利用CRIS PR/Cas9 技術創制水稻中花11 號dwarf1 突變體
2023-01-13 08:40:44馮彥釗朱慶鋒
廣東農業科學 2022年11期
馮彥釗,朱慶鋒,薛 皦,陳 沛,于 洋
(廣東省農業科學院農業生物基因研究中心/廣東省農作物種質資源保存與利用重點實驗室,廣東 廣州 510640)
【研究意義】隨著人口的迅速增長與全球氣候變化,植物的生長、抗性和品質等受到嚴峻挑戰。分子育種是改良目標性狀、獲得更加優質作物的強力手段。基因編輯是一種利用核酸酶對目標基因進行堿基定點刪除、插入、替換或修飾的技術,利用該技術對特定的基因進行改造可以創制新的種質[1-2]。水稻中花11 號是由中國農業科學院作物科學研究所育成的優質水稻品種,不僅具有良好的產量和抗逆性,同時也是植物分子生物學基礎研究領域最具代表性、使用最廣泛的模式品種之一。DWARF1(D1)是編碼水稻三聚體G 蛋白α 亞基的基因,調控水稻多種生命活動過程[2-3]。然而,在中花11 號中仍缺少對應的d1缺失突變體,給G 蛋白相關通路的遺傳學研究帶來諸多不便。
【前人研究進展】早期的基因編輯技術利用大范圍核酸內切酶誘發雙鏈斷裂,從而產生隨機的堿基插入/缺失或同源重組。然而這種內切酶切割效率較低,且需要長片段的識別序列[4-5]。隨后出現的鋅指核酸內切酶(Zinc Finger Nucleases,ZFN)利用氨基酸模塊與DNA 三連續堿基一一對應的關系進行靶向的DNA 切割,基因編輯效率大大提高[6-8]。不久,人們從侵染植物的黃單孢菌中獲得啟發,利用類轉錄激活效應因子核酸酶(Transcription Activator-like Effector Nucleases,TALEN)作為內切酶進行堿基編輯[9-10]。與之前的ZFN 相比,TALEN 中一個氨基酸模塊只對應一個堿基對,使設計更加簡易[11]。盡管如此,ZFN 和TALEN 均需要針對不同的靶點序列設計合成相應的蛋白,較耗時耗精力。CRISPR/Cas9 則是新一代基因編輯技術,最突出的優點是不依賴于蛋白質識別靶點DNA 序列,而是通過引導RNA(single guide RNA,sgRNA)與靶點DNA 互補配對,Cas9 識別前間隔序列鄰近基序(Protospacer Adjacent Motif,PAM)并行使核酸酶活性對靶基因進行切割[12-14]。至今,基于Cas9 的技術已經被開發出多種形式,可以實現基因激活、熒光定位、單堿基編輯、定向編輯等[15-18]。Cas9 的同家族蛋白Cas12 和Cas13 也相繼被開發用作編輯DNA 和RNA[19-20]。由于該技術的簡易性,CRISPR/Cas9 已成為應用最廣的基因編輯技術,在基礎研究和應用領域創制了大量寶貴材料[21-25],在改良農藝性狀上有廣闊的應用前景[26-27]。
異源三聚體G 蛋白是定位于動植物細胞膜表面的重要信號轉導因子,作用于受體下游。它由α、β 和γ 共3 個亞基組成,在植物中參與激素信號轉導、離子通道調控、免疫應答、細胞分裂等關鍵生理進程。與動物不同的是,植物基因組G 蛋白α 亞基拷貝數非常少,水稻只有一個編碼G 蛋白α 亞基的基因D1。D1基因有許多重要功能,與Gβ 和Gγ 組成復合體,調控水稻對氮素的利用和種子大小[34-38]。此外,D1在稻瘟病的抗性中,與OsRac1共同調控OsMAPK6的蛋白水平,影響下游防御基因的表達[39]。