發酵乳桿菌SS-31培養基及發酵條件的優化
2023-01-10 06:13:48秦雅莉于福田趙笑潁沈圓圓董詩瑜劉小玲
食品與生物技術學報 2022年12期
秦雅莉,于福田,趙笑潁,沈圓圓,董詩瑜,劉小玲*,2
(1.廣西大學輕工與食品工程學院,廣西 南寧 530004;2.廣西水牛乳工程技術研究中心,廣西 南寧 530004)
發酵乳桿菌(Lactobacillus fermentum)是常見的乳酸菌之一,廣泛存在于各類發酵制品中,是異型發酵乳酸菌[1]。美國食品和藥物管理局和歐洲食品安全局均將發酵乳桿菌評估為食品用微生物菌種,并且我國也將其列為《可用于食品的菌種名單》。近年來,隨著研究深入,發酵乳桿菌的功能和益生特性不斷被人們所證實。眾多研究結果表明,發酵乳桿菌在胃腸道中具有高存活率[2]、對病原體的抗菌活性[3]、調節腸道菌群[4]以及抗氧化能力[5],此外還具有免疫調節能力[6]。
乳酸菌廣泛存在于自然界,例如食品、植物、動物或人體腸道等多種生活環境中,是潛在的益生菌。作者所在課題組致力于挖掘柳州酸筍中乳酸菌資源,從酸筍中篩選得到一株具有較強降低RAW264.7 細胞分泌NO 能力的發酵乳桿菌SS-31[7]。該菌株具有較強耐酸能力、耐1 g/L 牛膽鹽、耐人工胃腸液以及高黏附性等特性;此外還可以顯著下調LPS 誘導的RAW264.7 細胞中炎癥因子IL-1β、IL-6的含量。綜上,該菌株具有優良生理特性和抗炎活性。而乳酸菌要在體內達到一定數量時,才能產生一定的益生效果。世界衛生組織規定,益生菌食品的益生菌活菌總數須超過1×107CFU/g 才可以有益生作用。但目前乳酸菌的較低發酵水平限制了其擴大應用。實現菌株低成本、高效率培養的核心是優化發酵培養基。乳酸菌的生長繁殖需要碳源、氮源、無機鹽、營養因子等營養物質,并且生長過程中需要不斷與外界進行物質、能量交換,受到外界環境(溫度、pH 等)影響。與普通培養相比,低成本、高效率的培養既可以更快速提高菌體密度,縮短生產周期,又能降低生產成本[8-9]。呂秀明等通過單因素結合響應面實驗和流加補料培養優化長雙歧桿菌L80培養基,得到活菌數為1.21×1010CFU/mL[10]。Magdalena 等通過響應面法優化鼠李糖乳桿菌培養基,優化后菌體密度達到5 g/L,是相同條件下MRS培養基中菌體密度(1.9 g/L)的2.63 倍[11]。
