油橄欖傳粉昆蟲群落結構特征及多樣性分析
2023-01-13 07:58:56陸歡歡黃思程劉文平朱恒星趙凱旋楊文華黃敦元
生態學報 2022年24期
關鍵詞:物種
陸歡歡,黃思程,2,劉文平,鄧 煜,朱恒星,趙凱旋,魏 瑋,楊文華,黃敦元,*
1 重慶師范大學生命科學學院媒介昆蟲重點實驗室,重慶 401331 2 西藏農牧學院資源與環境學院,林芝 860000 3甘肅省隴南市經濟林研究院油橄欖研究所,隴南 742500 4 重慶市林業科學研究院,重慶 401331
生態系統服務是指人類從生態系統中獲取的各種直接與間接收益的總和,也是自然生態系統及其組成物種得以維持和滿足人類生命活動的環境條件和過程[1]。生態系統服務的形成與維持是人類文明和社會經濟持續發展的根基,其形成過程貫穿了自然環境系統和社會系統,而生物多樣性是影響生態系統功能形成和制約生態系統服務發揮的關鍵因素之一[2—4]。生物多樣性是生物(動物、植物、微生物)與環境形成的生態復合體以及與此相關的各種生態過程的總和,其能夠在從局部到景觀的不同尺度上促進生態系統服務功能的提升,因而保護生物多樣性已成為當前國內外自然資源保護實踐的主要內容之一。
傳粉昆蟲是生物多樣性的組成部分,也是生態系統中傳粉服務的重要媒介,昆蟲傳粉在整個動物傳粉生態服務中占比約90%[5]。以蜜蜂類為代表的傳粉昆蟲在維持陸地生態系統平衡的同時,也為人類帶來直接和間接的利益。例如,世界上87.5%的被子植物、70%的經濟作物以及35%的糧食生產取決于以傳粉昆蟲為主的動物授粉,若傳粉昆蟲減少,將直接影響經濟作物和野生植物的授粉服務功能[6—8]。隨著人類活動的加劇,適宜生境的喪失和碎片化、氣候變化、殺蟲劑的大量施用等因素導致傳粉昆蟲生物多樣性受到了嚴重威脅[9—10],所以開展一定范圍的生物多樣性調查與保護顯得尤為重要,尤其是重點物種的調查保護。
油橄欖(Oleaeuropaea)屬木樨科(Oleaceae)木樨欖屬(Olea)常綠喬木,起源于地中海盆地,是世界四大木本食用油料樹種之一[11]。我國油橄欖種植主要分布于甘肅、云南、四川、重慶等地[12],其中甘肅隴南現有栽植面積達3.33萬hm2,占全國栽植總面積40%以上[13]。當前我國油橄欖產業發展迅速,種植面積逐年增加,但存在坐果率低、果實產量低等問題。因此,開展油橄欖花期傳粉生態學的相關研究,對于提高油橄欖產量和新品種選育等方面均具有十分重要的意義。
