產漆酶食用菌的篩選及食用菌漆酶降解黃曲霉毒素B1的研究進展
2023-02-12 03:03:06婁海偉楊創明牛永武田雙起趙仁勇
食品科學 2023年1期
婁海偉,楊創明,趙 玉,牛永武,田雙起,*,趙仁勇,*
(1.河南工業大學 小麥和玉米深加工國家工程研究中心,河南 鄭州 450001;2.河南工業大學糧油食品學院,河南 鄭州 450001)
黃曲霉毒素(aflatoxin,AF)是一類主要由黃曲霉(Aspergillus flavus)和寄生曲霉(A.parasiticus)產生的次級代謝產物[1]。目前已發現的AF有20余種,該類化合物均具有一個二呋喃環和一個氧雜萘鄰酮(香豆素),常見的AF有AFB1、AFB2、AFG1、AFG2、AFM1、AFM2等,其毒性由強到弱依次為AFB1、AFM1、AFG1、AFB2、AFM2、AFG2[2]。由于AFB1的毒性強、污染性廣、致癌性強,已被世界衛生組織國際癌癥研究機構列為IA類致癌物[3],AFB1污染食品會對人類和牲畜健康造成極大的危害[4]。因此,各個國家和組織對食品和飼料中AFB1含量有明確的限量要求,如我國GB 2761-2017《食品安全國家標準 食品中真菌毒素限量》和GB 13078-2017《飼料衛生標準》分別對食品和飼料原料、飼料產品中AFB1含量有明確的限量要求。不可控的自然環境因素(如高溫、降雨等)以及谷物收獲后不適宜的貯藏或加工條件都會引起谷物中霉菌大量繁殖并產生真菌毒素,最終導致谷物中AFB1含量超出限量標準,谷物食用價值降低或喪失,造成重大經濟損失,因此研究人員一直致力于研究減少或消除谷物中的AFB1的方法。目前,消減谷物中AFB1的方法主要有如下3 類[5]:1)物理法,包括高溫、輻照等方法,對AFB1消減有一定效果,但會導致谷物營養成分的損失;2)化學法,包括堿處理、氧化劑處理等,對AFB1的脫毒效果好,但會造成二次污染;3)生物法,該方法是利用微生物或酶制劑降解AFB1,是近年來的研究熱點,該方法對底物AFB1的專一性強、反應條件溫和、降解效果穩定且效率高,極具發展潛力[6]。漆酶(EC 1.10.3.2)[7-9]、錳過氧化物酶[10-11]、葡萄糖氧化酶[12]等均可以降解AFB1,其中漆酶是一種無污染的綠色環保型催化劑[13],因此利用漆酶降解AFB1引起了研究人員的廣泛關注。
漆酶是一種廣泛存在于植物(如漆樹、棉花、馬鈴薯等)、真菌(如云芝、平菇、杏鮑菇等)、細菌(如芽孢桿菌、假單胞桿菌等)和昆蟲中的酚類氧化酶[14],在食品工業、纖維改性、生物合成、環境保護、印染、造紙、生物傳感器、生物能源、免疫監測、飼料等領域已有廣泛的應用[13,15]。相對于植物漆酶和細菌漆酶,真菌漆酶的相關研究報道較多[16-17],而食用菌是真菌漆酶的主要來源之一[18]。無毒無害且可食用的產漆酶食用菌可作為食品或飼料領域降解AFB1的重要微生物。基于此,本文從美國國家生物技術信息中心(National Center for Biotechnology Information,NCBI)數據庫(www.ncbi.nlm.nih.gov)和Uniprot數據庫(Unified Protein Database,www.uniprot.org)中篩選具有漆酶基因序列或氨基酸序列的食用菌,總結已報道的產漆酶食用菌,并介紹食用菌漆酶的結構、理化性質及其對AFB1的降解作用,以期為食用菌漆酶安全、高效、綠色降解AFB1提供參考。
