納米抗體在真菌毒素檢測中的應用
2023-01-13 08:40:40吳紹文鄒新路胥錦濤晏石娟
廣東農業科學 2022年11期
吳紹文,鄒新路,胥錦濤,晏石娟
(廣東省農業科學院農業生物基因研究中心/廣東省農作物種質資源保存與利用重點實驗室,廣東 廣州 510640)
真菌毒素是由曲霉屬(Aspergillus)、青霉屬(Penicillium)、鐮刀霉菌屬(Fusarium)、麥角菌屬(Claviceps)等真菌產生的一類天然次生代謝產物。這些產毒真菌能夠污染含油量高的種子(如花生、玉米)、谷物和堅果等,而這些農作物及其加工而成的農產品是人類與畜禽的主要食物來源,被食用真菌毒素污染的食物會導致癌癥、免疫缺陷等多種疾病,危害人類生命健康[1-2]。目前,已經有數百種不同的真菌毒素被鑒定出來,其中對人類健康和牲畜造成嚴重危害的真菌毒素主要包括黃曲霉毒素(Aflatoxin,AF)、赭曲霉毒素(Ochratoxin,OT)、伏馬菌素B(Fumonisin B,FB)、玉米赤霉烯酮(Zearalenone,ZEN)和脫氧雪腐鐮刀菌烯醇(Deoxynivalenol,DON)等。產毒真菌的生長可能發生在農作物及農產品收獲前后、加工、運輸和存儲等過程中,而其產生的大多數真菌毒素的化學性質穩定,可以在食物鏈中殘留[3]。在農產品貿易全球化和全球氣候變暖的背景下,農產品存儲和運輸時間長、環境差異大等因素促使真菌毒素污染概率增加,從而導致國際貿易中的巨大經濟損失。
由于真菌毒素是食品安全風險的常見因素,許多國家和組織都制定了農產品中真菌毒素的限量標準。例如,歐盟委員會規定,未加工谷物中黃曲霉毒素B1(AFB1)、赭曲霉毒素A(OTA)、FB、DON 和ZEN 的限量分別為2、5、4 000、1 250、100 μg/kg[4]。因此,真菌毒素高靈敏檢測方法的研發對于防控真菌毒素污染至關重要。目前,常用的真菌毒素檢測方法包括高效液相色譜法、色譜-質譜聯用法和免疫測定法[5-7]。其中,免疫測定法基于抗原和抗體特異性結合的原理,具有低成本、高通量和高靈敏度等特點,且操作簡便,對檢測設備的要求較低,是真菌毒素大規模篩查的常用檢測手段。
抗體是免疫測定法的核心試劑,針對真菌毒素的檢測,人們已經研發了大量抗體,包括多克隆抗體、單克隆抗體和重組抗體等多種類型[4,8]。常規的單克隆抗體和多克隆抗體由兩條重鏈和兩條輕鏈組成,其中重鏈包含3 個恒定區(CH1、CH2、CH3)和1 個可變區(VH),而輕鏈只包含1 個恒定區(CL)和1 個可變區(VL),重鏈和輕鏈的互補性決定區(CDR)有助于形成抗 體的抗原結合表位(圖1)。在駱駝科和軟骨魚的血清中,存在一種缺乏輕鏈和CH1 結構域的重鏈抗體(hcAb),其抗原結合位點由重鏈的可變區(VHH)形成。這些獨特的VHH 結構域能夠通過重組蛋白表達的方式獲得,是目前已知的抗原-抗體相互作用的最小單元,由于其大小在納米尺度,因此被稱為納米抗體(Nanobody,Nb)。與抗原結合片段(Fab)、單鏈可變片段(scFv)等重組抗體片段相比,納米抗體具有相當或更強的抗原結合能力、優異的物理化學穩定性和折疊能力,且更容易進行遺傳改造,這些特性使得納米抗體成為用于免疫測定的極佳抗體試劑[9-10]。目前,已有許多與真菌毒素檢測相關的納米抗體被報道,這些抗體被廣泛應用于真菌毒素檢測技術的開發。本文圍繞常見真菌毒素檢測相關的納米抗體研發現狀進行總結,并分析基于納米抗體開發的真菌毒素檢測技術,以期為進一步開發高靈敏、低成本、易應用的真菌毒素檢測方法提供線索和思路。
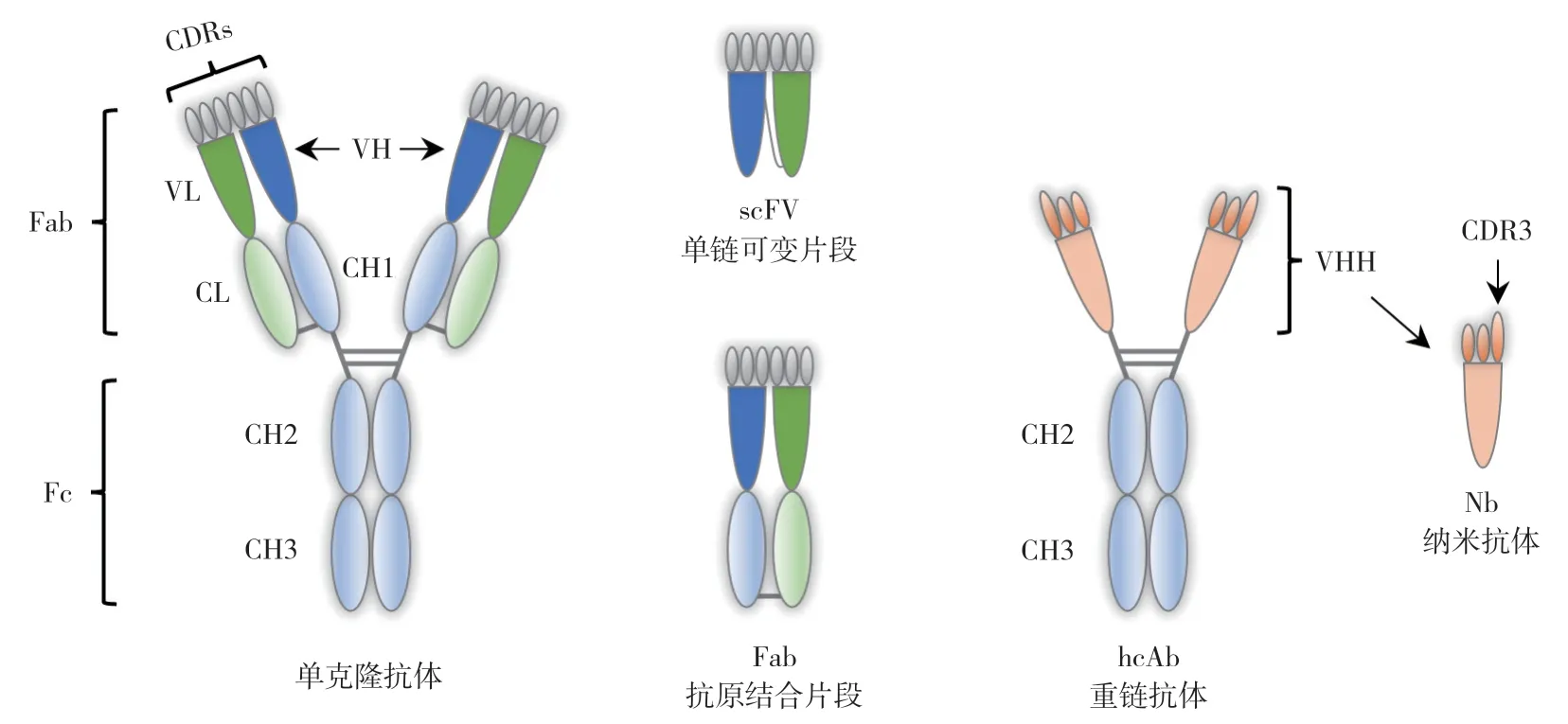
圖1 單克隆抗體、單鏈可變片段、抗原結合片段、重鏈抗體和納米抗體的結構示意圖Fig.1 Schematic structures of monoclonal antibody,single-chain variable fragment (scFV),antigen-binding fragment (Fab),heavy-chain-only antibody (hcAb) and nanobody (Nb)
