝體內(nèi)豐富的糖原及氨基酸降解生成。由圖7可知,太平洋牡蠣在不同流通階段乳酸、琥珀酸及蘋果酸均能檢出;牡蠣在暫養(yǎng)凈化后乳酸含量顯著高于凈化前水平(P<0.05),誘導(dǎo)休眠至保活終期呈下降趨勢(P<0.05),推測原因是牡蠣復(fù)水后由無氧呼吸轉(zhuǎn)為耗氧呼吸,糖分解代謝乳酸則增加,無水保活過程中低溫條件下牡蠣呼吸及能量代謝減弱,導(dǎo)致乳酸的積聚減少。而蘋果酸及琥珀酸在凈化后降低(P<0.05),在保活流通階段,隨著保活時(shí)間的延長而緩慢增加(P<0.05),這可能是牡蠣在保活初期,呼吸代謝旺盛導(dǎo)致箱體內(nèi)產(chǎn)生大量二氧化碳,隨著時(shí)間的延長從而抑制了機(jī)體呼吸,最終導(dǎo)致牡蠣組織中酸鮮味明顯增加。有機(jī)酸總量在整個(gè)流通階段呈波動(dòng)下降趨勢,但始終維持在110 mg/100 g以上水平,整體而言,牡蠣在長時(shí)保活流通過程中滋味未發(fā)生明顯變化,仍處于鮮活狀態(tài)。
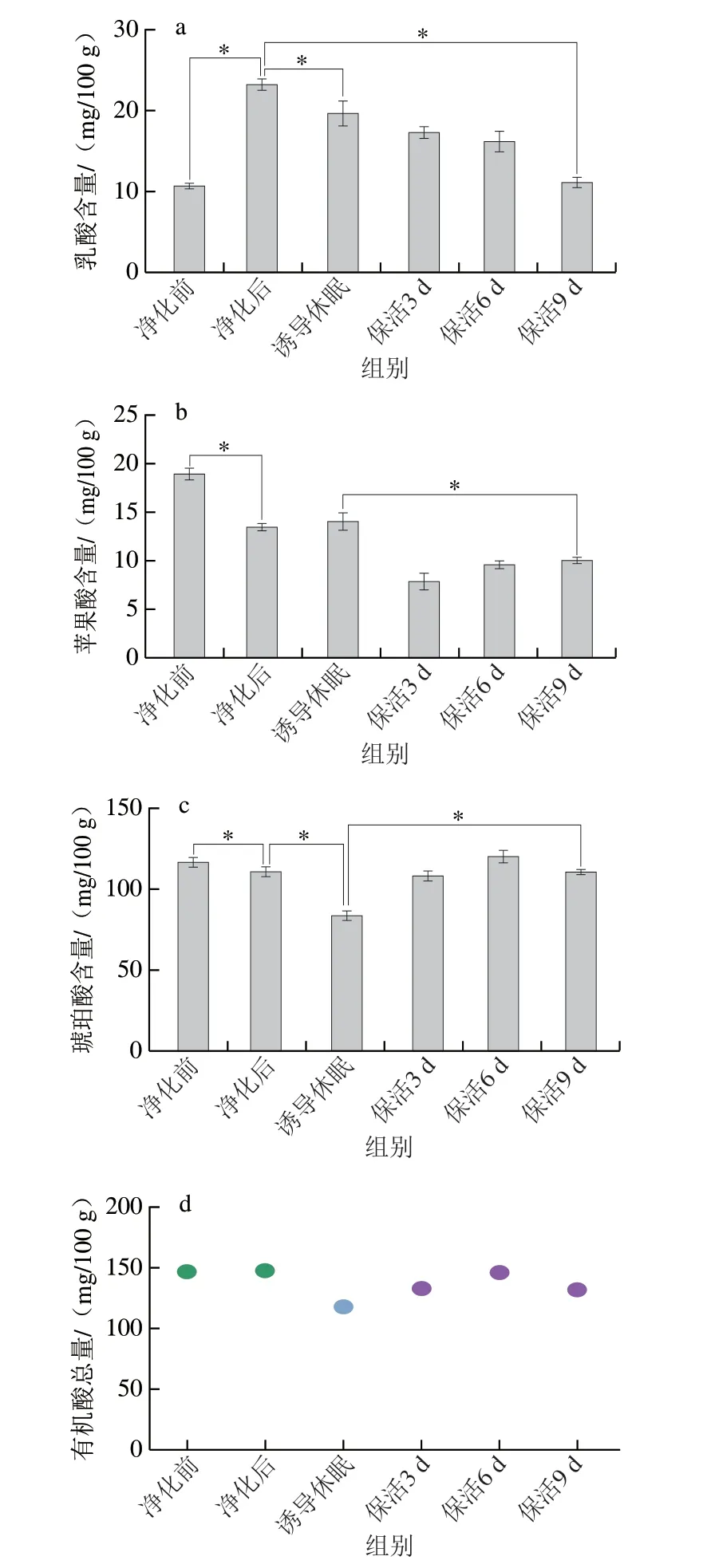
圖7 太平洋牡蠣流通過程中乳酸(a)、蘋果酸(b)、琥珀酸(c)、有機(jī)酸總量(d)變化Fig.7 Changes in amounts of lactic acid (a),malic acid (b),succinic acid (c),and total organic acids (d) in C.gigas during circulation
2.2.4 無機(jī)離子含量變化