1 產漆酶食用菌的篩選
1.1 基于生物信息學數據庫和文獻報道篩選產漆酶的食用菌
食用菌種類繁多,且不同菌種之間性狀差異很大,篩選產漆酶的菌種是一項任務非常繁重的工作。因此,基于生物信息學數據庫NCBI和Uniprot,能夠快速篩選到具有漆酶基因序列或氨基酸序列的食用菌,基于NCBI數據庫檢索具有漆酶基因序列食用菌的路徑為NCBI數據庫→選擇Nucleotide→輸入laccase并點擊Search→選擇“真核生物(eukaryotes)”條目下的“真菌(fungi)”,可獲得13858 條目(items);基于Uniprot數據庫檢索具有漆酶氨基酸序列食用菌的路徑為Uniprot數據庫→選擇UniProtKB→輸入laccase并點擊Search→獲得16885 條目,然后從上述獲得的條目中篩選出食用菌(表1)。此外,本文亦對已報道的產漆酶食用菌進行總結(表1)。從NCBI數據庫篩選出97 種具有漆酶基因序列的食用菌,從Uniprot數據庫篩選出93 種具有漆酶氨基酸序列的食用菌,已報道的能夠產漆酶食用菌有75 種,合計篩選出食用菌133 種,這有利于學者們縮小食用菌的篩選范圍或快速、簡便地鎖定產漆酶食用菌。

表1 產漆酶的食用菌和具有漆酶基因序列(或氨基酸序列)的食用菌Table 1 Edible fungi with the ability to secrete laccase and those with laccase gene sequences (or amino acid sequences)
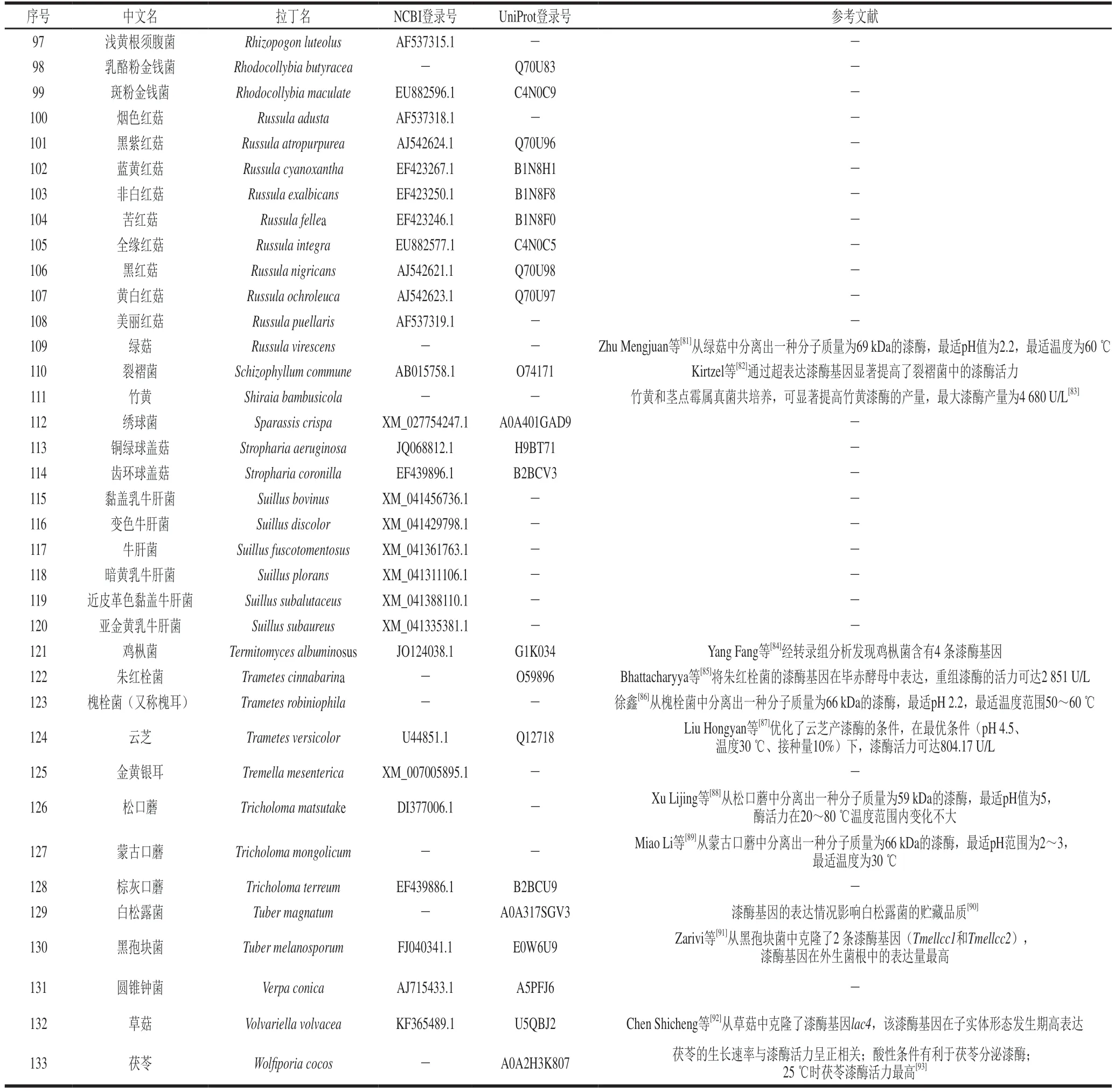
續表1
1.2 產漆酶食用菌的實驗篩選方法
從眾多的食用菌中篩選出產漆酶食用菌是一項繁重而復雜的工作,因此,學者們建立了產漆酶食用菌的實驗篩選方法(初篩和復篩)。即使對于表1中的食用菌,也需要采用如下方法進行驗證。
1.2.1 初篩方法
產漆酶食用菌的初篩方法為平板顯色法。根據培養基中添加底物的不同,平板顯色法劃分類型如表2所示。除了表2中的幾種初篩方法外,也有學者向培養基中添加聯苯胺、2,6-二甲氧基苯酚等試劑進行平板顯色初篩[23,100],但是相比而言,學者們認為愈創木酚平板顯色法產生的氧化帶顏色較明顯、顯色較早、結果較可靠且相關研究報道較多[101]。

表2 檢測食用菌是否產漆酶的平板顯色法Table 2 Features of chromogenic plate methods for detecting for the secretion of laccase from edible fungi