油橄欖雌雄同株但自交不親和,屬典型異花授粉[14—15],風媒傳粉是實現油橄欖的異花授粉的主要途徑[15—16]。花粉粒的形態學研究表明油橄欖介于風媒傳粉和蟲媒傳粉之間[17—18],在自然條件下,蟲媒傳粉能夠作為風媒傳粉的有效補充,對增強油橄欖異花授粉具有一定的效果[19—20]。因此,為闡明我國油橄欖傳粉昆蟲種類以及不同樣地間傳粉昆蟲群落的分布規律,本研究開展了我國油橄欖傳粉昆蟲多樣性調查,以期為利用昆蟲輔助授粉解決油橄欖因花粉資源限制而導致坐果率低下的問題提供新的思路。
1 材料與方法
1.1 樣地設置
我國油橄欖適生區分為三大區域:白龍江低山河谷區(甘肅隴南),金沙江干熱河谷區(云南麗江、四川西昌等地),長江三峽低山河谷區(重慶萬州、奉節等地)[21—22]。基于全面性、代表性和可達性原則,本研究在全國油橄欖主栽區域內,選取成片栽培且分布范圍較廣的13個典型樣地,進行了油橄欖花期昆蟲采集,以期全面調查油橄欖傳粉昆蟲多樣性。具體樣地信息見表1。
1.2 昆蟲采集與鑒定
2020年4月—2021年 5月中旬,利用兩年時間完成全國13個樣地油橄欖花期傳粉昆蟲的采集及群落調查。油橄欖花期在傳粉昆蟲活躍時間段(8:00—17:00),對每個調查樣地油橄欖傳粉昆蟲的情況進行初步踏查并根據踏查結果設置5個200 m2采集樣方(每個樣方間隔300 m),每個樣方由1人定點觀測并掃網采集。將采集的傳粉昆蟲獨立分裝于5 mL凍存管帶回實驗室進行形態學鑒定和DNA條形碼鑒定,其中形態學鑒定依據《中國動物志》、《中國經濟昆蟲志》、《浙江蜂類志》等文獻資料[23—25],檢索出所屬類群。對形態鑒定疑難種,借助DNA條形碼COI序列,PCR擴增采用通用引物[26](LCO1490: 5′-GGTCAACAAATCATAAAGA TATTGG- 3′; HCO2198: 5′-TAAACTTCAGGGTGACCAAAAAATCA- 3′),目的片段大小約為700 bp。對 COI序列進行質控剪切后,上傳至生命條形碼數據系統 (http://www.boldsystems.org/index.php/IDS_OpenIdEngine),以相似性≥98%為判斷依據進行鑒定,從而確定物種名稱及分類信息。