以發酵乳桿菌SS-31 為研究對象,以活菌數為指標,通過單因素、響應面實驗確定最佳培養基成分和含量。然后對其發酵培養條件(溫度、接種體積分數、pH)進行優化,最后將優化得到的最佳條件應用于15 L 發酵罐,篩選中和劑以進行擴大培養,實現發酵乳桿菌低成本、高效率的發酵培養方式,并為開發以乳酸菌作為主要成分的功能性食品,以及使飼料中益生菌達到相應數量的工業化生產奠定基礎。
1 材料與方法
1.1 實驗材料
發酵乳桿菌SS-31(L.fermentumSS-31):分離自酸筍發酵液,保藏于中國普通微生物菌種保藏管理中心,保藏編號為CGMCC NO.24925。
MRS 基礎培養基:每1 L 含葡萄糖20 g、胰蛋白胨20 g、磷酸氫二鉀0.9 g、硫酸錳0.50 g、硫酸鎂1.00 g、吐溫80 1.00 g,調節pH 至6.5,115 ℃高壓滅菌20 min。
1.2 主要試劑
葡萄糖、麥芽糖、蔗糖、乳糖、蛋白胨、牛肉浸粉、牛肉胨、胰蛋白胨(均為生物試劑):上海麥克林生化科技有限公司產品;氯化鉀、磷酸氫二鉀、磷酸二氫鉀、檸檬酸銨、醋酸鈉、氯化鈉、硫酸鎂、碳酸銨、氨水、碳酸鈉(均為分析純):北京索萊寶科技有限公司產品。
1.3 主要儀器
15 L 發酵罐:上海寶興生物設備工程有限公司產品;GI80TW 立式自動壓力蒸汽滅菌器:致微儀器有限公司產品;DZKW-D-1 電熱恒溫水浴鍋:北京市光明醫療儀器有限公司產品;SW-CJ-2F 潔凈工作臺:蘇州安泰空氣技術有限公司產品;SQP 電子天平:賽多利斯科學儀器有限公司產品;Sherlock 微生物計數儀:美國MIDI 公司產品;DW25-560 恒溫恒濕培養箱:上海知楚儀器有限公司產品;SUNRISE 酶標儀:帝肯(上海)貿易有限公司產品。
1.4 實驗方法
1.4.1 發酵乳桿菌增殖培養基成分的優化以平板涂布法測定的活菌數為評價指標,在MRS 基礎培養基上,替換培養基中碳源,添加量為20 g/L,探究不同種類碳源(葡萄糖、麥芽糖、乳糖、蔗糖)對發酵乳桿菌SS-31 生長的影響,篩選出最佳碳源。以蛋白胨、牛肉浸粉、牛肉胨、胰蛋白胨為氮源,替換上述已優化培養基中的氮源,以20 g/L 為添加量,篩選得到最優氮源。同樣以磷酸氫二鉀、磷酸二氫鉀、檸檬酸銨、醋酸鈉為無機鹽,添加量為9 g/L,篩選出最優無機鹽,以確定最佳增殖培養基成分。