以上方法常依據變色速率、顏色、變色圈直徑、變色圈和菌絲圈直徑的比值來判斷食用菌是否產漆酶和產漆酶的能力。筆者課題組研究(結果尚未發表)表明,采用平板顯色法判斷食用菌是否產漆酶的準確性較高,而通過顏色深淺、比值(變色圈和菌絲圈直徑的比值)判斷食用菌所產漆酶的活力大小有時會存在較大差異,因此,筆者認為平板顯色法適用于判斷食用菌是否產漆酶,不適用于對食用菌所產漆酶的活力進行定量分析。
1.2.2 復篩方法
由于初篩不能確定菌株所產漆酶的活力,因此,需要采用復篩來進一步確定菌株是否產漆酶及所產漆酶的活力。基于食用菌漆酶是胞外酶,在復篩時常將食用菌進行液體深層發酵,然后離心獲得含有漆酶的上清液,對含有漆酶的上清液進行酶活力測定[102]。測定漆酶活力的方法有高效液相色譜法、微量熱法、測O2法、分光光度計法、脈沖激光光聲法等。其中分光光度計法具有操作簡便、設備成本低、測定快速、準確等優點,其應用廣泛,相關研究報道也最多。根據底物的不同,分光光度計法又分為愈創木酚法[103]、ABTS法[23]、丁香醛連氮法[104]等,其中ABTS法、愈創木酚法測定漆酶活力的穩定性和重現性較好,較為常用,如Xu Jianzhong等[33]采用ABTS法測定大杯傘漆酶的活力(701 U/L),Umar等[41]采用愈創木酚法測定白肉靈芝漆酶的活力(6540 U/L)。
2 食用菌漆酶的結構和理化性質
2.1 食用菌漆酶的結構
漆酶是一類含銅離子的多酚氧化酶,食用菌漆酶一般為高度糖基化的胞外分泌的蛋白[105],其穩定性很大程度上是由糖基化貢獻的[106],通常含有4 個銅離子(I型銅離子1 個、II型銅離子1 個、III型銅離子2 個),4 個銅離子分布在漆酶的3 個氧化還原活性中心。T1活性中心(即I型銅離子(單電子受體)所在位點)是漆酶與底物的結合位點,該位點的氧化還原電勢直接影響漆酶的氧化能力[107]。食用菌漆酶在催化底物時,其T1活性中心首先與底物結合,然后I型銅離子(Cu2+)奪取底物電子并轉變為Cu+,電子再經由His-Cys-His三肽區域傳遞給II型銅離子和III型銅離子中心,在此催化過程中,底物被氧化降解,O2被轉化為H2O,無其他副產物產生[108],因此,漆酶被稱為綠色催化劑[13]。
2.2 食用菌漆酶的理化性質
不同來源漆酶的理化性質存在一定的差異,漆酶的理化性質主要由其蛋白編碼基因決定。不同食用菌漆酶的氨基酸序列(或核苷酸序列)并不相同,而且食用菌基因組中通常存在多條漆酶編碼基因,如平菇有12 條漆酶編碼基因[109]、香菇有14 條漆酶編碼基因[110]、真姬菇有10 條漆酶編碼基因[111]、黑木耳有7 條漆酶編碼基因[29]、草菇有11 條漆酶編碼基因[112],這些差別使食用菌漆酶具有不同的理化性質。
2.2.1 pH值