1.3 數據分析
1.3.1傳粉昆蟲多樣性分析
利用Excel 2019軟件建立13個樣地油橄欖傳粉昆蟲分布信息數據庫。基于不同科級和不同樣地的角度,分別統計傳粉昆蟲的物種豐富度(Species richness,S)、個體數(Individual number,N)、Berger-Parker優勢度指數(Dominance index,DBP)。使用Excel 2019計算上述指數,計算公式見文獻[27],通過數量等級將物種劃分為優勢類群(DBP≥10%)、 常見類群(10%>DBP≥1%)和稀有類群(DBP<1%)[28]。樣地間油橄欖傳粉昆蟲多樣性分析包括群落豐富度、均勻度以及多樣性3個層次[29]。其中,豐富度包括Chao1指數[30](Chao1 index, Chao1)和Margalef指數[31](Margalef′s richness index,RMar);均勻度包括 Simpson均勻度指數[32](Equitability,Jsim);多樣性包括[33]Shannon多樣性(Shannon′s diversity,Hens)和Simpson多樣性(Simpson′s diversity,Dens)。不同多樣性指數均使用R語言vegan包 estimateR函數、diversity 函數計算。
1.3.2傳粉昆蟲群落相似性分析
通過計算不同樣地間群落組成相似程度或距離以量化傳粉昆蟲群落相似性。為減輕S?rensen 相似性指數可用性的負面影響,本研究中油橄欖傳粉昆蟲群落相似性測度采用基于Chao1豐富度估計量的S?rensen 相似性指數[34](Chao-S?rensen-Est Abundance-based,SS)。使用EstimateS 9.1.0計算SS,且使用corrplot包繪制相似性圖。根據SS與S1-S和為1,轉化計算S1-S。基于S1-S的數據,以中值非權重成對組法(UPGMA)的平均聚合聚類為分類策略,使用R語言執行hclust函數并設定選擇觀測的聚類群數量為4(k=4),從而分別對各樣地種群特征進行定性層次分析。根據定性指數相似性原理劃分樣地間相似程度,當0.75 2020年4月—2021年5月,13個采集樣地共采集油橄欖傳粉昆蟲564只,隸屬于3目13科25屬,其中膜翅目413 只、雙翅目132 只、鞘翅目19 只(表2,附錄1)。