動物DNA甲基化研究現狀
2023-02-13 02:20:50麻錦楠
四川動物 2023年1期
關鍵詞:小鼠
麻錦楠
(云南師范大學,昆明650092)
表觀遺傳修飾主要包括DNA甲基化、組蛋白修飾、RNA修飾和非編碼RNA等。DNA甲基化作為一種被廣泛研究的表觀遺傳修飾方式,是動物和其他真核生物共享的DNA修飾方式。DNA甲基化是指甲基基團在甲基轉移酶的作用下,結合到基因組DNA分子上的生物學過程。DNA甲基化能引起染色質結構、DNA構象、DNA穩定性及DNA與蛋白質相互作用方式的改變,從而調控基因表達(Jinet al.,2011);作為重要的基因組水平的化學修飾,參與了動物生長發育中許多關鍵過程。當原始生殖細胞向生殖脊轉變時,首先通過重編程過程進行全基因組的去甲基化,之后再重新建立新的甲基化形式(Smith & Meissner,2013)。在胚胎發育過程中,也發現了大規模DNA甲基化遺傳修飾的改變(Giffordet al.,2013;Xieet al.,2013;Wanget al.,2014)。此外,嬰兒的發育過程同樣存在DNA甲基化的動態改變,并且DNA甲基化的改變還會受到年齡、疾病、營養等的影響(Chenet al.,2018;Matherset al.,2018;Zhouet al.,2020;He,2022)。本文結合動物DNA甲基化研究文獻,綜述了DNA甲基化特征、DNA甲基化與基因表達調控、DNA甲基化的生物學意義,以及DNA甲基化檢測技術等方面的研究進展。
1 DNA甲基化特征
DNA的4種堿基中,胞嘧啶和腺嘌呤都可以被甲基化,形成5mC和6mA的修飾。胞嘧啶甲基化在原核及真核生物中均廣泛存在,但腺嘌呤甲基化主要存在于植物和細菌中(Yi,2017)。細菌中DNA甲基化可以將細菌自身基因組與噬菌體基因組區分開,噬菌體的DNA得以被宿主的限制性酶切割(Chinnusamy & Zhu,2009)。哺乳動物中,CG類型基序最常見,且脊椎動物體細胞中60%~80%的 CpG 位點都是高甲基化的(Schübeler,2015)。但在哺乳動物某些組織(胚胎干細胞、造血干細胞和神經細胞)中,非CG類型的甲基化占比更高(Ramsahoyeet al.,2000)。在植物中,DNA高甲基化作用發生在3種甲基化基序中,包括CG、CHG和CHH(H代表A、C或T),且大多數植物甲基化區域與非甲基化區域是交替出現的(Schübeler,2015)。DNA甲基化水平受生長發育和環境適應等因素的影響,處于動態變化的過程中。DNA甲基化變化可劃分為3個階段:建立、維持和去除。哺乳動物中,甲基化位點的建立在甲基轉移酶3的調控下進行(Chen & Li,2004),發生部位通常是轉錄活性較弱的啟動子區和轉錄活性較強的基因體區。這些區域往往包含了大量H3K4me3(Piunti & Shilatifard,2016)。甲基轉移酶1可維持DNA復制過程CpG位點的對稱性,避免由于復制造成的甲基化位點缺失。之前被認為是DNA甲基轉移酶的DNMT2已被證實為一種tRNA甲基轉移酶,并已更名為tRNA天冬氨酸甲基轉移酶1(Gollet al.,2006)。此外,DNA甲基化是一種動態修飾過程,不僅因為其能通過酶來調控甲基化與去甲基化的作用,甲基化胞嘧啶還能自發地脫氨,導致基因位點突變頻率的增加。5-甲基胞嘧啶在自發脫氨作用下轉化為胸腺嘧啶,CpG二核苷酸轉化為TpG二核苷酸,而未甲基化的胞嘧啶在自發脫氨作用下產生的尿嘧啶會很快被細胞識別并修復(Landeret al.,2001)。實際上,在進化過程中,許多甲基化CpG位點脫氨并穩定保存,這也解釋了人類基因組中甲基化頻率低于預期值的原因。