1 抗不同真菌毒素特異性納米抗體的研發
用于真菌毒素檢測的納米抗體主要包括抗真菌毒素特異性抗體和抗真菌毒素單克隆抗體的獨特型抗體兩種類型(圖2),其中抗真菌毒素特異性納米抗體可直接結合毒素小分子或其偶聯的蛋白復合物,而抗獨特型納米抗體則通常結合真菌毒素的單克隆抗體。在已知的真菌毒素中,黃曲霉毒素和赭曲霉毒素由于毒性強、危害大、分布廣等特點,被歐盟委員會列為致癌物質,并引起人們的廣泛關注,進而在這兩種真菌毒素的抗特異性納米抗體和檢測技術的研發上做了大量工作。
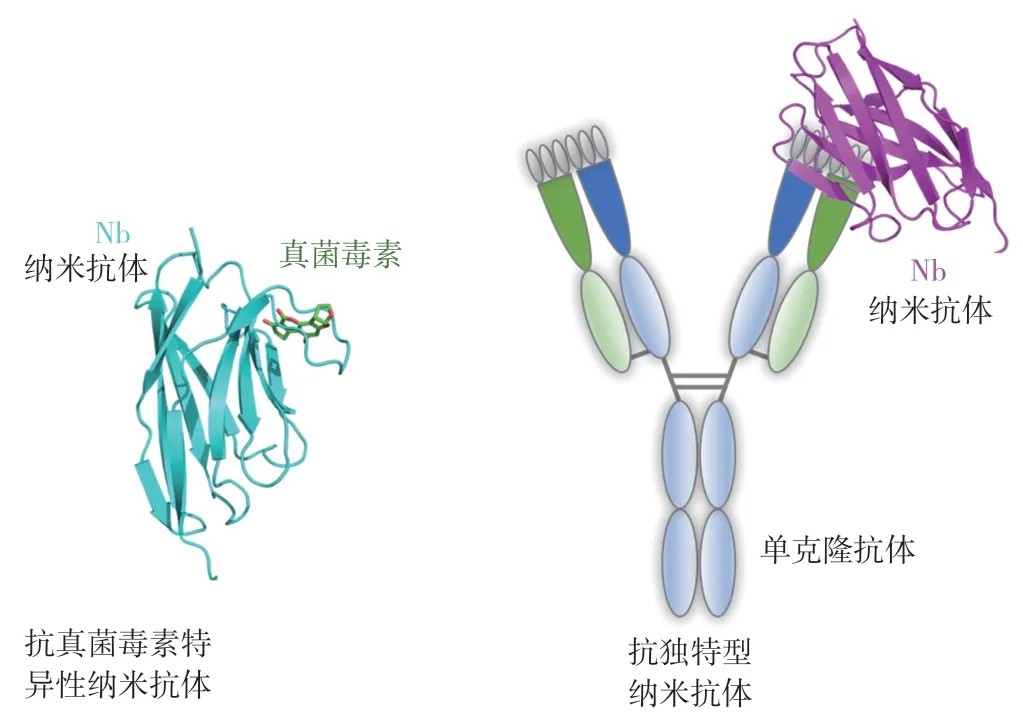
圖2 抗真菌毒素特異性納米抗體和抗獨特型納米抗體的結構示意圖Fig.2 Schematic structures of anti-mycotoxin nanobody and anti-idiotypic nanobody
1.1 抗黃曲霉毒素納米抗體
黃曲霉毒素主要由生長在土壤、腐爛植被、干草和谷物中的一些霉菌(特別是黃曲霉菌、寄生曲霉菌等)產生。這些霉菌容易污染谷物(如玉米、高粱、小麥和大米)、油料作物種子(如大豆、花生、向日葵和棉花種子)、香料作物(如辣椒、黑胡椒、芫荽、姜黃和生姜)、堅果(如開心果、杏仁、胡桃和巴西堅果)等及其加工而成的農產品(如陳皮)[1-2,11]。目前已有20 余種黃曲霉毒素被報道,主要包括AFB1、AFB2、AFG1 和AFG2 等4 種類型[12]。此外,在飼喂受污染飼料后動物乳汁中也發現含有AFM1。黃曲霉毒素具有遺傳毒性,可以造成DNA 損傷,因此食用被黃曲霉毒素污染的食物與癌癥的發病率呈正相關性,大劑量的黃曲霉毒素還可導致急性中毒甚至危及生命。
早在20 世紀80 年代,人們就開始了抗黃曲霉毒素單克隆抗體和多克隆抗體的研發,而抗黃曲霉毒素的納米抗體則在近10 年才取得突破[12]。2014 年,He 等[12]首次使用牛血清白蛋白BSA偶聯的AFB1(AFB1-BSA)免疫羊駝構建的VHH 噬菌體展示文庫,篩選得到兩種抗AFB1 的特異性納米抗體NB26 和NB28。基于這兩種納米抗體的酶聯免疫吸附測定(ELISA)實驗結果顯示,其檢測AFB1 的半數抑制率(IC50)分別為0.754、1.012 ng/mL,表現出很強的AFB1 結合能力。不同黃曲霉毒素的交叉反應實驗結果顯示,NB26 和NB28 對AFB2、AFG1、AFG2 的交叉反應性均低于10%,而對AFM1 的交叉反應性分別為1 0.75%和32.7%,表明這兩種納米抗體結合AFB1 的特異性均較強。進一步試驗發現這兩種納米抗體的熱穩定性和抗有機溶劑的特性明顯優于傳統單克隆抗體。在水稻、花生、玉米和飼料等實際樣品的檢測中,基于NB26 的ELISA 實驗檢測的回收率在80%~115%。上述結果表明,這兩種抗AFB1的納米抗體具有優異的檢測能力。曹冬梅等[13-14]使用相似方法篩選得到抗AFB1 的納米抗體G8,其結合AFB1 的能力比NB26 和NB28 稍弱,在最優的ELISA 實驗條件下測定結合的IC50值為4.61 ng/mL。
除上述提到的ELISA 方法外,抗黃曲霉毒素特異性的納米抗體還被廣泛應用于不同免疫測定方法的開發,主要包括一步法酶聯免疫吸附分析ALP-ELISA、生物發光酶聯免疫分析BLEIA和磁性化學發光酶聯免疫分析MCLEIA 等。使用納米抗體G8,曹冬梅等[13]開發了一種基于G8-堿性磷酸酶ALP 融合蛋白的ALP-ELISA。使用G8-納米熒光素酶NLUC 融合蛋白,Ren 等[15]開發了一種檢測AFB1 的BLEIA 方法,通過對檢測體系進行優化,使用這一方法檢測AFB1 的IC50值為0.41 ng/mL,低于ALP-ELISA 和傳統ELISA 方法,檢測靈敏度更高。