優勢類群為蜜蜂科(Apidae)、食蚜蠅科(Syrphidae),其中意大利蜜蜂(Apismelliferaligustica)、中華蜜蜂(Apisceranacerana)和灰帶管蚜蠅(Eristaliscerealis)的數量占優;常見類群有隧蜂科(Halictidae)、金龜科(Scarabaeidae)、胡蜂科(Vespidae)與切葉蜂科(Megachilidae),其中以銅色隧蜂(Seladoniaaeraria)、斯馬蜂(Polistessnelleni)和灰地種蠅(Deliaplatura)等為主;稀有類群包含方頭泥蜂科(Crabronidae)、花蠅科(Anthomyiidae)、擬牛天科(Oedemeridae)、分舌蜂科(Colletidae)、鼻蠅科(Rhiniidae)、麻蠅科(Sarcophagidae)與土蜂科(Scoliidae)。不同樣地的優勢類群均在2—3種(除涼山州冕寧縣樣地外),以蜜蜂科(Apidae)、食蚜蠅科(Syrphidae)或蜜蜂科(Apidae)、隧蜂科(Halictidae)兩種組合常見。除重慶市奉節縣樣地存在2種稀有類群外,其他樣地均不包含稀有類群;常見類群為(2.15±0.42)種,相比于其他樣地,隴南市桑家灣社區樣地和隴南市兩水鎮樣地均出現擬牛天科(Oedemeridae)和金龜科(Scarabaeidae)物種。 表2 油橄欖傳粉昆蟲群落組成 多樣性分析結果表明:油橄欖傳粉昆蟲物種數和個體數最多的是重慶市奉節縣樣地,物種數和個體數最少的是西昌市海濱中路樣地;DBP指數最高的是重慶市奉節縣樣地,最低的是西昌市海濱中路樣地;Jsim指數最低的是隴南市兩水鎮樣地,最高的是西昌市海濱中路樣地;Hens指數和Dens指數最高的是重慶市萬州區樣地,Chao1指數和RMar指數最高的是隴南市兩水鎮樣地;Hens指數、Dens指數、Chao1指數和RMar指數最低的均是西昌市海濱中路樣地(表3)。綜上所述,重慶市奉節縣樣地和萬州區樣地群落多樣性程度較高,西昌市海濱中路樣地群落多樣性程度最低。 表3 不同樣地傳粉昆蟲群落多樣性指標 根據相似性指數的結果(圖1),在不同樣地群落物種組成上,兩兩樣地極為相似個數為16個、中等相似為38個、中等不相似為13個和極不相似為13個。