無機(jī)離子是貝類中不可缺少的呈味輔助劑及鮮味增強(qiáng)劑,研究表明Na+、K+、Cl-、Mg2+等離子被確認(rèn)為對牡蠣的呈味起關(guān)鍵作用[25]。如:Na+具有濃烈的咸味,對甜、咸、鮮味有增強(qiáng)作用;Cl-可抑制酸味并提高鮮甜味;而K+的缺失會(huì)降低貝類的總體風(fēng)味[26]。如圖8所示,活體牡蠣中共檢測出Na+、K+、Cl-、Mg2+、Ca2+,其中Na+、K+含量最高,其次是Ca2+、Mg2+、Cl-離子。在整個(gè)流通階段除Ca2+損失較大外,其他離子變化不大甚至高于初始水平,可見其在長時(shí)保活流通過程中整體風(fēng)味未發(fā)生明顯改變,仍處于鮮活狀態(tài)。與本研究結(jié)果相似的是,羅非魚[8]在無水保活中無機(jī)離子損失量低于有水保活;珍珠龍膽石斑魚[27]在模擬運(yùn)輸過程中無機(jī)離子損耗較低;近江牡蠣(Crassostrea rivularis)[28]在生態(tài)冰溫保活9 d后Ca2+損耗較大。
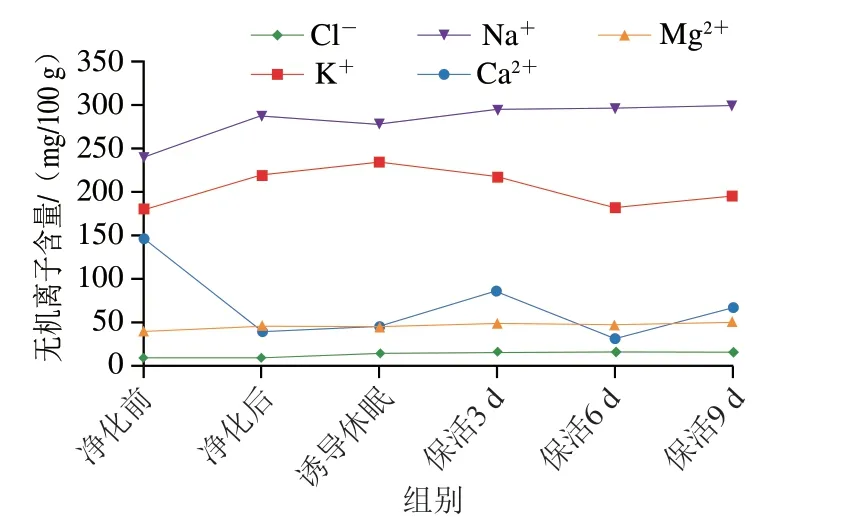
圖8 太平洋牡蠣流通過程中無機(jī)離子含量變化Fig.8 Changes in mineral ion contents of C.gigas during circulation
2.2.5 滋味物質(zhì)的呈味強(qiáng)度評價(jià)
太平洋牡蠣中呈味物質(zhì)的組成、含量、閾值共同作用下使其表現(xiàn)出一定的滋味,而這些滋味物質(zhì)的呈味強(qiáng)度通常以TAV評價(jià),其中TAV大于1表明該物質(zhì)對樣品的呈味有較大貢獻(xiàn),反之則貢獻(xiàn)較小[13]。牡蠣流通過程中滋味物質(zhì)的呈味作用及TAV見表2。活體牡蠣在整個(gè)流通期內(nèi)Asp、Glu、Arg、Ala、IMP、乳酸、琥珀酸、Na+、K+TAV大于1,可見這幾種化合物對活體牡蠣的呈味有顯著影響。