使用生物素標記的納米抗體NB26,Yan 等[16]開發了一種基于生物素-鏈霉親和素放大檢測信號的BA-ELISA 方法,該方法檢測AFB1 的IC50值為0.21 ng/mL,檢測靈敏度得到進一步提高。通過篩選與納米抗體NB28 結合的多肽,以多肽模擬人工抗原,Zhao 等[17]開發了一種基于磁珠快速定向競爭的MB-dcELISA 方法,這一方法檢測AFB1 的IC50值為0.75 ng/mL。使用相似策略,Zou 等[18]建立了一種直接非競爭性磁化學發光酶聯免疫分析方法Nc-MCLEIA,將基于NB28 免疫測定的IC50值降低至0.089 ng/mL,明顯提高了AFB1 檢測的靈敏度。更為重要的是,He 等[19]使用核磁共振和分子對接的方法解析了納米抗體NB26 的三維結構及其結合AFB1 的可能機制,并根據NB26 與AFB1 的結合位點信息設計了NB28 的突變體,用于提高其對AFB1 的結合能力,其中基于NB28的A50V、S102D 兩個突變體的免疫分析檢測靈敏度分別比野生型提高2.3 倍和3.3 倍。
近年來,納米抗體與傳感器聯用的方法得到廣泛關注。例如,Pan 等[20]首次篩選了多個抗AFB1 的納米抗體NB30、NB33 和NB70,盡管其未對這些納米抗體結合AFB1 的能力進行詳細評估,但使用NB70 制備的電化學免疫傳感器(EC-immunosensor)測定AFB1 的檢測限低至0.0033 ng/mL。而Liu 等[21]將納米抗體G8 和辣根過氧化物酶HRP 串聯體修飾的雜交鏈反應信號放大系統相結合,構建了一種新型電化學競爭免疫傳感器(ECC-immunosensor),其檢測限低至68 fg/mL,且具有良好的檢測特異性和穩定性。這些研究表明,將納米抗體與其他材料相結合能夠進一步拓寬其在真菌毒素檢測方面的應用。
1.2 抗赭曲霉毒素納米抗體
赭曲霉毒素是由赭曲霉菌、碳曲霉菌等曲霉菌以及一些青霉菌產生的真菌毒素,主要包括OTA、OTB 和OTC 等類型,其中OTA 是最常見的赭曲霉毒素,主要在谷物、葡萄酒、肉類、奶類和面包等農產品的儲藏過程中形成[22]。OTA對人類健康具有多種毒性作用,包括明顯的腎臟毒性、免疫毒性、致畸性和致突變性等,會導致腎癌等癌癥,被國際癌癥研究機構列為2B 類致癌物,其對胎兒發育和免疫系統也有影響[23]。因此,許多國家均對農產品和食品中的OTA 含量進行嚴格限定,如我國食品安全國家標準(GB 2761-2017)中規定谷物、豆類和堅果中的OTA 限量為5 μg/kg,葡萄酒中的限量為2 μg/kg。
抗OTA 納米抗體的研發最早可以追溯到2014 年,Liu 等[24]從免疫羊駝獲得的VHH 文庫中篩選得到VHH15、VHH28、VHH32 和VHH36等4 個抗OTA 納米抗體。其中,VHH28(也稱NB28)在噬菌體ELISA 實驗中測定的IC50值為0.31 ng/mL,交叉反應實驗顯示其對OTB 的交叉反應率為3.5%,表明NB28 對OTA 具有優異的選擇性。基于這一抗體開發的實時免疫PCR(RT-IPCR)在2.5% 的甲醇-PBS 溶液中OTA 檢測限為3.7 pg/mL、線性范圍IC20~80為0.01~1 000 pg/mL,在玉米、小麥和大米等實際樣品中檢測的回收率為80%~126%,表明基于NB28 開發的方法具有優異的OTA 檢測能力。Sun 等[25]開發了一種使用NB28-短肽AviTag 融合蛋白檢測OTA 毒素的BA-ELISA 方法,該方法檢測OTA 的IC50值為0.14 ng/mL、檢測限為0.028 ng/mL,在大麥、燕麥和大米等實際樣品中的OTA 檢測限分別為1.40、0.56、0.84 μg/kg。