其中,綿陽市三臺縣樣地和西昌市南寧區樣地相似度最高(SS=0.97,P<0.01),而隴南市外納鄉樣地和西昌市海濱中路樣地、西昌市南寧區樣地及綿陽市松埡鎮樣地則沒有相關性。結合聚類群觀察數(k=4),各簇內部均有相似物種,而各簇之間則以物種有無被區分。Ⅰ簇各樣地均含有中華蜜蜂(A.ceranacerana)、灰帶管蚜蠅(E.cerealis)和銅色隧蜂(S.aerarius);Ⅱ簇各樣地均含有中華蜜蜂(A.ceranacerana)和意大利蜜蜂(A.melliferaligustica),而Ⅱ簇樣地間相似性主要受是灰帶管蚜蠅(E.cerealis)、銅色隧蜂(S.aerarius)和中華木蜂(Xylocopasinensis)等物種影響;Ⅲ簇西昌市海濱中路樣地則以中華蜜蜂(A.ceranacerana)和灰地種蠅(D.platura)為主;Ⅳ簇則以灰帶管蚜蠅(E.cerealis)和星花金龜屬(Protaetiasp.)為主要特征。 圖1 不同樣地間的相似性分析及層次聚類分析Fig.1 Similarity analysis and hierarchical cluster analysis between different sampling plotsEP1—13表示13個采集實驗樣地;Ss: 基于Chao1豐富度估計量的Sφrensen相似然指數 油橄欖廣泛分布世界各地,現有種植面積達1000萬hm2,有1200多個品種,其中我國先后從地中海地區引進了150多個品種[15]。油橄欖雌雄同株,但由于大多數品種的花粉管穿過柱頭表面后,其生長受到抑制從而導致成熟的花粉粒不能在柱頭表面萌發,使油橄欖表現出自交不親和現象,而花粉管生長停滯可能是受到程序性細胞死亡的干預,因此油橄欖為典型異花授粉植物[14—15]。此外,油橄欖花期可產生大量花粉,但很少產生花蜜,研究表明油橄欖花粉粒呈橢圓形和三環溝形,其形態介于風媒種和蟲媒種特征之間[17—18],而風媒傳粉的需求是該品種近期進化事件[35]。自然條件下,蟲媒傳粉和風媒傳粉共同作用以增強油橄欖異花授粉效果,而昆蟲傳粉協助植物異花授粉,有利于維持植物的遺傳多樣性,增強后代變異性和生境適應性[5],因而,傳粉昆蟲對解決油橄欖的花粉資源限制與增強油橄欖后代適應性具有重要作用。