不同食用菌漆酶常具有不同的最適pH值,但通常最適pH值偏酸性,如松杉靈芝漆酶(分子質量52 kDa)的最適pH值為4,在此pH值條件下,松杉靈芝漆酶具有良好的穩定性,當pH值降低或升高時,其穩定性逐漸下降[113];巴爾喀什蘑菇漆酶的最適pH值為2.2,在pH值為7~9時,其活力基本喪失[19];一種中國美味蘑菇漆酶的最適pH值為5.0[23]。綜上,食用菌漆酶的最適pH值通常在2~6之間,偏離此pH值范圍,漆酶活力會顯著降低。如果欲在堿性條件下使用食用菌漆酶,可考慮采用基因工程技術改變漆酶的最適pH值或pH值適用范圍,如采用定點突變技術改變漆酶的氨基酸序列,篩選獲得在堿性條件下具有良好漆酶活力的基因工程改造酶,然后通過異源表達獲得大量改造后的漆酶,以滿足應用需求。
2.2.2 溫度
食用菌漆酶的最適溫度一般不超過60 ℃,這主要與其結構和來源有關,如花臉香蘑漆酶有3 個保守結構域(Cu-oxidase I、Cu-oxidase II和Cu-oxidase III),其最適溫度為60 ℃,當溫度低于60 ℃時,其活力隨著溫度的升高而逐漸增大,當溫度高于60 ℃時,其活力隨著溫度的升高而急劇減小[55];巴爾喀什蘑菇漆酶不耐高溫,最適溫度為40 ℃,在4~50 ℃之間相對穩定,當溫度高于50 ℃時,其酶活力急劇下降[19]。來自同一食用菌的不同基因編碼漆酶,其最適溫度也存在不一樣的情況,如白靈側耳的漆酶Lac1(分子質量68 kDa)和Lac3(分子質量51 kDa)的最適溫度為50~60 ℃,漆酶Lac2(分子質量64 kDa)的最適溫度為60 ℃,在熱穩定性方面,漆酶Lac2較Lac1和Lac3更穩定[114]。食用菌漆酶的不耐高溫特性使其應用范圍受到一定的限制,如果需要耐高溫的漆酶,可采用基因工程技術(如定點突變和異源表達)改造漆酶的氨基酸序列以構建耐高溫漆酶。
2.2.3 金屬離子
金屬離子對食用菌漆酶的活性有顯著的影響,對漆酶活力起促進作用還是抑制作用,這與金屬離子的種類和濃度有關,如Fe3+對平菇漆酶的活力起抑制作用,隨著Fe3+濃度的增加,對漆酶活力的抑制作用增強,而Cu2+對平菇漆酶活力的影響主要與其濃度有關,當Cu2+濃度為1 mmol/L時,對漆酶的活力起促進作用,當Cu2+濃度為5 mmol/L時,漆酶的活力受到抑制[115];Na+、K+、Mg2+對花臉香蘑漆酶的活力無顯著影響,Zn2+、Al3+抑制花臉香蘑漆酶的活力且隨離子濃度的增加抑制作用顯著增強,Mn2+在低濃度時促進花臉香蘑漆酶活力,在高濃度時抑制漆酶活力[55];Hg2+顯著抑制口蘑漆酶的活力[89],Hg2+和Cd2+亦顯著抑制中國美味蘑菇漆酶的活力[23]。因此,在使用食用菌漆酶時要嚴格控制或調整金屬離子的種類和濃度,使漆酶的活力得到充分發揮。
3 食用菌漆酶對AFB1的降解作用
3.1 食用菌漆酶的作用
漆酶在食用菌生長過程中主要起到如下5 個方面作用:1)參與食用菌色素的形成[13];2)降解木質素,為食用菌生長發育提供營養[116];3)參與食用菌的呼吸作用并承擔傳遞電子的作用,促進食用菌的生長發育[117];4)催化木質素降解產生酚類或醌類物質,酚類和醌類物質的毒性能夠抑制雜菌的生長[118];5)調控食用菌原基的分化,改善食用菌產量和品質[119-120]。
3.2 漆酶的底物