2 DNA甲基化與基因表達調控
基因組上不同區域的甲基化對基因表達具有不同的調控作用。哺乳動物中存在富含CpG的區域,被稱為CpG島,其相對較小,平均約1 000 bp。人類和小鼠Mus musculu中30%~40%的CpG島在正常組織中是甲基化的(Yi,2017)。人和小鼠的基因組中,絕大多數的CpG島位于基因的啟動子區,少數位于基因體區的可作為可替代啟動子調節基因表達(Illingworthet al.,2010)。目前CpG島的定義不統一,不同研究者對CpG島的選取有一定差異,因此很多研究都將關注點集中于CpG島最多的啟動子區。啟動子區高密度的甲基化可導致基因的沉默。CpG位點的甲基化能通過2種作用機制影響基因的表達。首先,CpG區的甲基化通過抑制轉錄因子與核苷酸的結合,影響基因的表達調控。6 bp區域中只要存在單個位點的甲基化修飾就能抑制轉錄因子的結合(Watt & Molloy,1988)。其次,甲基化的CpG區域能被含有甲基-CpG結合結構域的蛋白質家族識別。這些蛋白質家族通過不同的domain識別甲基化位點,招募組蛋白脫乙酰激酶HDACs和其他一些染色質重塑蛋白,改變組蛋白的結構,從而形成緊密閉合無活性的染色質,抑制轉錄的過程(Jones & Laird,1999)。越來越多的研究也將啟動子區甲基化水平與基因表達進行聯合分析,研究甲基化在介導生物組織器官發育(Waterlandet al.,2009;Liet al.,2012;Eharaet al.,2015;Maet al.,2022)、疾病發生(Br?gelmannet al.,2021)等過程中的重要作用。
目前大多數研究已證實基因啟動子區域的甲基化會抑制基因的表達,但是除啟動子區域外的其他區域,如基因體區的甲基化功能還是未知的。Brenet等(2011)發現哺乳動物細胞系中首位外顯子的甲基化與基因表達呈負相關性,且首位外顯子的調控作用比啟動子區更顯著。首位內含子甲基化對基因表達同樣存在調控作用:首位內含子與TSS位置較近,其甲基化通過影響轉錄起始位點來影響基因轉錄(Hartonoet al.,2015)。首位內含子的甲基化與基因表達相關性的實驗已經在包括癌癥細胞、胎兒及成年人組織、小鼠CD4+細胞、精神分裂癥患者的白細胞等中進行了驗證(Anastasiadiet al.,2018)。基因體甲基化除有部分調控基因表達的功能外,可能還在調控轉錄的準確性上起重要作用。Neri等(2017)在小鼠胚胎干細胞的研究中發現,DNA甲基轉移酶Dnmt3b介導的甲基化過程可防止生物體內異常轉錄事件的發生,確保轉錄過程的準確性。如果Dnmt3a和Dnmt3b甲基化轉移酶被抑制,胚胎會因轉錄因子的異常表達導致發育畸形而終止妊娠(Kinoshitaet al.,2021)。此外,位于外顯子的DNA甲基化區可通過招募多功能蛋白MeCP2增強HDAC對外顯子的識別,維持局部組蛋白低乙酰化,從而調節RNA的可變剪切(Maunakeaet al.,2013)。