雖然其他游離氨基酸、核苷酸、有機(jī)酸、無機(jī)離子的TAV均小于1,但它們之間具有協(xié)同增效作用,如,Glu、Asp與IMP協(xié)同作用下能增強(qiáng)貝類鮮味,也可以改善甜味及增加整體風(fēng)味[23];此外,一些苦味氨基酸其含量低于閾值時(shí),可以增強(qiáng)鮮味和甜味氨基酸的呈味強(qiáng)度[29]。
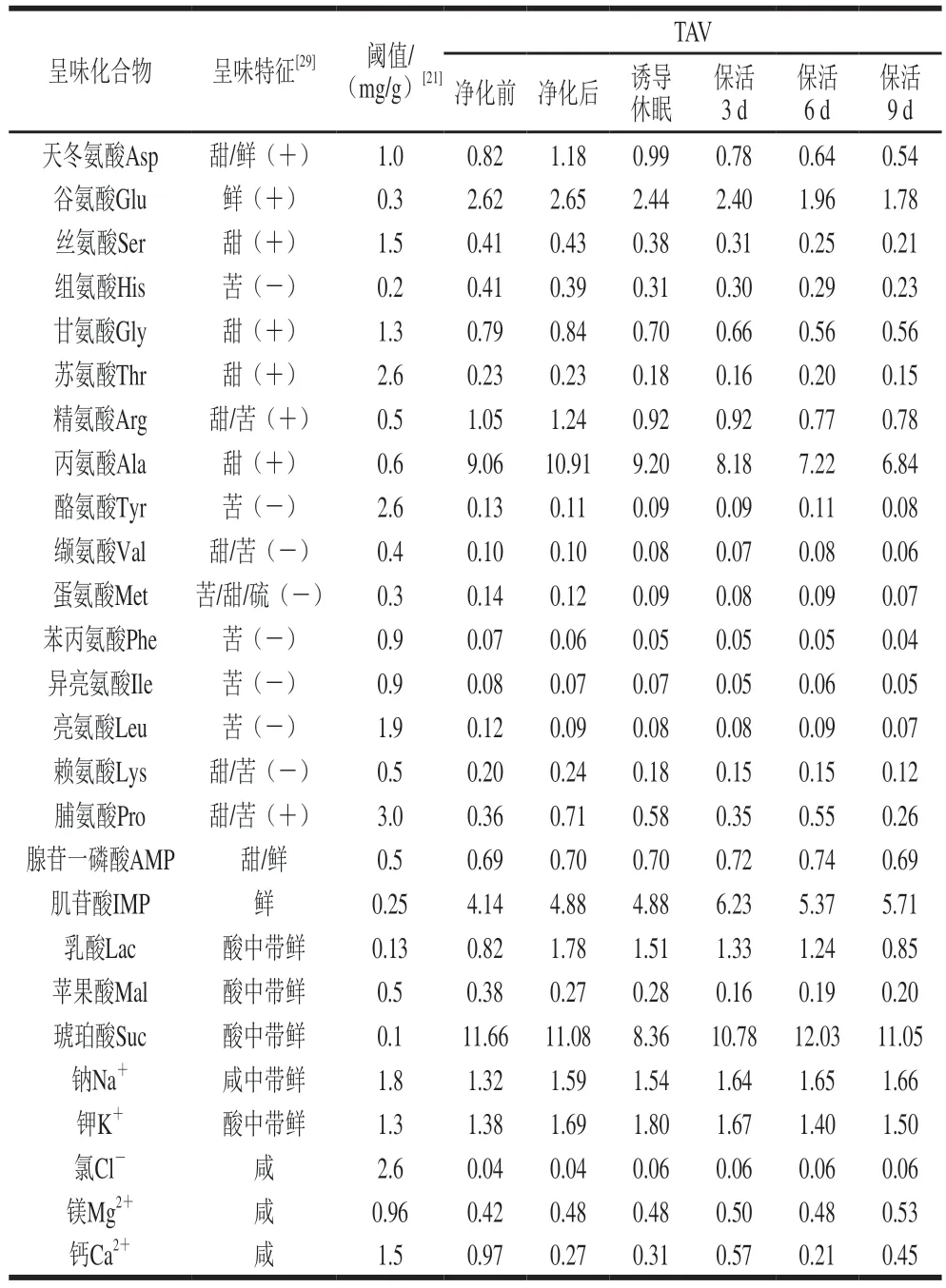
表2 太平洋牡蠣流通過程中滋味物質(zhì)的呈味作用及TAVTable 2 Taste characteristics and TAV of taste substances in C.gigas during circulation
3 結(jié)論
太平洋牡蠣采捕后運(yùn)輸及環(huán)境脅迫對其活力品質(zhì)、呈味物質(zhì)均有顯著影響,通過24 h暫養(yǎng)凈化,機(jī)體活力及風(fēng)味物質(zhì)呈明顯的恢復(fù)性,牡蠣鮮甜滋味得到明顯改善;誘導(dǎo)休眠至保活流通終期,牡蠣遭受低溫及缺氧脅迫,需調(diào)動(dòng)更多能量物質(zhì)以維持機(jī)體平衡,ATP、AEC值處于穩(wěn)定的較低水平,游離氨基酸總量呈下降趨勢;呈味化合物中AMP、IMP出現(xiàn)累積特征,有機(jī)酸、無機(jī)離子呈波動(dòng)下降趨勢,但呈味物質(zhì)未發(fā)生整體性改變。由此得出結(jié)論,太平洋牡蠣采捕后在“暫養(yǎng)凈化24 h-梯度降溫誘導(dǎo)休眠-生態(tài)冰溫保活9 d”模式下可維持較高的活力及風(fēng)味品質(zhì)。