本研究通過對油橄欖傳粉昆蟲調查發現,優勢類群主要有蜜蜂科、食蚜蠅科和隧蜂科昆蟲,其中數量最占優勢的是中華蜜蜂、意大利蜜蜂、灰帶管蚜蠅和銅色隧蜂。其他研究認為蜜蜂科和食蚜蠅科昆蟲在提高植物坐果率與果實品質方面發揮著重要作用[36—37],余海清等認為大葉金頂杜鵑的訪花昆蟲主要為雙翅目食蚜蠅類和蠅類,膜翅目蜂類[37];肖云麗等結果表明意大利蜜蜂授粉可以顯著提高蘋果、大櫻桃的座果率、維生素C含量[38]。因此,推測中華蜜蜂、意大利蜜蜂和灰帶管蚜蠅等優勢傳粉昆蟲對緩解油橄欖花粉資源限制,提高油橄欖坐果率起到積極作用。 昆蟲群落相似性和物種多樣性是衡量一定地區昆蟲資源豐富程度的客觀指標,但特定樣地昆蟲群落相似性和物種多樣性又受當地生境和人為活動等因素的影響[39]。在昆蟲群落相似性方面,楚雄州永仁縣樣地和麗江市玉龍縣樣地在地理位置上相距較近,其自然背景環境相近可能導致兩樣地群落物種相似,隴南地區3個樣地也如是;西昌市海濱中路樣地和西昌市南寧區樣地、涼山州冕寧縣樣地雖地理位置上相距較近,但由于西昌市海濱中路樣地受到較大人為活動干擾,傳粉昆蟲數量驟減,與其他兩個樣地的傳粉昆蟲群落不相似。因此,形成各樣地間傳粉昆蟲群落相似性的原因可能是區間自然環境相似性越高,昆蟲群落物種越相似。這種推測與其他研究者的結論一致,如陳曉曉等對梵凈山國家級自然保護區葉蟬群落結構特征的研究表明,自然環境的相似是昆蟲群落物種相似的基礎[40];劉高峰和楊茂發認為各樣地土壤甲螨群落的結構與其生境條件密切相關,生境異質性越高,狹布類群越多,類群相似性越低[41]。 在昆蟲物種多樣性方面,西昌市海濱中路樣地、西昌市南寧區樣地和涼山州冕寧縣樣地由于海拔相對較高且種植林區內經常進行雜草雜灌清理導致粉(蜜)源植被覆蓋度較低,尤其是西昌市海濱中路樣地人類活動頻繁,因此這些樣地油橄欖傳粉昆蟲多樣性較低。綿陽市三臺縣樣地、重慶市萬州區樣地和重慶市奉節縣樣地海拔相對較低且在油橄欖林區保留大量野生植被,生境類型多樣且幾乎沒有太多的人類活動,為各類昆蟲的取食、棲息、繁殖提供了良好的自然條件,所以油橄欖傳粉昆蟲多樣性相對較高。蔡東章和王德森的結果也表明由于林地中豐富的植物種類,針闊混交林、闊葉混交林和灌草叢中昆蟲群落多樣性的各項指標均較高[42]。因此影響油橄欖傳粉昆蟲多樣性的因素多樣,但油橄欖種植區域內的植物群落(尤其是昆蟲寄主植物及粉源植物)結構越復雜、生境類型越多樣、人類干擾程度越小,生境質量就越優越,昆蟲多樣性就越高。 