在淹水脅迫環境中,D1在乙烯誘導缺氧薄壁細胞程序性死亡的過程中是必需的[40-42]。同時,D1也是油菜素內酯和赤霉素中信號轉導通路中的一員[43]。運用突變體對D1進行研究已有相當長的歷史,但其突變體遺傳背景局限于日本晴、Taichung65(T65)和Kinmaze 中[2,44]。日本晴背景的d1突變體DK22 第5 個外顯子G1721T 的點突變產生終止密碼,導致較短和較圓的籽粒以及對赤霉素和油菜素內酯的敏感性降低[45]。同為日本晴背景的HO541 有833 bp 缺失,出現短且深綠的葉片,花序變得更緊湊[2]。Kinmaze 背景下的CM1361-1 在D1的第5 個外顯子中發生19 bp 插入,導致節間變短[3,45]。T65 背景d1突變體中的D1基因最后一個外顯子發生2 個堿基缺失,對稻瘟病更易感[3]。中花11 號作為我國水稻基礎研究常用的品種之一,遺傳轉化體系非常成熟,許多基因在此背景中都有相應的突變體,極大促進了水稻基因功能研究。
【本研究切入點】構建有效的中花11 號背景下的d1突變植株,將為水稻基因功能研究提供便利。利用CRISPR/Cas9 技術對中花11 號野生型植株D1基因位點進行編輯,可以在中花11號背景中產生新的D1等位突變,豐富現有的d1突變體種質。【擬解決的關鍵問題】本研究利用CRISPR/Cas9 技術編輯水稻中花11 號的D1基因,篩選及鑒定無抗、有效移碼突變的d1突變植株,并分析d1突變體的表型,以期為進一步研究D1的未知功能提供寶貴材料。
1 材料與方法
1.1 試驗材料
本研究所用水稻材料為中花11 號,為廣東省農業科學院農業生物基因研究中心分子育種技術研究室保存;所用大腸桿菌感受態為Trans1-T1 Phage Resistant Chemically Competent Cell,購自北京全式金生物技術有限公司;所用農桿菌感受態為EHA105,購自上海唯地生物技術有限公司。試驗地點為廣東省農業科學院農業生物基因研究中心,試驗時間為2021 年3 月至2022 年9 月。
1.2 靶向D1 的CRISPR/Cas9 載體構建與遺傳轉化
利用He 等[46]報道的pEntry A 與pRHCas9載體進行構建。基因編輯靶點用CRISPR-P 2.0 進行篩選(http://crispr.hzau.edu.cn/CRISPR2/),選擇的靶點為5'GCTTTGATGAGGCAGAACTT3',靶點在基因組中的位置為625~644(圖1B),合成引物(表1)。以pEntry A 為模板,用ENTRY-F/D1-gRNA-41R、D1-gRNA-41F/ENTRY-R 擴增成兩個片段。將這兩個片段以摩爾比1∶1 混合,再以ENTRY-F/ENTRY-R 引物進行第二輪擴增。反應程序為98 ℃ 3 min;98 ℃10 s、60 ℃ 30 s、68 ℃ 30 s,35 個循環;68 ℃終延伸5 min。第二輪擴增的產物經瓊脂糖凝膠回收后,按照He 等[46]方法連接至pRHCas9 中。構建好的載體轉入大腸桿菌感受態,提取質粒,測序比對后再轉入農桿菌感受態,參照崔瑩等[47]方法侵染水稻胚愈傷組織,獲得轉化植株。
1.3 DNA 提取及PCR 檢測