為簡化基于抗OTA 納米抗體的檢測方法,Wang 等[26]開發了基于Nb28-堿性磷酸酶融合蛋白的AP-ELISA,該方法檢測OTA 的IC50值為0.46 ng/mL、檢測限為0.12 ng/mL、線性范圍為0.20~1.26 ng/mL。Tang 等[27]開發了一種非競爭性的熒光共振能量轉移免疫測定法(FRET immunoassay),實現了對OTA 和OTB 兩種毒素的同時檢測,這一方法對OTA 和OTB 的檢測限分別為0.06、0.12 ng/mL;該作者還進一步搭建了基于FRET 的免疫傳感器(Nb-FRET immunosensor),這一傳感器的檢測限為5 pg/mL[28]。Li 等[29]研發了一種基于納米抗體和Au/CaCO3的新型電化學發光免疫測定法(ECIimmunosensor),該方法的檢測限為5.7 pg/mL,接近于Nb-FRET immunosensor 的檢測限,實現了對OTA 的超靈敏檢測。
2021 年以來,研究者們利用蛋白質工程開發了一系列更加簡便的OTA檢測方法。例如,Su等[30]基于Nb28-SGFP、SGFP-Nb28兩種不同熒光蛋白-納米抗體融合蛋白,構建了OTA 檢測的熒光納米傳感器(FN-Nanosens),FN-Nanosens 的檢測限為5 pg/mL、線性范圍為5~5 000 pg/mL。而Xie等[31]開發了一種基于納米抗體/NanoBiT 系統的生物發光免疫傳感器(NBL-Immunosens),這一傳感器檢測OTA 的IC50值為0.31 ng/mL、檢測限為0.01 ng/mL、線性范圍為0.04~2.23 ng/mL。以上兩種傳感器均可對OTA 完成快速高靈敏檢測。此外,Wang 等[32]使用定向進化和飽和突變的方法獲得NB28 的雙位點突變體G53Q &S102D,不同溫度的測試實驗顯示G53Q &S102D 突變體具有良好的耐熱性,而使用野生型和這一突變體檢測OTA的IC50值分別為0.41、0.29 ng/mL,表明突變體的檢測靈敏度相比野生型約提高1.4 倍。基于NB28 突變體-堿性磷酸酶融合蛋白和MnO2納米片,Zhang 等[33]建立了酶級聯放大免疫測定法(ECAIA)用于檢測咖啡中的OTA,這一方法檢測的IC50值為7.65 ng/mL、檢測限為3.38 ng/mL,實現了對咖啡中OTA 的高靈敏檢測。綜合來看,基于抗OTA 納米抗體NB28 開發了多種快速、簡便且靈敏度高的檢測方法,借鑒這些思路開發其他真菌毒素的快速高靈敏檢測方法是未來值得開展的研究。
1.3 其他真菌毒素的特異性納米抗體
目前,抗細交鏈孢菌酮酸(Tenuazonic acid,TeA)、異細交鏈孢菌酮酸(Iso-tenuazonic acid,ITeA)、DON 和15-乙酰基脫氧雪腐鐮刀菌烯醇(15-Acetyldeoxynivalenol,15-AcDON)等真菌毒素的特異性納米抗體也有報道。細交鏈孢菌能夠污染谷物和水果等農產品并產生TeA,TeA 是一種獨特的四酸衍生物,具有強效的真核蛋白質合成抑制活性,可阻止核糖體釋放新合成的蛋白質,也是細交鏈孢菌產生的毒性最強的一種毒素。Wang 等[34]使用噬菌體展示文庫篩選得到一種抗TeA 的特異性納米抗體Nb-3F9,并基于這一抗體及其與NLUC 的融合蛋白分別開發了ELISA、化學發光酶聯免疫分析(CLEIA)和BLEIA 等TeA檢測技術,這些方法中ELISA 檢測的IC50值為54.8 ng/mL,CLEIA 和BLEIA 檢測的IC50值分別為8.6 ng/mL 和9.3 ng/mL,靈敏度比ELISA 提高近6 倍。ITeA 是TeA 的同分異構體,化學性質和毒性有所不同。Wang 等[35]篩選得到一種抗ITeA的特異性納米抗體B3G3,基于這一抗體建立的ELISA 方法檢測ITeA 的IC50值為1.3 ng/mL,交叉反應實驗結果表明其檢測TeA 的IC50值為19.8 ng/mL、交叉反應率為6.6%,顯示出較好的ITeA 結合特異性。