漆酶的作用底物十分廣泛,主要包括:1)酚類及其衍生物[108];2)芳胺類及其衍生物(芳胺類化合物與酚類化合物的結構相似)[121];3)羥酸及其衍生物[122];4)生物色素和甾體激素[123-124];5)金屬有機化合物[125];6)一些非酚類化合物[126]。漆酶催化底物氧化需要滿足2 個條件,首先底物能夠進入漆酶的活性中心并與漆酶結合;其次底物和漆酶之間要存在氧化還原電勢差。如果底物的氧化還原電勢較高或無法進入漆酶的活性中心,導致底物不能被漆酶直接氧化,此時可借助小分子介體實現底物和漆酶之間電子的傳遞,以間接實現底物的氧化,這就是漆酶的介體體系,此體系可擴大漆酶催化的底物范圍[6,127]。
3.3 食用菌漆酶對AFB1的降解
食用菌是一類可以食用或藥用的大型真菌[128],其產生的漆酶也是安全的,因此,采用食用菌或食用菌漆酶來降解谷物中的AFB1是一種安全、高效且極具發展潛力的方法,如果采用食用菌發酵已污染AFB1的谷物,不僅能夠降解AFB1,同時還能豐富谷物的營養成分。目前,已報道的能夠降解AFB1的產漆酶食用菌主要有如下幾種。
3.3.1 平菇漆酶
有關平菇產漆酶的研究報道較多,然而有關平菇漆酶降解AFB1的報道較少。Alberts等[129]研究了不同液體培養基(葡萄糖-麥芽提取物-酵母提取物培養基、基礎鹽培養基、礦物鹽-麥芽提取物培養基、小麥秸稈培養基)對平菇產漆酶的影響,發現平菇在基礎鹽培養基中所產的漆酶活力最高(416.39 U/L),此培養基濾液(含漆酶)對AFB1的降解率可達35.90%。平菇不僅能夠降解谷物中的AFB1,還能降解農業副產物中的AFB1,如采用平菇發酵已污染AFB1的水稻秸稈,平菇菌株MTCC142和菌株GHBBF10對AFB1的降解率分別可達89.14%和91.76%,而且加入Cu2+和Zn2+可有效提高AFB1的降解率;因此,采用平菇發酵動物飼料是一個生態環境友好的降解AFB1的方法[130]。平菇的漆酶活力越高,則降解AFB1的能力越強,即平菇的漆酶活力與其AFB1降解率呈正相關[131]。
3.3.2 云芝漆酶
云芝漆酶能夠直接氧化AFB1,且AFB1降解產物的致突變性、促氧化能力和肝毒性顯著降低[8,132]。何杜鵑等[133]采用雙水相法萃取云芝漆酶,并將萃取的云芝漆酶(80 U/mL)用于降解AFB1,使AFB1的降解率達到了46.32%。Alberts等[129]采用Sigma公司生產的云芝漆酶(1 U/mL)降解AFB1,AFB1的降解率達到了87.34%,遠遠高于何杜鵑等研究中的結果,這可能是酶解條件(酶解溫度、酶解時間和AFB1的濃度)不同所致,Alberts等[129]認為漆酶能夠破壞AFB1分子中呋喃環的雙鍵,從而影響其熒光和致突變性。Liu Yingli等[134]研究發現漆酶與AF的結合能力由強到弱依次為AFB1>AFG2>AFG1>AFB2,結合能力越強,則漆酶降解AF的效率越高,同時,其將云芝漆酶基因在釀酒酵母中進行異源表達,然后采用重組的云芝漆酶分別降解AFB1、AFB2、AFG1和AFG2,降解率分別可達90.33%、74.23%、85.24%和87.58%,最適降解條件為30 ℃、0.1 mol/L磷酸鹽緩沖液、pH 5.7、降解時間48 h。