DNA 提取參考王齊紅等[48]方法進行。基因型檢測的引物為D1-CRISPR-926F71/D1-CRISPR-926R69,利用KOD FX 聚合酶擴增,反應程序為98 ℃ 3 min;98 ℃ 10 s、68 ℃ 60 s,35 個循環;68 ℃終延伸5 min。潮霉素、載體骨架和ACTIN基因檢測引物分別為HPT-F/HPT-R、VEC39-315F74/VEC39-315R75、ACTIN-F/ACTIN-R,序列如表1 所示。反應程序為 98 ℃ 3 min;98 ℃ 10 s,62 ℃ 30 s,68 ℃ 60 s,35 個循環;68 ℃終延伸5 min。
1.4 RNA 提取與qRT-PCR 檢測
按照Simms 等[49]方法進行RNA 提取,利用PrimeScript? RT reagent Kit with gDNA Eraser(Perfect Real Time)(RR047)進行反轉錄,以TB Green?Premix Ex Taq? II(Tli RNaseH Plus)(RR820)進行熒光定量PCR,D1的qRT-PCR引物為D1-qF/D1-qR,內參基因引物為UBQ5-qF/UBQ5-qR,序列如表1 所示。反應程序為95 ℃ 30 s;95 ℃ 10 s、60 ℃ 30 s,40 個循環。相對表達量用2-〔d1(CtD1-CtUBQ5)-WT(CtD1-CtUBQ5)〕計算。顯著性分析采用Student’st-test。

表1 供試引物及其序列Table 1 Primers and their sequences used in this study
1.5 表型測量與統計
株高用自地面到主穗頂長度的平均值表示,n=10。穗長用穗頸節到穗頂的長度平均值表示,n=15。粒長、粒寬分別為飽滿種子的長度和寬度平均值n=12。千粒重用1000 粒飽滿種子的質量平均值換算而成,n=20。顯著性分析采用Student’st-test。
2 結果與分析
2.1 水稻中花11 號d1 突變體的創制
為創制中花11 號背景下的d1突變體,我們采用CRISPR/Cas9 基因編輯技術對D1基因進行突變。首先選取位于第二個外顯子上的一段序列為靶點序列(見1.2)作為靶點,此位置與已報道的D1突變位點均不相同。將靶點序列正向引物與下游gRNA 的5'端序列融合,sgRNA 反向引物與上游U6 啟動子的3'端融合,再與pEntry A載體上的引物進行融合PCR,得到U6 啟動子-靶點-gRNA 表達盒。將擴增產物與pRHCas9 載體用PstI 和SpeI 雙酶切,再進行連接得到D1-Cas9 載體(圖1A)。將D1-Cas9 載體導入農桿菌EHA105 中,侵染ZH11 愈傷組織,隨后通過共培養、篩選、分化等步驟得到d1突變體幼苗。
2.2 突變體基因型分析及表達水平檢測
為分析Cas9 是否對D1基因位點進行編輯,我們對d1突變體T1代植株進行DNA 提取,以其為模板,在編輯位點上下游400~500 bp 處設計引物進行PCR 擴增(圖1B),對產物進行Sanger測序,將測序結果與WT 進行比對。發現突變體的D1基因位點發生編輯,3 個株系均為純合移碼突變,其中d1-#1為-17+7 bp,d1-#2為+2 bp,d1-#3為+1 bp(圖2A)。為了解突變株中D1的表達水平,我們對T2代純合突變植株的幼苗進行RNA 提取并采用熒光定量PCR 檢測D1的mRNA水平。結果顯示,3 個突變株系D1的mRNA 水平均顯著下調,下調幅度高達80%~90%(圖2B)。這說明d1突變株在mRNA 和蛋白兩個水平上抑制D1的功能,成功獲得有效的d1突變植株。
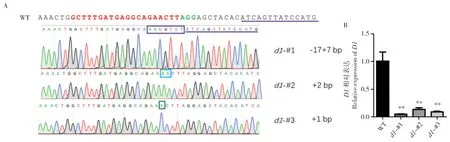