DON 及其衍生物3-乙酰脫氧雪腐鐮刀菌烯醇(3-AcDON)和15-AcDON 是谷物中常見的真菌毒素,主要由禾谷鐮刀菌和亞洲鐮刀菌在侵染小麥時產生[36]。動物和人類攝入含有DON的食物會導致惡心、胃腸不適、嘔吐和腹瀉等癥狀,乙酰化的DON 由于在腸道中的吸收速度更快,故毒性比DON 更強[37]。Doyle 等[38]克隆、表達并表征了一種抗15-AcDON 的特異性納米抗體NAT-267,但這一抗體結合15-AcDON的能力較弱,熒光偏振分析實驗(Fluorescence polarization,FP)測定其結合的解離常數為1.24 μmol/L(即419.6 ng/mL)。進一步研究發現,將納米抗體NAT-267 在酵母細胞內表達能夠明顯降低15-AcDON 對細胞的毒性[39]。隨后,Tu 等[40]使用噬菌體展示文庫篩選得到抗DON 的特異性納米抗體B10、C8 和C15,噬菌體ELISA 實驗發現B10 結合DON 的能力更強。上述研發的特異性納米抗體在真菌毒素的檢測方面表現出良好的性能和應用前景(表1),因此,進一步研發其他常見真菌毒素的特異性納米抗體并提高其應用性能,是未來值得開展的重要工作。
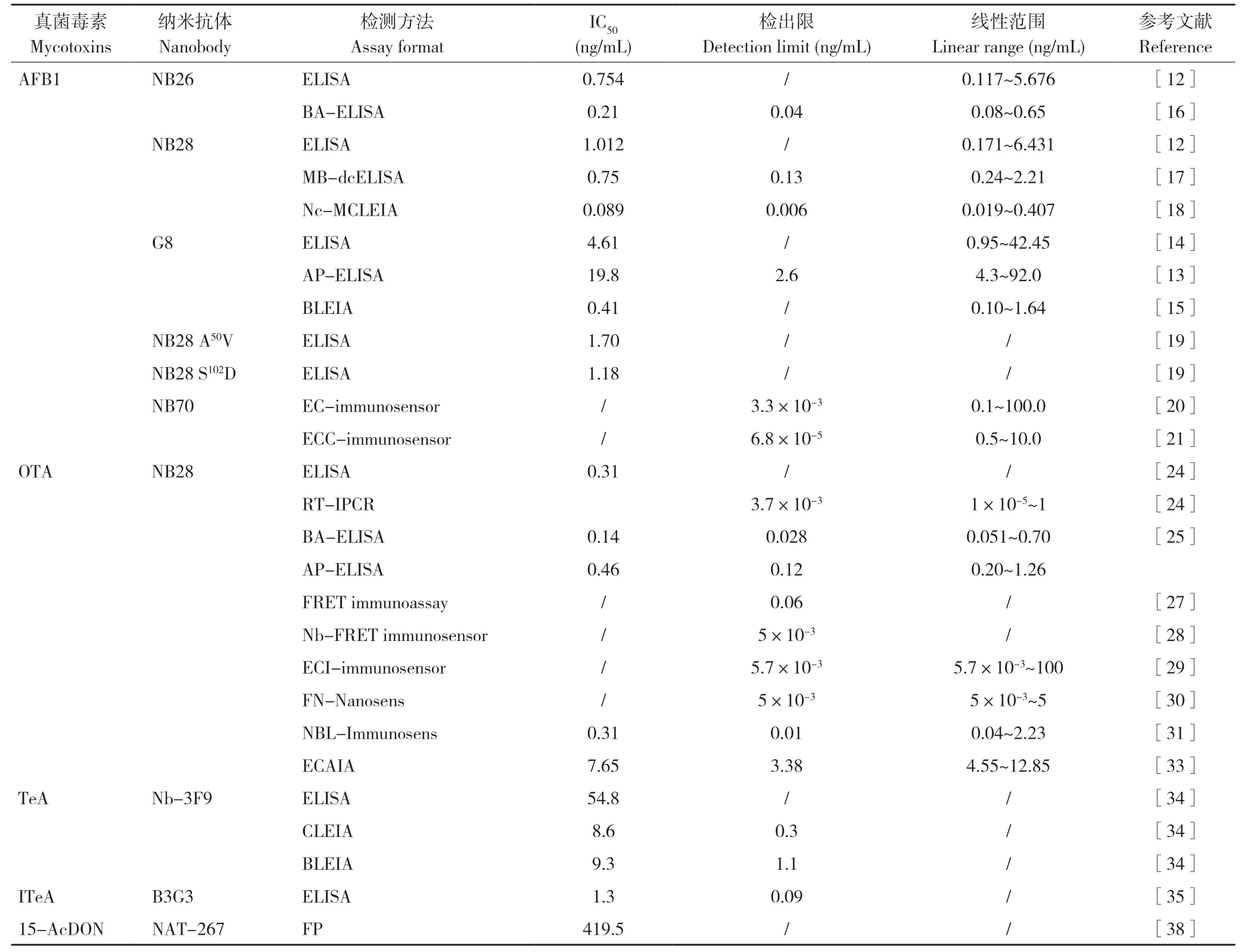
表1 基于抗特異性納米抗體的真菌毒素檢測方法和性能Table 1 Performance of methods for mycotoxins detection using anti-mycotoxin nanobodies
2 抗獨特型納米抗體在真菌毒素檢測中的應用
抗獨特型納米抗體是一類可以與抗原競爭結合其一級抗體可變區的第二抗體,能夠替代小分子化合物和蛋白偶聯物用于免 疫測定[41-42]。因此,相比于抗真菌毒素的特異性納米抗體,抗獨特型納米抗體應用于真菌毒素檢測時有一個明顯優勢,即可代替真菌毒素和蛋白偶聯物作為抗原,從而降低檢測的潛在風險,更加綠色環保[41]。抗獨特型納米抗體已經被廣泛應用于常見真菌毒素的檢測。
2.1 應用于黃曲霉毒素檢測的抗獨特型納米抗體
使用抗黃曲霉毒素單克隆抗體1C11 免疫羊駝的噬菌體文庫,Wang 等[43]篩選得到3 種抗獨特型納米抗體VHH2-5、VHH2-12 和VHH2-29,并將其作為包被抗原應用于黃曲霉毒素的免疫分析。ELISA 實驗結果表明,單克隆抗體1C11和納米抗體VHH2-5 的組合能夠識別AFB1、AFB2、AFG1、AFG2 和AFM1 等5 種黃曲霉毒素,其中識別AFB1 的靈敏度最高,檢測的IC50值為0.16 ng/mL。基于含有VHH2-5 的M13 噬菌體及其編碼DNA,Lei 等[44]建立了一種用于檢測農產品中黃曲霉毒素的RT-IPCR 方法,這一方法的檢測限為0.02 ng/mL,檢測靈敏度優于傳統的ELISA 方法。隨后,Wang 等[45]通過建立VHH 2-5 與AFB1 之間的定量轉換方程,研發了一種定量測定農產品中AFB1 含量的方法(toxinfree ELISA),這一方法的靈敏度進一步提升,其檢測限為0.015 ng/mL。此外,Lei 等[46]基于VHH2-5-噬菌體體系研發了一種免疫環介導等溫擴增(iLAMP)檢測方法,能夠完成對實際樣本中黃曲霉毒素的現場檢測。
Tang 等[47]使用抗AFM1 的單克隆抗體2C9篩選得到兩種獨特型納米抗體VHH4-1-1 和VHH2-3-1,ELIS A 方法檢測得到這些納米抗體結合AFM1 和2C9 的IC50值分別為8.54 ng/mL 和3.33 ng/mL;進一步使用納米抗體VHH4-1-1 功能化的絲網印刷碳電極,研發得到一種無毒電化學免疫傳感器(Toxin-free EC-immunosensor),用于檢測牛奶中的AFM1,結果表明這一傳感器檢測AFM1 的線性范圍為0.25~5.00 ng/mL、檢測限為0.09 ng/mL,性能優異。
2.2 應用于赭曲霉毒素檢測的抗獨特型納米抗體