1.4.2 發酵乳桿菌增殖培養基成分質量濃度的優化以平板涂布法測定的活菌數為評價指標,在上述最佳增殖培養基的基礎上,選擇碳源的添加量為5、10、15、20、25、30 g/L,探究其最適添加量。選取上述最適碳源添加量,設置氮源添加量為5、10、15、20、25、30 g/L,確定其最適添加量。選取最佳碳源和氮源添加量,設置無機鹽添加量為3、5、7、9、11、13 g/L,確定其最適添加量。
1.4.3 響應面法優化發酵乳桿菌增殖培養基選擇上述最佳碳源、氮源、無機鹽進行響應面實驗。碳源質量濃度為10、15、20 g/L;氮源質量濃度為15、20、25 g/L,無機鹽質量濃度為7、9、11 g/L,采用三因素三水平(見表1),應用Design-Expert V 8.0.6進行Box-Behnken 實驗設計,優化出增殖培養基最佳配方。
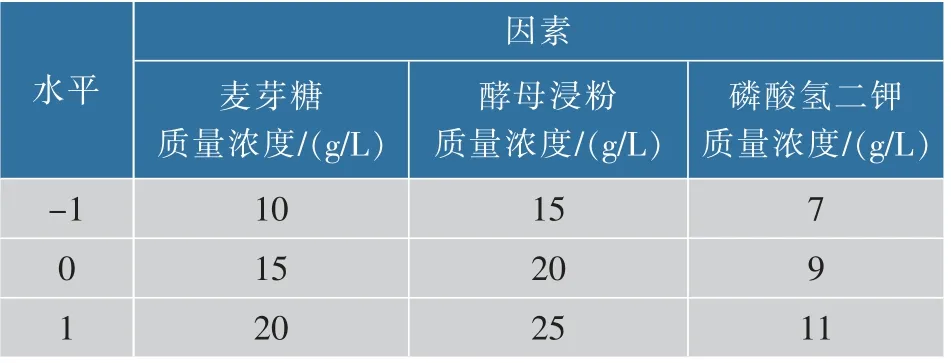
表1 Box-Behnken 實驗設計因素及水平Table 1 Box-Behnken design test factors and levels
1.4.4 發酵乳桿菌SS-31 高密度培養條件的優化
1)接種體積分數的選擇 將已活化3 代后的發酵乳桿菌SS-31 37 ℃恒溫培養24 h 后,按接種體積分數1%、2%、3%、4%、5%,分別接種于增殖培養基中,調節初始pH 為6.8,置于37 ℃恒溫箱內培養24 h,測定其活菌數,確定菌體最佳接種體積分數。
2)初始pH 的選擇 調節增殖培養基初始pH為5.8、6.3、6.8、7.3、7.8,按最佳接種體積分數接種于增殖培養基中,置于37 ℃恒溫培養24 h,測定其活菌數,確定增殖培養基最佳初始pH。
3)溫度的選擇 調節增殖培養基初始pH 至最佳,按最佳接種體積分數接種于增殖培養基中,分別置于31、34、37、40、43 ℃恒溫箱內培養24 h,測定其活菌數,確定最佳培養溫度。
4)中和劑的選擇 為了更快測定菌體濃度,選用OD600為指標反映菌體濃度。選擇常用的Na2CO3溶液和氨水為中和劑,質量濃度均為20 g/L,通過流加中和劑維持培養基pH 為6.8±0.02,以不流加中和劑作空白對照,每隔3 h 測定OD600,并利用平板記數法測定24 h 活菌數。利用15 L 全自動發酵罐,按最終確定的培養條件進行發酵并比較兩種中和劑對發酵乳桿菌SS-31 高密度培養的影響。
1.5 數據處理與統計分析