圖2 d1 突變株的基因型鑒定及D1 表達量檢測Fig.2 Genotyping of d1 mutants and detection of expression level of D1
2.3 無潮霉素抗性突變體篩選
轉基因標記是作物向農業生產應用的障礙,易引發公眾對轉基因安全性的憂慮。同時,轉基因產生抗性標記的存在可能為以后基礎研究所需的雜交帶來不便。如兩親本抗性相同,難以用抗性準確篩選雜交成功的后代植株。因此,篩選無潮霉素抗性的d1突變株具有重要意義。農桿菌介導轉基因通常以單拷貝形式隨機插入到基因組中,通過 自交產生T1代時,抗性基因可通過同源重組或自由組合分離,得到無抗性的突變體。利用該原理,我們對T2代植株葉片的DNA進行提取,并且通過PCR 擴增HPT外源基因片段,同時擴增了ACTIN啟動子片段作為內源基因的參照。結果顯示,每個株系隨機選取的若干植株中,均分離出不含外源基因的突變株(圖3),為后續研究與應用奠定基礎。
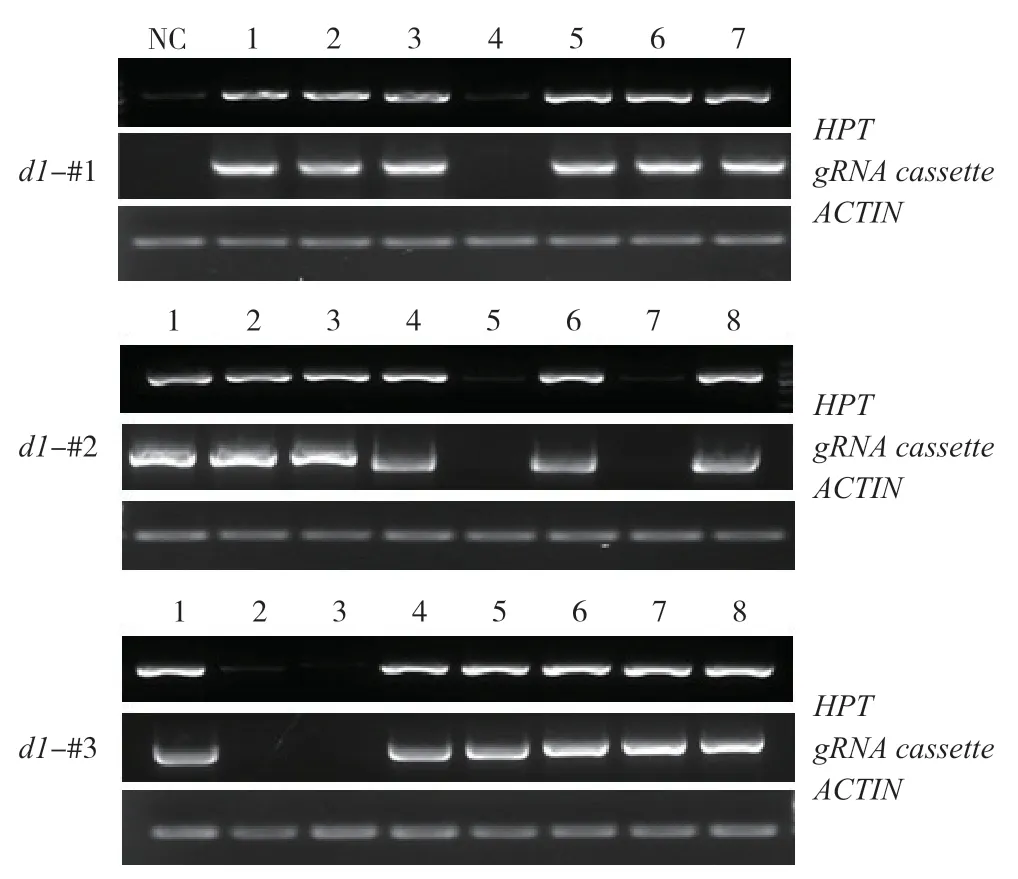
圖3 d1 各突變株系不同植株DNA 中轉基因標記的PCR 檢測Fig.3 Detection of transgenes in genomic DNA of d1 mutants by PCR
2.4 d1 突變體表型分析
以往研究表明,d1突變體的株型、穗型、籽粒形態與大小等與野生型有明顯差異。為全面了解中花11 號背景下d1突變體的表型,我們對植株形態、穗形和種子大小進行觀察(圖4)與統計(圖5)。統計結果顯示,3 個d1突變株系均顯著矮化,株高僅有野生型的約50 %。此外,穗長只占野生型的約60%。d1突變體的種子大小、形態同樣發生明顯變化,其中粒長變得更短,野生型平均粒長為0.74(±0.05)cm,而3 個d1突變株系分別為0.55(±0.06)、0.62(±0.03)、0.55(±0.04)cm。然而,粒寬在d1突變體中無明顯變化,因此,d1突變體種子變得更圓,長寬比為1.29(±0.16)、1.46(±0.12)、1.34(±0.15),中花11 號的種子長寬比則為1.85(±0.10)。同時,d1植株千粒重顯著降低,約為野生型的60 %(圖5F),可見中花11 號背景下d1突變體產生嚴重的生長缺陷。

圖4 d1 突變株的形態特征Fig.4 Morphological characteristics of d1 mutants
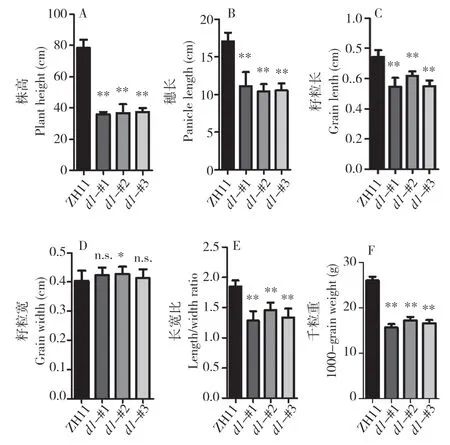
圖5 d1 突變株的表型統計Fig.5 Phenotype statistics of d1 mutants
3 討論
D1是植物生長發育過程中一個重要的調控基因,日本在20 世紀初期已有該基因的水稻突變體。D1基因功能具有多效性,近年陸續出現其他背景下的D1等位突變體,如T65、Shiokari、Kinmaze。這些突變體突變形式各異,包括點突變、堿基插入/缺失導致蛋白翻譯提前終止、結構域失活等,也有剪接位點改變和反義轉錄本導致的RNA 水平降低[44]。此外,科學家還發現了天然的表觀遺傳d1突變體,其DNA 序列并沒有發生變化,然而矮化的d1表觀突變體轉錄起始位點附近DNA 發生甲基化,同時,在D1基因座內,組蛋白H3K9 乙酰化水平降低而H3K9 二甲基化水平升高,表明d1表觀突變體D1的表達被沉默[50]。在雙子葉模式植物擬南芥中,D1同源基因的缺失突變體已有4 個株系被報道,它們屬于Ws和Col背景[51-52]。其他植物種類如大豆、煙草、西紅柿等的D1同源基因功能研究并不深入,多數僅停留在基因的克隆、鑒定階段,缺乏相應的功能缺失突變體[53]。本研究采用CRISPR/Cas9 介導的基因編輯技術對中花11 號的D1基因目標位置進行編輯,鑒定出3 個等位突變株系,豐富了現有的d1突變體類型。