使用抗OTA 的單克隆抗體和天然納米抗體文庫,Ji 等[48]篩選得到抗獨特型納米抗體P28,基于P28 的噬菌體ELISA 實驗顯示其檢測OTA 的IC50值為0.3 ng/mL。使用抗OTA 的單克隆抗體1H2,Zhang 等[49]篩選得到其抗獨特型納米抗體VHH2-24,ELISA 實驗顯示VHH2-24 能夠特異性地識別1H2,而不能結合抗AFB1、DON 和ZEN的單克隆抗體,表明其具有良好的抗體結合特異性,基于VHH2-24 的ELISA 實驗顯示其檢測OTA 的線性范圍為0.003~0.1673 ng/mL、檢測限為0.001 ng/mL。使用相同的單克隆抗體1H2,Zhang等[50]篩選得到兩種新的抗獨特型納米抗體NCA1和NCA2,序列分析發現,NCA1 和NCA2 兩種納米抗體在CDR 區域有6 個氨基酸不同,它們檢測OTA 的IC50值分別為0.052、0.015 ng/mL,表明NCA1 的檢測靈敏度比NCA2 低。以上結果表明抗獨特型納米抗體CDR 區域的氨基酸變化能夠改變其結合親和力從而提高免疫分析的靈敏度。隨后,Zhang 等[50]使用納米抗體NCA2 建立了一種環境友好的ELISA 方法,用于快速檢測谷物中的OTA 含量,優化后的IC50值為0.017 ng/mL、檢測限為0.003 ng/mL,這一方法在玉米、大米和小麥等實際樣品上的平均檢測回收率在80.0%~114.8%,具有良好的應用性。
2.3 應用于伏馬菌素檢測的抗獨特型納米抗體
伏馬菌素是一類由鐮刀菌屬和赤霉菌屬侵染收獲前或存儲期間的農產品而產生的真菌毒素,目前已鑒定出10 多種結構,其中伏馬菌素B1(FB1)的含量最高、毒性最強,能夠引起癌癥等多種疾病,并被國際組織列為2B 類致癌物[41]。Shu 等[41,52]首先從抗FB1 單克隆抗體的噬菌體展示文庫中篩選得到其抗獨特型納米抗體B26,ELISA 實驗測定得到其檢測FB1 的IC50值為1.07 ng/mL,線性范圍為0.27~5.92 ng/mL,其靈敏度約為基于FB1-BSA 偶聯物的ELISA 方法的20 倍(IC50值為21.14 ng/mL)。進一步交叉反應實驗結果顯示,基于納米抗體B26 的ELISA 實驗檢測伏馬菌素B2(FB2)的IC50值為27.67 ng/mL、交叉反應率為4.93%,表明B26 具有很好的FB1結合特異性。使用玉米、大米和飼料等實際樣本檢測的結果顯示,基于B26 的ELISA 方法的檢測回收率為71.90%~112.15%,表明其適用于實際樣品的檢測。基于B26-堿性磷酸酶融合蛋白,Shu等[52]進一步開發了用于FB1 檢測的AP-ELISA,這一方法檢測的IC50值為0.89 ng/mL、線性范圍為0.29~2.68 ng/mL,靈敏度略高于傳統ELISA。
2.4 應用于脫氧雪腐鐮刀菌烯醇檢測的抗獨特型納米抗體
為建立基于納米抗體的環境友好型DON 免疫分析方法,Qiu 等[53]使用抗DON 單克隆抗體和天然噬菌體展示文庫篩選得到兩種抗獨特型納米抗體N-28 和N-31,ELISA 實驗結果表明這兩種抗體檢測DON 的IC50值分別為8.77、19.97 ng/mL,檢測靈敏度分別約為傳統基于DON-BSA 的ELISA 方法(IC50值為163.69 ng/mL)的18、8 倍;隨后,作者又使用飽和誘變技術進行N-28 的突變體篩選,并證實T102Y、V103L 和Y105F 等位點的突變能夠進一步提高N-28 檢測DON 的靈敏度[54]。
2.5 應用于玉米赤霉烯酮檢測的抗獨特型納米抗體
玉米赤霉烯酮(ZEN)是一種由禾谷鐮刀菌、木賊鐮刀菌和黃色鐮刀菌等鐮刀菌屬合成的非甾體雌激素類真菌毒素,具有致畸性、遺傳毒性、致癌性、肝毒性和免疫抑制等多種危害作用,也是谷物中常見的風險因素之一。為建立基于無毒替代試劑的ZEN 免疫分析方法,Wang 等[55]使用抗ZEN 單克隆抗體和VHH 噬菌體展示文庫篩選得到納米抗體Z1,這一納米抗體具有很好的抗ZEN 單克隆抗體結合能力;該作者利用納米抗體Z1 建立了間接競爭噬菌體ELISA 方法,結果顯示該方法檢測ZEN 的IC50值為0.25 ng/mL、線性范圍為0.11~0.55 ng/mL,相比于傳統的基于ZENBSA 的方法靈敏度提高26 倍。