為增加油橄欖花期傳粉昆蟲多樣性及群落結構穩定性,可通過減少過度人類干擾,保持油橄欖林區及其周邊林區原有植被的覆蓋,尤其是增加油橄欖同花期蜜(粉)源植物的多樣性。本研究中傳粉昆蟲多樣性較高的重慶市奉節縣和重慶市萬州區等樣地的自然生境,可為傳粉昆蟲多樣性低的西昌市海濱中路和綿陽市松埡鎮等樣地的傳粉昆蟲適生生境改造提供參考。總之,為利用傳粉昆蟲對油橄欖進行授粉,良好的設計規劃和合理的保護措施便于保護和恢復昆蟲類群賴以生存的生境,是保護昆蟲多樣性、保證昆蟲資源可持續發展的關鍵。 附錄1 油橄欖傳粉昆蟲種類和數量 Appendix 1 Species and individual number of pollinators inO.europaea 目名Order科名Family屬名Genus種名Species數量Number鞘翅目擬牛天科(Oedemeridae)雙擬天牛屬(Ditylus)Ditylus sp.3Coleoptera金龜科(Scarabaeidae)星花金龜屬(Protaetia)Protaetia sp.16雙翅目鼻蠅科(Rhiniidae)口鼻蠅屬(Stomorhina)不顯口鼻蠅(S. obsoleta)1Diptera麻蠅科(Sarcophagidae)亞麻蠅屬(Parasarcophaga)結節亞麻蠅(P. tuberosa)1花蠅科(Anthomyiidae)地種蠅屬(Delia)灰地種蠅(D. platura)5食蚜蠅科(Syrphidae)優食蚜蠅屬(Eupeodes)大灰優食蚜蠅(E. corollae)14黑帶食蚜蠅屬(Episyrphus)黑帶食蚜蠅(E. balteatus)2管蚜蠅屬(Eristalis)灰帶管蚜蠅(E. cerealis)94鼓額食蚜蠅屬(Scaeva)高加索鼓額食蚜蠅(S. caucasica)1粗股蚜蠅屬(Syritta)黃環粗股蚜蠅(S. pipiens)1印度粗股蚜蠅(S. indica)4細腹蚜蠅屬(Sphaerophoria)印度細腹食蚜蠅(S. indiana)4寬盾蚜蠅屬(Phytomia)羽芒寬盾蚜蠅(P. zonata)5膜翅目蜜蜂科(Apidae)蜜蜂屬(Apis)意大利蜜蜂(A. )104Hymenoptera中華蜜蜂(A. cerana