3.3.3 杏鮑菇漆酶
有關杏鮑菇漆酶降解AFB1的研究報道較少,但杏鮑菇漆酶能夠有效降解AFB1。Brana等[135]從杏鮑菇菌糠中提取漆酶并獲得了漆酶粗提物,在25 ℃條件下,采用漆酶粗提物降解AFB1,分別孵育1、3、7 d,使AFB1的降解率分別達到了50%、75%和90%。介體不僅可提高漆酶對AFB1的降解效率,還可使漆酶同時降解多種真菌毒素,如Loi等[6]采用釀酒酵母表達杏鮑菇漆酶,經純化獲得了重組漆酶,然后采用純化的重組漆酶分別降解AFB1、伏馬毒素B1、赭曲霉毒素A、玉米赤霉烯酮和T-2,在25 ℃條件下降解72 h,上述毒素的降解率分別達到了73%、74%、27%、100%和40%,采用重組漆酶和氧化還原介體可同時降解AFB1和玉米赤霉烯酮,使AFB1和玉米赤霉烯酮的降解率分別達到了86%和100%。杏鮑菇在液體培養基和固體培養基中均可降解AFB1,在含有128 μg/kg AFB1和質量分數25%玉米的生長培養基上栽培杏鮑菇,28 d時AFB1降解率為86%,而且杏鮑菇的生物效率和產量無顯著降低,在杏鮑菇菌絲體和子實體中均未檢測到AFB1及其降解產物,表明培養基中的AFB1并不向杏鮑菇中轉移[136]。
此外,肺形側耳漆酶亦能夠直接氧化AFB1,其對AFB1的直接氧化降解率為23%,添加乙酰丁香酮介體后,AFB1的降解率提高至90%,肺形側耳漆酶不僅能夠降解AFB1,而且還能夠完全降解AFM1[137]。除了上述幾種食用菌,其他產漆酶食用菌降解AFB1的研究鮮有報道。
4 結語
由于自然環境因素,每年存在大量的谷物被AFB1污染,不但造成重大經濟損失,而且嚴重危害人體健康和生態環境,因此,高效降解AFB1已成為亟待解決的問題。目前,降解AFB1的方法有物理法、化學法和生物法,生物法因具有底物特異性強、降解條件溫和等優點,受到學者的廣泛關注。在降解谷物中AFB1時應采用食用安全的微生物,因此,具有食用安全、營養豐富等特點的食用菌可用于降解谷物中的AFB1。前人的研究結果表明,食用菌漆酶能夠高效降解AFB1。基于此,本文首先從生物信息學數據庫(NCBI和Uniprot)中篩選具有漆酶基因序列或氨基酸序列的食用菌,對已報道的能夠產漆酶的食用菌進行了總結,并簡述了產漆酶食用菌的實驗篩選方法;其次,探討了食用菌漆酶的結構和理化性質;最后,闡述了食用菌漆酶的作用、漆酶的作用底物及食用菌漆酶對AFB1降解作用的研究進展,可為食用菌漆酶綠色、安全、高效降解AFB1提供參考,亦可為擴大食用菌漆酶在食品或飼料領域的應用提供思路。
采用食用菌漆酶降解AFB1極具發展潛力,但是目前仍然存在如下幾個亟待解決的問題:1)篩選高產漆酶的食用菌菌株;2)采用基因工程技術構建耐高溫、耐酸或耐堿、酶活力高的工程漆酶以滿足不同領域的需求;3)鑒定AFB1降解產物的結構和分析降解產物的性質,以揭示漆酶對AFB1的降解機理;4)開發食用安全、無環境污染的介體以擴大漆酶作用的底物范圍或提高漆酶的催化效率;5)谷物中存在同時污染多種真菌毒素的情況,需開發可同時高效降解多種真菌毒素的食用菌漆酶。