而使用這一納米抗體建立的噬菌體展示介導的IPCR 方法檢測限更低(6.5 pg/mL)、線性范圍為0.01~100 ng/mL,比噬菌體ELISA 方法的靈敏度更高、檢測范圍更寬[55]。此外,Tang 等[56]使用兩種抗獨特型納米抗體建立了一種能夠同時檢測AFB1 和ZEN 的時間分辨熒光免疫層析法,這一方法的最大特點是實現了兩種真菌毒素的同時檢測,其IC50值分別為0.46、0.86 ng/mL。隨后,Ren 等[57]又開發了一種雙功能噬菌體展示IPCR 技術用于同時檢測AFB1 和ZEN,這一方法檢測AFB1 和ZEN 的IC50值分別為0.43、2.12 ng/mL,檢測限為0.03、0.15 ng/mL;試驗表明這一方法可以同時檢測小麥、大米、玉米和飼料樣品中的AFB1 和ZEN,拓寬了抗獨特型納米抗體在真菌毒素檢測中的應用范圍。
2.6 應用于其他真菌毒素檢測的抗獨特型納米抗體
除上述常見的真菌毒素外,還有一些關于TeA 和橘霉素(Citrinin,CTN)的抗獨特型納米抗體的研究。例如,Wang 等[58]使用抗TeA 的單克隆抗體3F10 和噬菌體展示文庫篩選得到用于TeA 檢測的獨特型納米抗體1B、2C 和2D,ELISA實驗測定其檢測TeA 的IC50值分別為45.8、129.3、24.5 ng/mL,表明納米抗體2D 的檢測靈敏度最高。Wang 等[58]進一步使用2D-NLUC 和2D-鐵蛋白-NLUC 建立了TeA 檢測的BLEIA 方法,其中鐵蛋白能夠形成24 個分子的聚合體,因此被認為能夠進一步增強檢測靈敏度;結果顯示,基于2D-NLUC 的BLEIA 實驗檢測的IC50值為26.6 ng/mL,而基于2D-鐵蛋白-NLUC的BLEIA 實驗檢測的IC50值為10.3 ng/mL,靈敏度相比于2D-NLUC 提高約2.5 倍。
CTN 是一種由曲霉屬、青霉屬和紅曲屬等多種菌種產生的真菌毒素,主要存在于谷物、豆類、水果、蔬菜、草藥和香料以及其他植物中,且通常與其他真菌毒素尤其赭曲霉毒素共存[59]。已有研究表明CTN 具有腎毒性,且能夠通過氧化應激參與誘導細胞凋亡[60]。為建立基于抗獨特型納米抗體的CTN 檢測技術,Xu 等[61]使用抗CTN的單克隆抗體和天然噬菌體VHH 展示文庫篩選得到納米抗體X27,基于這一抗體建立的ELISA方法檢測CTN 的IC50值為44.6 ng/mL、線性范圍為5.0~300.0 ng/mL,其靈敏度是傳統ELISA 方法的2 倍。Xu 等[62]還研發了另一種用于CTN 檢測的抗獨特型納米抗體,其ELISA 檢測的IC50值為10.9 ng/mL,靈敏度進一步提高。Huang 等[63]使用納米抗體X27 建立了CTN 檢測的IPCR 方法,其IC50值為9.86 ng/mL、線性范圍為0.1~1 000 ng/mL,該方法檢測靈敏度提高、且檢測范圍更寬。目前已開發出多種常見真菌毒素的抗獨特型納米抗體,這些抗體在多種方法中表現出很好的檢測性能(表2),但是基于抗獨特型納米抗體的新的檢測方法開發相對較少,是未來值得開展的研究工作。
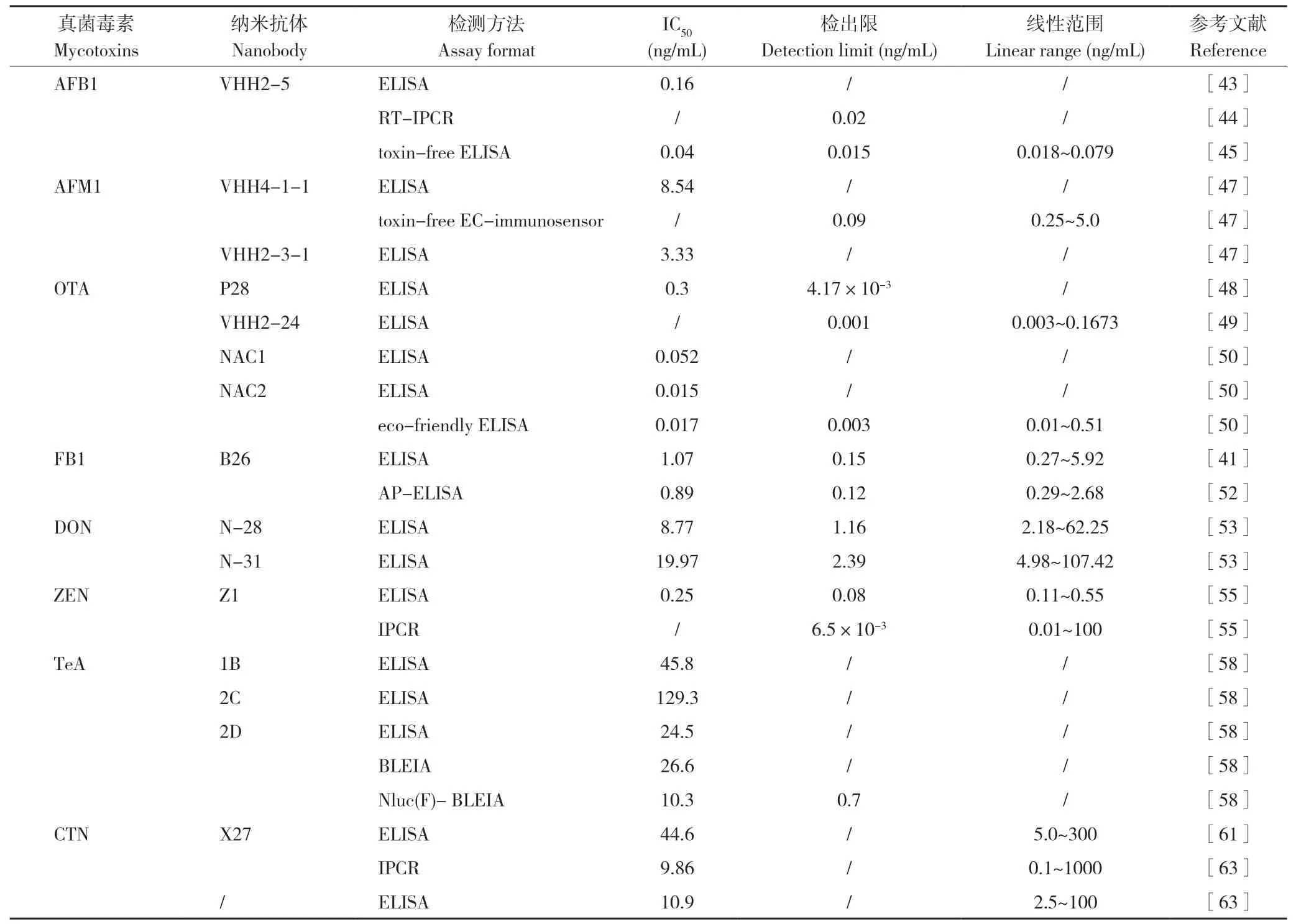
表2 基于抗獨特型納米抗體的真菌毒素檢測方法和性能Table 2 Performance of methods for mycotoxins detection using anti-idiotypic nanobodies
3 總結與展望
隨著我國社會經濟的不斷發展,人們的食品消費觀念正在快速轉變,由“吃得飽”向“吃得好”、“吃得營養”、“吃得健康”轉變,而要滿足人們“吃得健康”的需求則必須加強食品安全監管。根據聯合國糧農組織統計,全球每年約有25%的農產品受到真菌毒素污染。此外,很多中藥如柏子仁、檳榔、地龍等也容易受真菌毒素的污染。中藥在我國人民群眾生命健康和社會經濟發展中發揮著特殊作用,近年來國家也加強了對中藥中真菌毒素限量的監管,如《中國藥典(2015 版)》明確了其收載的19 味中藥中黃曲霉毒素含量的限值,并在《中國藥典(2020 版)》中將限制的中藥品種提高到24 味。我國對真菌毒素污染的監管必將進一步加強,因此迫切需要建立快速、靈敏、成本低且適用于多種毒素同時大規模檢測的方法。