3 DNA甲基化的生物學意義
DNA甲基化的改變與遺傳水平的改變均反映了生物對外界環境的適應機制,但DNA甲基化改變機制的靈活性更高,且具可逆性(Norouzitallabet al.,2018)。DNA甲基化作用在生殖細胞發育階段就已經出現,當原始生殖細胞向配子轉變時,親本特有的表觀遺傳印記會被清除,之后性別特異性DNA甲基化印記又在雄性和雌性配子體發育過程中重新建立(Sasaki & Matsui,2008)。而在由受精卵向早期胚胎的發育中,同樣又循環了去甲基化與甲基化重構的過程(Smallwood & Kelsey,2012)。在胚胎發育過程中,母體環境(飲食結構、生存壓力、生活習慣、行為方式等)會對胚胎發育和表觀遺傳信息造成影響。當母體受到不良因素誘導后,會造成新生兒體質量的改變,并可能導致后代罹患高血壓、冠心病、骨質疏松、糖尿病等(Relton& Smith,2010)。此外,母體適應新環境的能力同樣也決定了胚胎對環境變化的可塑性,這樣的可塑性也能在種系間穩定地遺傳(Devanapallyet al.,2015)。產后器官發育過程也同樣受到甲基化調控。剛出生的幼體肝臟就能通過脂肪酸β氧化過程從母乳中攝取能量,但其中的機制卻鮮為人知。Ehara等(2015)通過對出生前18.5 d和出生后2 d、16 d、28 d的小鼠肝臟組織甲基化情況進行比較分析,發現脂肪酸β氧化相關基因在出生后存在低甲基化高表達的現象,而這些低甲基化基因啟動子區預測到許多氧化物酶體增殖劑激活受體(PPAR)結合基序。其中,PPARα受體在調控出生后小鼠肝臟脂肪酸β氧化相關基因的去甲基化過程中扮演了重要角色。Cannon等(2016)的研究也發現,出生后小鼠肝臟發育過程伴隨著甲基化的改變,從E18.5到P20的6個時間點的甲基化組數據顯示,DNA甲基化的改變發生在造血干細胞遷移后,與肝細胞的分化同時進行。除了模式物種外,非模式物種器官發育中甲基化的調控作用也引起了科學家的關注。Yang等(2021)通過繪制長白豬妊娠前33 d至出生后180 d共27個生長發育時間點的骨骼肌全基因組甲基化和轉錄組圖譜,揭示DNA甲基化在調控豬骨骼肌生長發育中的重要作用。Ma等(2022)對大熊貓Ailuropoda melanoleuca未進食組、吃乳組和成年組的肝臟和胰臟進行DNA甲基化組測序,發現發育過程中肝臟和胰臟甲基化水平發生了動態變化,尤其是營養代謝關鍵基因的甲基化形式隨食性改變發生了顯著改變。
除了發育初期和胚胎階段會發生表觀遺傳修飾外,發育完全的個體同樣會受到環境因素引起的DNA甲基化修飾,導致表型改變。大量關于脊椎動物和無脊椎動物的研究發現,環境因素與表觀遺傳之間存在相關性,包括環境污染(如無機污染物、農藥污染和抗菌物污染等)、飲食營養、父母行為等(Wang Tet al.,2017;Wang Yet al.,2017;Kelleyet al.,2021)。其中,營養作為最重要的DNA甲基化影響因子,在表觀遺傳標記建立和去除中起重要作用。