兩棲動物骨齡學研究進展
2023-02-22 02:49:46李云磊陸宇燕李丕鵬
野生動物學報 2023年1期
李云磊,原 濤,陸宇燕 ,李丕鵬
(1.沈陽師范大學兩棲爬行動物研究所,沈陽,110034;2.濰坊市寒亭區第一中學,濰坊,261100)
在種群生態學研究中,準確判斷種群年齡結構可以為評估種群生存現狀、預測種群未來發展趨勢、調節和管控野生動物種群提供科學依據,并為制定相應的保護利用和管理計劃提供基礎數據[1-3]。種群年齡結構是指不同年齡組別的個體數目在整個種群內的比例或配置情況,目前年齡結構特征主要分為增長型、穩定型和衰退型3 種類型[4],表示種群年齡分布特征最直接的方法是金字塔法,其中橫軸表示分布的比例,縱軸表示年齡。若種群年齡分布特征呈金字塔型,則表示該種群處于增長狀態,屬于增長型;若種群年齡分布特征呈倒金字塔型,則表示該種群處于老齡化狀態,屬于衰退型;若種群年齡分布特征呈鐘形,則表示該種群各年齡組分布相對均衡和穩定,屬于穩定型。
全球兩棲類物種數量呈急劇減少的趨勢,甚至有些物種瀕臨滅絕[5-6]。2012年的統計數據顯示,有2 469 種兩棲類物種數量逐漸減少[7],自20 世紀70年代以來,保守估計有3.1%的兩棲類物種已經滅絕[8],因此,對兩棲類物種種群動態監測和分析尤為重要,動態信息的掌握對兩棲動物生理、生態,甚至形態分類學的研究都具有重要意義[9]。
1 骨齡學基本概述
1.1 骨齡學方法
個體年齡是種群特征研究中重要的信息之一,但在野外調查和后續分析中,很難直接準確地判斷動物個體的年齡。人們逐漸摸索出了借助于一些常規的生物學量度、解剖觀察和特定的實驗技術等間接推斷動物的年齡。兩棲類動物代謝功能的強弱會隨環境溫度的變化而變化[10],這種變化在骨骼生長過程中有明顯的印記。通過骨組織或骨板上的特殊生長標記來確定個體年齡的方式已經在一些動物年齡研究中得到了應用,如龜類,通過在龜板上留下類似年輪的特殊標記線來推斷龜類的年齡[11-12]。這種利用動物生長在骨組織上的特殊標記線來推斷年齡的方法叫作骨齡學方法[13]。
馬常夫[14]曾利用“十氫奈泡脊椎骨”的方法對兩棲動物的年齡進行探究,將解剖出來的動物脊椎骨去除肌肉,自然干燥后放入十氫奈中處理一定時間后,在顯微鏡下可觀察到椎骨關節凹盤處呈現的黑白相間的年齡環帶,以計算個體的年齡。由于凹盤的左右側壁向外傾斜,其上的環帶易被觀察到,但也有個體椎骨凹盤上的年齡環帶不夠清晰,因此對年齡的判斷可能出現誤差。另外,觀察時需要將動物處死,不利于對物種的保護。
目前,在骨齡學方法研究中最為廣泛運用并被廣大研究者接受和認可的是利用兩棲類動物的趾骨或指骨制作石蠟切片[15],HE 染色,觀察骨切片上的生長抑制線(也稱為年輪線,lines of arrested growth,LAG)數目,進而確定個體年齡。該方法操作簡便且得出的結果相對準確[16-18],更重要的是對研究個體損傷有限,不危及生命,但是此法無法確定處于四季交替不明顯、沒有明顯骨骼生長痕跡的動物個體的年齡[19]。
1.2 骨齡學原理