近年來,在噬菌體展示技術和蛋白質工程等方法的推動下,多種常見真菌毒素相關的納米抗體和檢測方法被成功研發。就特定的真菌毒素而言,黃曲霉毒素和赭曲霉毒素有多種抗特異性和抗獨特型納米抗體及相關的檢測方法被開發出來。伏馬菌素、玉米赤霉烯酮和脫氧雪腐鐮刀菌烯醇的抗獨特型納米抗體研究較多,但其抗特異性納米抗體的研究很少。抗獨特型納米抗體相比于抗特異性納米抗體的優勢在于其能夠替代真菌毒素-蛋白偶聯物作為抗原的特性,使得檢測體系更環保,但是需要使用單克隆抗體進行檢測,考慮到單克隆抗體通常無法體外表達,在檢測成本上未必具有優勢。目前,還尚未見到基于這兩種納米抗體的檢測技術在穩定性、應用性和檢測成本等方面的詳細比較,值得未來進一步探索。
從檢測方法來看,相比于傳統的ELISA 方法,ALP-ELISA、BLEIA、熒光和免疫傳感器以及實時免疫PCR 等方法的靈敏度均有提升,檢測參數也均能滿足實際檢測的需求。其中,納米抗體-ALP、納米抗體-NLUC 融合蛋白均能通過體外表達體系獲得,且檢測所需的設備簡單,適合應用于大規模篩查。其中,NLUC 能夠在催化底物存在而不需要其他輔助因子的條件下發出高強度熒光,反應條件簡單,但催化底物的成本偏高。因此,通過蛋白質工程、小分子設計等方法改造NLUC 及其催化底物,從而研發出一套成本更低、靈敏度更高的生物發光體系,有望進一步提升其在一步法酶聯免疫分析技術中的應用性。
從檢測的真菌毒素種類來看,目前免疫分析測定大多數針對單個真菌毒素,少數方法能夠實現兩種真菌毒素的同時檢測。然而,現實農業生產過程中農產品的污染往往是多種真菌毒素同時存在,建立一種能夠實現多種真菌毒素同時檢測的免疫分析技術無疑具有重要意義。另外,建立多種真菌毒素同時檢測的免疫分析技術依賴于高性能的納米抗體,而許多常見的真菌毒素仍然沒有抗特異性納米抗體的研究報道,已有報道的納米抗體也還存在一些應用的缺陷。并且真菌毒素均為小分子化合物,從噬菌體展示文庫中篩選理想的納米抗體具有較高的難度。因此,利用抗體研發的新技術,進一步開發具有高親和力和穩定性的納米抗體極具有挑戰性,也是拓展納米抗體在真菌毒素檢測中應用能力的重要方向。
近年來,抗體和小分子結合機制分析技術、抗體結合多肽的篩選和設計技術、蛋白質理性設計、定向進化、飽和突變和虛擬篩選等前沿技術迅猛發展,為真菌毒素相關納米抗體的研發和改造提供了全新思路。已有研究表明,通過突變體篩選和設計的方式能夠提高納米抗體的真菌毒素結合能力和穩定性,因此通過實驗和計算方法表征真菌毒素與抗體結合的分子機制、解析參與結合的關鍵氨基酸和相互作用力,從而得到抗體與真菌毒素的結合界面信息,對抗體的設計和改造具有重要作用。基于抗體和真菌毒素結合界面的信息,一方面,可以指導抗體結合多肽的實驗和計算篩選,根據結合界面的氨基酸性質,設計和優化多肽文庫,以得到高親和力和穩定性的多肽,并進一步開發基于抗體-模擬肽的真菌毒素檢測方法;另一方面,可以指導納米抗體的定向進化和飽和突變文庫的設計,針對結合界面的氨基酸進行突變,能夠更高效地篩選高親和力的納米抗體突變體,針對結合界面以外的不穩定氨基酸進行突變,能夠進一步篩選高穩定性的突變體甚至研發全新的抗真菌毒素納米抗體,這些研究是未來非常值得嘗試的方向。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
兒童故事畫報(2019年5期)2019-05-26 14:26:14
海峽科技與產業(2016年3期)2016-05-17 04:32:12
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
小雪花·成長指南(2015年7期)2015-08-11 15:03:12