在人類和小鼠中,高脂飲食導致大量基因啟動子甲基化改變,影響器官發育和功能(Jacobsenet al.,2012;Zhanget al.,2015)。低營養飲食也對甲基化模式產生影響(Yaskolkaet al.,2021)。營養物質可能通過3種方式影響DNA甲基化模式(圖1):(1)改變甲基化過程涉及的底物;(2)改變調節葉酸循環中酶的活性;(3)改變DNA甲基化過程中酶的活性(Zhang,2015)。SAM屬于DNA和蛋白質甲基轉移酶的主要供體,通過甲硫氨酸循環過程合成。以上方式可直接或間接影響SAM合成過程,導致體內整體DNA甲基化水平的改變(Niculescu,2012;Kadayifciet al.,2018)。
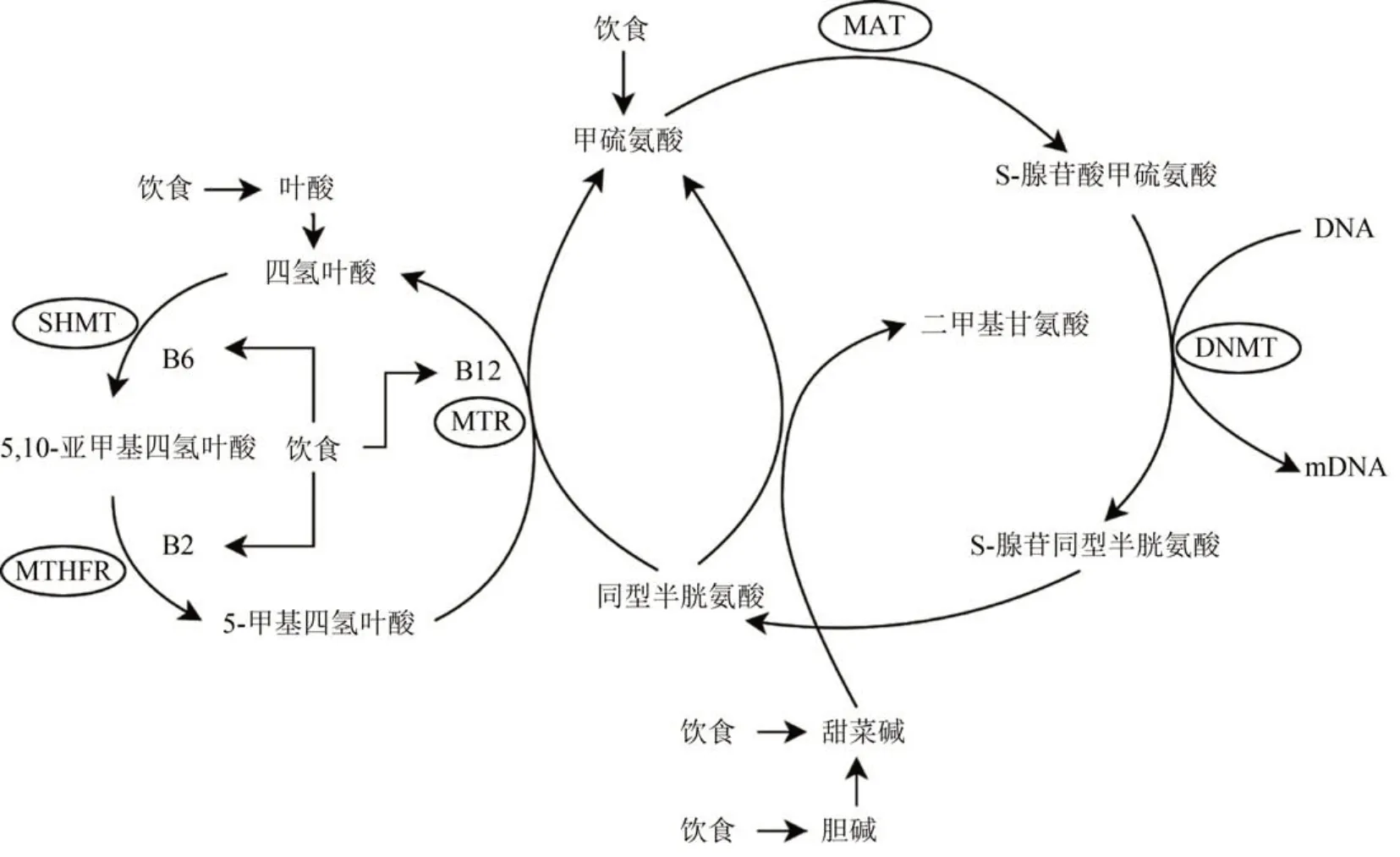
圖1 營養物質影響DNA甲基化的方式(Zhang, 2015)Fig. 1 Possible nutritional ways that nutrition influences DNA methylation(Zhang, 2015)
環境誘導的DNA甲基化修飾通過改變基因表達,影響個體及其后代的表型,為后代適應新環境提供了有效保障。Norouzitallab等(2014)對孤雌生殖的雌性鹵蟲Artemia進行非致死性熱休克刺激后,所有親本個體產生了一系列響應機制,包括Hsp70表達水平提高,對熱應激的耐受性增加及對致病性弧菌的抗病性增強等。值得注意的是,這些表型能夠穩定、連續遺傳到子三代。子代鹵蟲表型的改變與DNA甲基化有關,證實了表觀遺傳修飾產生的適應性表型的可遺傳性。蚜蟲Acyrthosiphon pisum的跨代實驗也得到相似的結果,當蚜蟲飼養環境中存在空間和食物資源競爭時,將誘發蚜蟲從無翅個體轉變為有翅個體,這樣的轉變往往發生在個體發育的早期階段,并與蚜蟲幼激素結合蛋白的甲基化有關(Walshet al.,2010)。在哺乳動物的研究中同樣發現了DNA甲基化的可遺傳性,Li等(2011)對連續6代的小鼠進行甲基飲食喂養后,甲基供體導致了小鼠基因組數千個位點甲基化的隨機改變。隨著世代的增加,個體間表觀遺傳變異也隨之增加。此外,小鼠不同世代受甲基化影響較大的基因主要參與了基因表達、器官形成和細胞發育的過程。總之,由環境介導的、表觀遺傳修飾產生的表型改變可能獨立于自然選擇以外,這些表觀遺傳修飾可以讓生物在短期環境的改變下快速進行適應性轉變。而長期暴露于某一環境中的物種,其體內會產生持久的表觀遺傳變化,這些變化又通過增加5-甲基胞嘧啶的變異度促進可遺傳突變的發生。
DNA甲基化修飾涉及到大量的生物學過程,異常的甲基化還會導致生物發育異常,甚至疾病的發生。腫瘤抑制基因(TSG)和癌基因的異常DNA甲基化在癌癥的發病機制中起關鍵作用。多種癌癥中,普遍存在TSG的超甲基化現象,包括P16/INK4基因(Parrellaet al.,2004)、CADM基因(Heet al.,2013)、RASSF1A基 因(Huanget al.,2016)等。TSG啟動子區高甲基化后會導致細胞凋亡,細胞粘附性及血管生成過程的失調,誘導癌癥的發生。此外,癌癥中除了異常高甲基化修飾外,也存在整體去甲基化及原癌基因低甲基化的情況。