現有研究表明,水稻d1突變體會導致植株矮小、直立,葉長變短且深綠、谷粒變短變圓、節間變短、對GA 和BR 敏感性降低、免疫應答改變等多效表型[44]。我們創制的中花11 號背景下的d1突變體也出現植株矮小、穗小、籽粒變短、籽粒變薄等表型,與前人報道的其他背景下的d1突變體表型相吻合,表明D1在不同水稻品種中的功能高度保守,也證明本研究所得到的材料可有效應用于相關領域研究。
CRISP R/Cas9 基因編輯產生的移碼插入或缺失突變通常引起翻譯提前終止,導致無義RNA降解(Nonsense mRNA Decay,NMD)[54]。本研究所創制的d1突變體材料中,D1基因的mRNA水平下調約80%,我們分析d1突變體的編輯情況發現,d1-#1和d1-#2突變體均使D1 蛋白在第94 位氨基酸位置處提前終止,而d1-#3突變體會使D1 蛋白翻譯在第35 氨基酸處提前終止,這些因素很可能觸發mRNA 質量監控機制,導致D1mRNA 經歷NMD 而下調,這些結果與我們之前在其他基因的CRISPR 敲除植株上觀察到的相似[55]。然而,CRISPR/Cas9 導致NMD 并不是絕對的,與sgRNA 到下游外顯子或終止子的距離遠近有關[56]。可見,CRISPR/Cas9 介導的基因編輯有可能從mRNA 水平和蛋白水平兩方面導致基因功能失活。
4 結論
本研究利用CRISPR/Cas9 技術在中花11 號背景中創制出3 個移碼突變的d1突變株系,qRT-PCR 結果顯示,D1mRNA 水平顯著下調。中花11 號背景的d1敲除植株出現植株矮小、穗小、籽粒變小、變圓等表型,千粒重也顯著減少,株高、穗長、粒長、長寬比和千粒重均下降40%~50%,而粒寬無明顯變化,這些表型與其他背景中現有的d1突變體相似,說明D1基因功能已經失活。我們還從T2代植株篩選出不含轉基因標記的植株。鑒于中花1 1 號基因組已被研究得非常透徹,被廣泛用于基因功能研究,現有的突變體庫也以它為背景創制[22,23],擁有中花11 背景的d1突變株對創制D1與其他基因的多重突變株更加便利。因此,本研究結果可以為后續的水稻基礎研究與育種應用提供安全、有效的遺傳材料。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
汽車工程師(2021年12期)2022-01-17 02:29:54
軍事文摘(2021年22期)2021-11-26 00:43:51
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2020年14期)2021-01-08 09:30:42
文苑(2020年6期)2020-06-22 08:41:52
科技傳播(2019年22期)2020-01-14 03:06:54
文苑(2019年22期)2019-12-07 05:29:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