骨齡學方法確定年齡的主要依據是兩棲動物生長發育的年周期變化,在環境溫度適宜、食物和陽光充足的情況下,機體代謝功能很快得到提升,表現為個體生長(包括骨骼的生長)速度加快,即生長期。在環境溫度過低或過高時,兩棲類動物主動調節代謝水平,降低到一定水平后進入冬眠或夏眠狀態,此時機體的生長速度極度減緩,稱為生長的慢速期或停滯期[20]。在這種生長的周期性變化中,兩棲動物長骨骨干中的有機物和礦物質的沉積也呈現明顯的規律性,即快速生長期,骨質的快速生成導致有機物和礦物質沉積密度較低;而生長停滯期的骨質生長雖然緩慢,但有機物和礦物質沉積卻顯現出較高的密度。因此,顯微觀察長骨骨干部切片可清晰分辨出第1 年冬眠期及其前后生長緩慢階段形成的骨環(染色較深)和第2 年生活期形成的骨環(染色較淺)之間形成的同心環狀分界線,這個環狀分界線被稱為LAG[21],依 據LAG 數目可 間接判 斷出個體的年齡。
1.3 骨齡學LAG識別
最早研究幼蛙和成年蛙長骨組織結構的Kastschenko 指出,兩棲動物和哺乳動物在長骨組織學結構上存在著明顯的差異性[22],兩棲動物長骨骨質缺少像哺乳類那樣獨立的、完整的骨單位[23],會呈現出同心圓狀的層層推進的生長方式,這正是骨齡學研究的結構基礎。
對LAG 進行識別時,需要對骨骼切片的結構有一定的了解(圖1),首先在顯微鏡下觀察長骨切面時,骨髓腔的外側有一著色較淺的環狀骨質結構為內骨板,內骨板離心生長,從骨髓腔處推進式向外生長[25-26],通常在經歷了第2 個冬眠期后才可觀察到(1齡蛙的長骨組織切片上未觀察到內骨板結構),之后每年都會在原有內骨板的外側形成1 層新的內骨板層[22]。
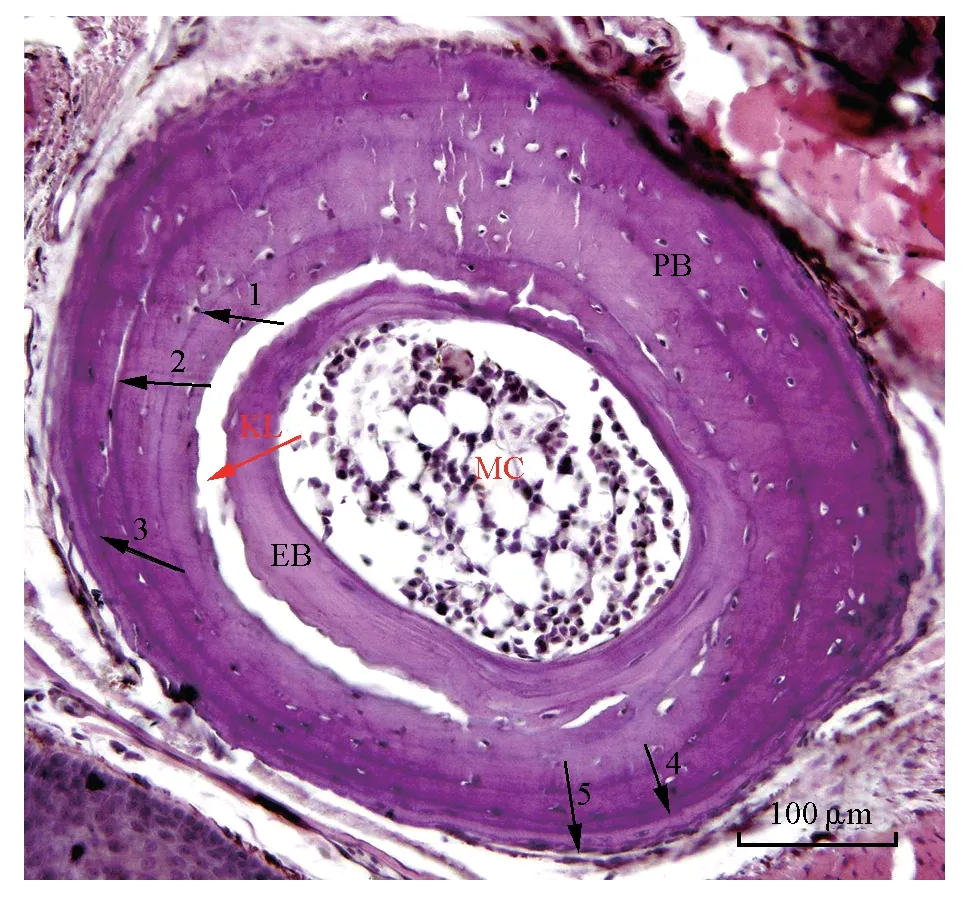
圖1 西藏蟾蜍5齡雄性趾骨切片[24]Fig.1 The phalangeal bone of 5th instar male of Bufo tibetanus
在內骨板與外骨板之間有2 條染色特征不同的環狀線條,靠近內骨板外側的1 條著色較深且較粗的線條以及其外側靠近外骨板內側著色較淺且較細的線條。前者為重吸收線,又稱回復線,其高度礦化,能夠被蘇木精深染色,同時可作為內骨板與外骨板的分界線,在估計動物的年齡時不可用,即沒有年代學價值[27];后者是以首次發現此結構的Kastschenko的名字命名[28],稱Kastschenko 線(KL 線),其組織學結構為1 層均質的軟骨組織。在觀察長骨切片時常可見內、外骨板間出現部分分離的現象,這是由于內、外骨板組織學結構特性不同,在制片過程中化學和物理因素的收縮程度差異導致在KL 線處出現撕裂現象,這種現象并不影響內、外骨板結構的完整性,不妨礙對外骨板上LAG 的計數[29-30]。Kastschenko 認為KL 線的軟骨組織結構可能是幼體軟骨組織的部分殘余,該結構在成體動物骨骼發育中的作用還尚不清楚。也有學者認為KL 與內骨板的形成有關[31]。在KL線附近常常會發生重吸收現象[18],該現象通常發生在近外骨板的區域,有可能會損傷外骨板最內圈的LAG1,致使動物個體年齡有被低估的可能[32-33]。重吸收現象僅出現在最內圈LAG1 局部,并未發現將最內圈LAG1 全部吸收的現象,只要切片質量較好,并仔細觀察,對于LAG 計數的影響可以忽略[34]。Castanet 等[35]認為外骨板骨質的重吸收程度與個體的性成熟有關,性成熟越晚重吸收程度越嚴重,但是對博斯歐螈(Triturus boscai)的研究顯示,重吸收的程度與物種的性成熟年齡幾乎沒有必要的聯系,個體1 齡之前生長期的時間長短才是重吸收程度的決定性因素[36]。