甲基化水平降低會引發基因組的不穩定性,激活重復序列元件,導致基因組印記缺失及不可控的細胞增殖現象(Esteller,2007)。全基因組甲基化測序發現,原癌基因的高表達與CpG島的低甲基化有關。Zhao等(2020)關于前列腺癌的研究發現,原癌基因附近轉錄因子和增強子結合位點的低甲基化,是驅動原癌基因表達的主要因素。Suzuki等(2014)研究發現,基因組的CpG位點甲基化的模式與癌癥診斷和預后相關,某些基因簇中CpG島的甲基化模式已被用于腫瘤的預后和分類。
4 DNA甲基化的檢測
在過去10年中,已開發許多DNA甲基化檢測技術(Laird,2010)。部分方法集中于檢測DNA甲基化區域,如甲基化DNA免疫沉淀法將基因組隨機打斷為300~600 bp的片段,使用抗5mC的抗體孵育后富集出甲基化片段,設計特異性引物檢測目的基因甲基化水平(Jacintoet al.,2008);甲基化敏感限制性內切酶法利用甲基敏感性內切酶(HpaII、Hin6I和AciI)切割出含有甲基化CpG位點的序列,通過電泳分離對甲基化片段進行分析(Maunakeaet al.,2010);其他方法則集中對單一位點甲基化進行檢測,主要步驟:用重亞硫酸鹽將DNA中的C轉化為U,經PCR后轉化為T,而未甲基化的C位點不會發生變化,仍以C的形式存在(Frommeret al.,1992)。通過芯片技術及二代高通量測序技術即可將基因組所有C位點進行測序。此外,一些單分子測序技術采用合成寡核苷酸探針來直接鑒定甲基化胞嘧啶(Yanget al.,2015)。
在這些方法中,有2種方法被廣泛用于評估單核胞嘧啶DNA甲基化水平,分別是簡化重亞硫酸鹽測序(RRBS)(Meissneret al.,2005)和全基因組重亞硫酸氫鹽測序(WGBS,又稱BS-seq、methyl-seq或methylC-seq)(Cokuset al.,2008)。這2種方法均需要將提取的DNA進行重亞硫酸鹽處理。WGBS在基因組的覆蓋范圍更廣,能用于檢測每個C位點的甲基化水平,為了獲得可信度高的甲基化值,該方法需保證每個位點的reads覆蓋度達到5~10×,所以價格更高。RRBS需要限制性酶MspI消化DNA,并富集含CCGG位點的50~300 bp的片段。該方法只需要對基因組部分位置進行檢測,其測序覆蓋度較低。這些新技術的出現使更好、更全面認識DNA甲基化修飾成為可能,幫助我們從少量基因位點的研究擴大到基因組范圍。
5 展望
動物DNA甲基化研究涉及生長、發育、環境、營養等多個方面。DNA甲基化通過調控基因表達,影響真核細胞的發育、分化等過程,并在癌癥、腫瘤等疾病發展中起到重要作用。越來越多的研究使用DNA甲基化作為生物標志物,用于疾病的識別和診斷、動物生長性狀標記、酮體性狀標記等。但目前的研究仍面臨許多挑戰,如某些疾病樣本及動物組織樣本獲取困難;由于DNA甲基化的動態性及相對不穩定性,難以建立穩定的DNA甲基化參照模型等。隨著后期DNA全基因組甲基化生物信息庫的建立及DNA甲基化檢測技術的發展,這些問題將會逐步得到解決。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