cerana)182Apis sp.11熊蜂屬(Bombus)短頭熊蜂(B. breviceps)9黑足熊蜂 (B. atripes)1蘆蜂屬(Ceratina)沖繩蘆蜂(C. okinawana)5南方蘆蜂(C. cognata)4擬黃蘆蜂(C. hieroglyphica)2木蜂屬(Xylocopa)中華木蜂(X. sinensis)10竹木蜂(X. nasalis)10隧蜂科(Halictidae)淡脈隧蜂屬(Lasioglossum)紅鐮淡脈隧蜂(L. rubsectum)3Lasioglossum sp.-14Lasioglossum sp.-29近淡脈隧蜂(L. affine)8隧蜂屬(Halictus)銅色隧蜂(Seladonia aeraria)23分舌蜂科(Colletidae)葉舌蜂屬(Hylaeus)H. nobuyukii(無中文名)3切葉蜂科(Megachilidae)壁蜂屬(Osmia)Osmia sp.7胡蜂科(Vespidae)直盾蜾贏屬(Stenodynerus)中華直盾蜾贏(S.chinensis chinensis)3隹盾蜾贏屬(Euodynerus)日本隹盾蜾贏(E. nipanicus)1黃胡蜂屬(Vespula)細黃胡蜂(V. flaviceps)1馬蜂屬(Polistes)斯馬蜂(P. snelleni)5中華馬蜂(P. chinensis)2方頭泥蜂科(Crabronidae)陽完眼泥蜂屬(Solierella)S. cypriaca(無中文名)5土蜂科(Scoliidae)長腹土蜂屬(Campsomeris)金毛長腹土蜂(C. prismatica)12 結果與分析
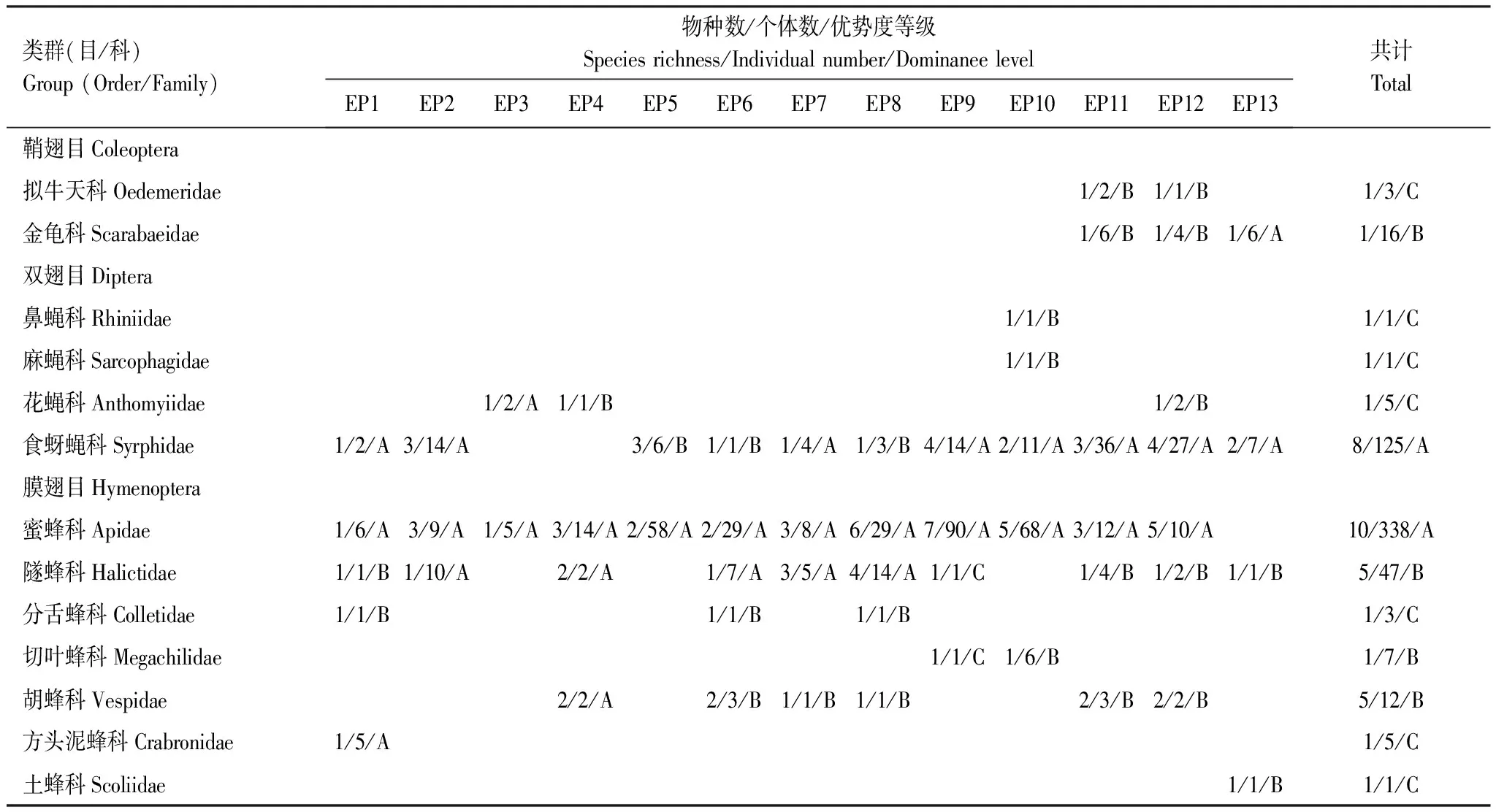
2.1 油橄欖傳粉昆蟲群落組成
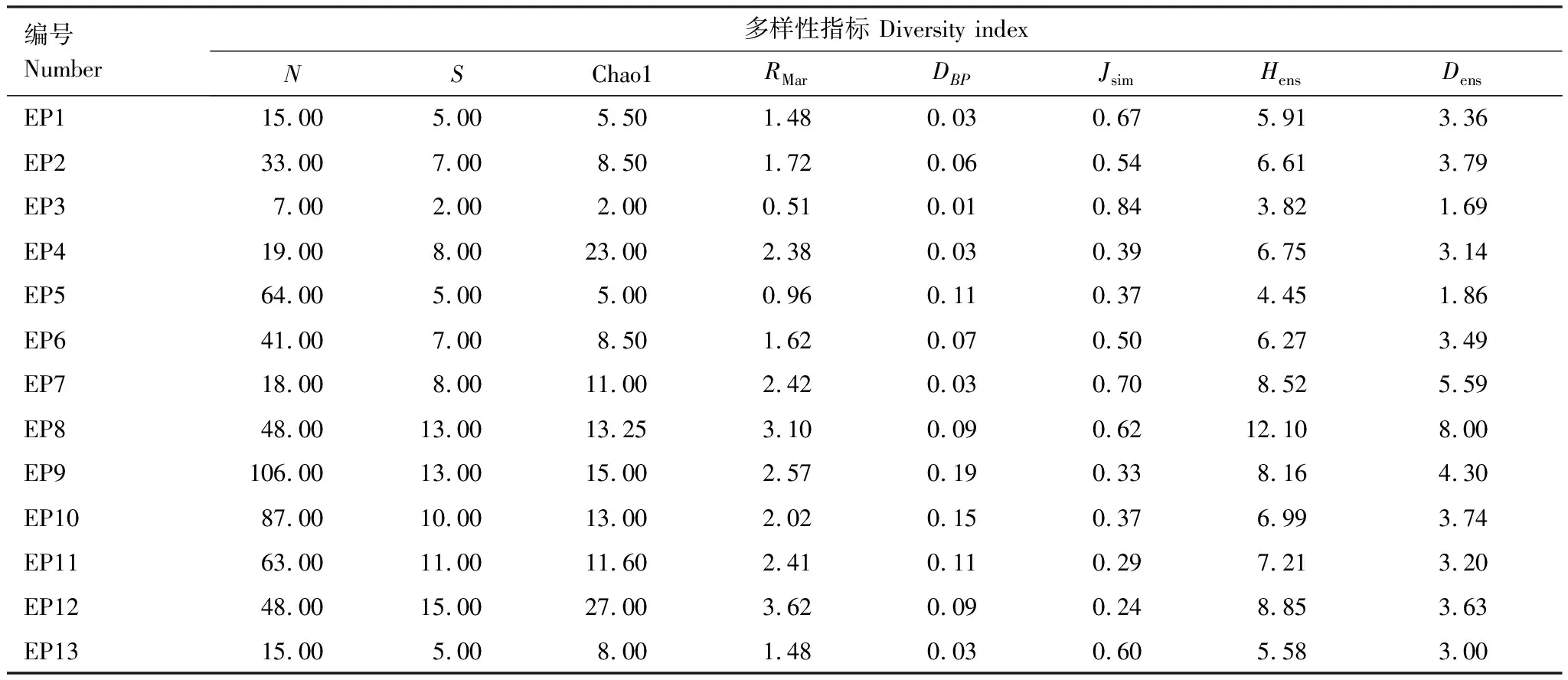
2.2 不同樣地油橄欖傳粉昆蟲群落多樣性比較分析
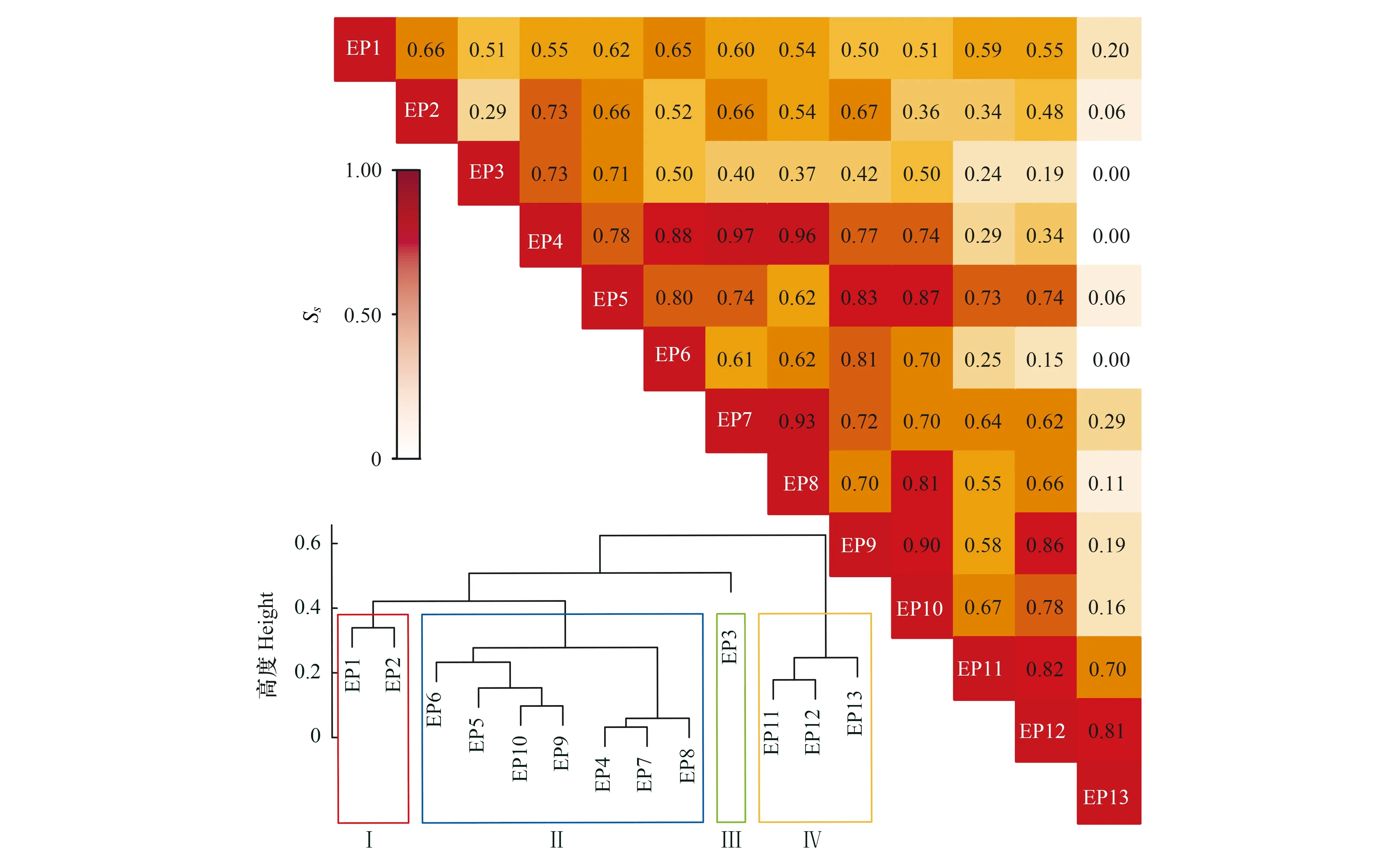
2.3 不同樣地油橄欖傳粉昆蟲群落物種相似性及聚類分析
3 討論與結論
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