在KL 區域內可能存在著與骨質形成有關的成骨細胞和與骨質重吸收有關的破骨細胞。成骨細胞作用于內骨板的離心生長;破骨細胞則作用于外骨板內側的重吸收。在最外層以骨髓腔為中心呈現輻射狀生長的為外骨板,它的生長使長骨骨壁逐年變粗[22]。外骨板由致密的骨組織構成,顯微觀察長骨橫切片時,可以清晰地看到外骨板上經HE染色后同心排列的LAG[37],它是鑒定年齡的唯一標記。確定KL 后,可以輔助確定LAG1 的位置[38],從第1 條LAG開始到外骨板最外緣所有的LAG 數目,即是實驗動物的年齡[39]。
1.4 骨齡學LAG提示信息
在觀察中發現,每相鄰2 條LAG 之間骨組織的顏色與寬度均不同[40],寬度反映了兩棲動物對應年份的生長情況。2 條LAG 之間距離較寬,提示對應年份環境條件(溫度、食物等)良好,適宜個體生長,骨質積累狀態良好。而環境條件相對較差時,個體的生長速度明顯降低,2 條抑制線之間的寬度就較窄[41]。研究證明兩棲類個體在進入繁殖期前有一個限制性條件,即個體必須到達本物種的最小體長[42],完成變態發育后,均有幾年的快速生長期,因而在高海拔寒冷區域生活的兩棲類較溫暖區域的兩棲類物種需要更長的時間達到最小繁殖體長[43],這種現象在長骨切面有良好的印記,完成變態發育后1~4 年(不同物種有所不同)主要為營養生長期,此時2 條LAG 線間的距離較寬。一旦觀察到2 條LAG 線間的寬度明顯小于其內側2 條LAG 線間的寬度時,則預示著此個體達到了性成熟年齡[41]。究其原因,是由于兩棲類動物個體在營養生長和生殖生長之間的能量投入發生了顯著變化,在前期為了盡早達到性成熟的最小體長,兩棲類動物將大部分能量用于生長和發育,在達到目標后轉而將更多的能量投入到生殖腺及其相關器官的結構和功能的構建和完善中,為進入繁殖期做好充分的準備[44-45]。當然也存在一些特殊情況,如個體達到性成熟后,來年的環境條件優渥,機體在保證生殖投入的前提下,仍能保持較好的生長速度,就會出現性成熟后1 年的2 條LAG 線間寬度大于性成熟當年所形成的LAG 線寬度的情況[46]。
2 骨齡學方法的運用
對兩棲動物骨齡學的研究可以追溯到20 世紀40 年代,兩棲動物第1 次骨齡學技術的成功嘗試是以顱骨為試驗材料完成的[13],1940—1968 年,有學者利用不同的骨塊作為試驗材料探究了兩棲動物的年齡問題。Senning[13]采用副蝶骨測定了泥螈(Necturus maculosus)的個體年齡,Schroeder 等[47]采用翼狀骨對牛蛙(Lithobatus catesbeianus)的年齡進行分析和研究,在后續的研究中分別有學者利用肱骨、股骨、脛腓骨和椎骨等作為試驗材料探究動物的年齡[48],直到Smirina[18]的試驗證明動物趾/指骨同樣能良好地呈現出LAG 的結構特點,指出該試驗的取材方式可以更好地避免對動物個體造成破壞性傷害,此后利用趾/指骨測定動物年齡的方法逐步被認可并廣泛使用。
2003 年,Pancharatna 等[48]對印度大頭蛙(Limnonectes limnocharis)的肱骨、股骨、脛腓骨、橈尺骨和趾骨5 種長骨的LAG 對比分析,結果顯示趾/指骨的LAG 數量基本與其他長骨的LAG 數量保持一致,進一步證明趾/指骨作為兩棲動物骨齡學研究材料的可行性。2008年,美國內政部(U.S.Department of the Interior)和美國地質調查局(U.S.Geological Survey)組織專家編寫使用骨齡學方法研究無尾兩棲動物年齡的方法指南[49],旨在對骨齡學試驗技術方法建立一個標準,促進研究工作的規范化。陳貴英等[50]在2008 年提出,根據骨齡學方法利用動物指骨或趾骨除可鑒定動物年齡外,還可兼顧種群生態學研究中的個體標記和分子生物學研究中的組織取樣,在必要的取材需求下,盡量降低對個體的創傷以達到對兩棲動物資源的保護。對近年來的研究結果統計,顯示現存的8 146種兩棲動物中,只有266種(3.3%)進行了骨齡學研究,其中蠑螈56 種、無尾兩棲類208種,溫帶物種多于熱帶物種[51]。由此可見,兩棲類種群的年齡結構信息仍存在嚴重不足,對全面掌握兩棲類種群現狀、發展信息及物種保護極為不利。
3 骨齡學LAG計數
骨齡學方法雖然快速、簡便,但在年齡的確定過程中依舊容易出現誤差,需要掌握形態學方面的知識來避免年齡鑒定過程中出現的錯誤。
3.1 選取觀察部位
Rozenblut 等[22]通過對3 種歐洲水蛙(Rana lessonae,R.ridibunda,R.esculenta)長骨分化與發育的研究,發現在蝌蚪40~44 期時,軟骨原始細胞最外層的細胞分化成外骨膜,開始逐漸形成外骨板(外骨膜骨);從41 期開始,長骨骨干中央肥大的軟骨細胞退化,骨髓腔逐漸形成;到46 期,骨髓腔向兩端擴大至靠近骨骺處,最終形成1 個沙漏型的長管狀。因此在做長骨切片觀察LAG 時,首選長骨近中部髓腔較小的部位切片,以便觀察到較為完整的外骨板結構,為準確計數LAG 提供保障[22]。尤其在長骨骨干中部,長度為整塊骨長度的25%~35%處,可得到LAG的最大數值[52]。
3.2 附加線
附加線即所謂的假線,假線的形成是由于個體受傷或遇到短暫的氣候劇烈變化或食物缺乏而在長骨切面上的反映,一般假線僅在局部出現,未見其形成閉合環狀結構,且線條結構不清晰[53]。它雖然代表動物在某個生長期的停滯生長卻不代表年周期的生長停滯[54],計數時很容易與LAG區分。
3.3 雙重線
有些兩棲動物以夏眠的方式度過酷熱缺水的夏季,此時在長骨切面上將出現2 條清晰且非常靠近的類似LAG 的結構,稱之為“雙重線”[54]。在計數時,通常把這種情況記為1條LAG。
3.4 外側LAG判斷
在觀察一些老年個體的LAG 時,由于生長緩慢致使2 個年周期的LAG 距離極為接近,若同時出現假線將更加難以區分[39],此時需要有高質量的組織學切片,在高倍鏡下還可較清晰地辨認每條LAG,對計數結果影響不大[55]。如果是在生長期后期或冬眠前期捕獲的個體,可觀察到在外骨板的最外側有一淺染的骨質環,此環是當年生長期產生的骨質,由于沒有冬眠期產生的致密骨質的“襯托”,其尚未形成清晰的LAG,因此,計數骨齡時需要將其計算在內,若忽略此環則會低估個體年齡。
4 結論
出于對兩棲動物保護的迫切需要,骨齡學領域的研究越來越活躍,除了對生長停滯具有明顯周期性變化的溫帶兩棲動物,如大蟾蜍(Bufo bufo)[56]、美洲林蛙(Rana sylvatica)[57]和歐洲林蛙(R.temporaria)[42]等開展研究外,對代謝功能變化不明顯的熱帶和亞熱帶兩棲動物年齡的研究也逐漸增多[58-59],一部分研究表明骨齡學方法可以應用于沒有明顯四季變化的熱帶和亞熱帶物種,但也有一部分熱帶兩棲動物骨骼沒有明顯的生長痕跡。在未來的研究中,挖掘出更多骨齡學研究信息,是值得期待和努力的。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
文苑(2018年22期)2018-11-19 02:54:14
汽車觀察(2018年10期)2018-11-06 07:05:26