數據采用SPSS 22.0 軟件進行統計分析,實驗結果以平均值±標準差表示,實驗重復3 次。顯著性(P<0.05)分析使用Duncan 檢驗。使用Origin 9.0 軟件繪圖。
2 結果與分析
2.1 培養基成分的確定
為了篩選出最佳碳源、氮源、無機鹽,以MRS為基礎培養基,采用平板涂布法測定活菌數,以確定最佳碳源、氮源、無機鹽,結果見圖1。在葡萄糖、蔗糖、乳糖、麥芽糖4 種碳源中,麥芽糖是L.fermentumSS-31 生長繁殖的最佳碳源,此時活菌數達到最大值1.64×109CFU/mL,而碳源為葡萄糖的MRS 基礎培養基中活菌數僅為4.67×108CFU/mL。孫媛媛分析了7 種碳源對異型發酵乳桿菌FXJCJ6-1 生長的影響,發現其對單糖(葡萄糖)利用率最高[12]。張興昌等發現以雙糖 (蔗糖) 作為碳源時,Lactobacillus fermentumF6 菌體密度顯著高于其他碳源[13]。但在該實驗中,L.fermentumSS-31 對麥芽糖的利用程度顯著高于其他碳源,說明不同菌株對碳源的利用有差異。
由圖1(b)可知,在胰蛋白胨、牛肉胨、蛋白胨和酵母浸粉4 種氮源中,酵母浸粉是菌體生長繁殖的最佳氮源,此時活菌數達到最大值3.21×109CFU/mL。洪梅等對發酵乳桿菌BLF01 研究發現當酵母粉作為氮源時,獲得的活菌數更高[14]。Gao 等發現酵母粉作為發酵乳桿菌的氮源時,對其菌體生長具有顯著促進作用[15],這也與該實驗研究結果相同。由于酵母浸粉中富含多種氨基酸,還含有較高的尼克酸、葉酸、鈷胺酸,且它們在中、酸性環境中對熱穩定[16],所以酵母浸粉對L.fermentumSS-31 生長有顯著促進作用。
由圖1(c)可知,在磷酸氫二鉀、檸檬酸銨、醋酸鈉、磷酸二氫鉀4 種無機鹽中,磷酸氫二鉀是最適無機鹽,此時活菌數達到最大值3.05×109CFU/mL。無機鹽在一定范圍內可中和菌體在生長過程中產生的酸性物質,并且參與細胞的合成代謝,調節滲透壓穩定[17]。呂佳璐等研究4 種無機鹽對干酪乳桿菌YQ336 生長的影響時,發現磷酸氫二鉀最適宜該菌株的增殖培養[18],這與該實驗結果相同。
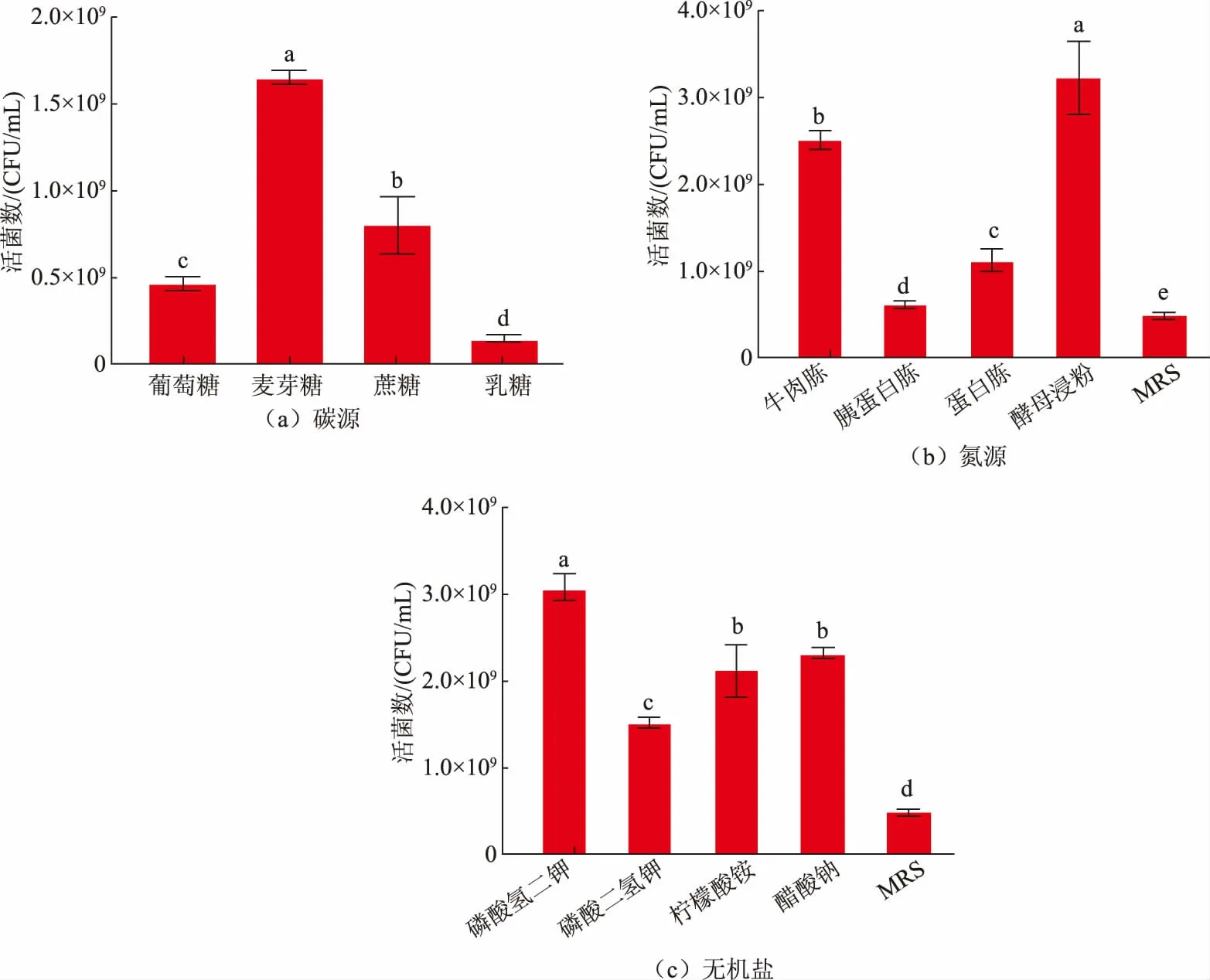
圖1 不同碳源、氮源、無機鹽對發酵乳桿菌SS-31 生長的影響Fig.1 Effects of different carbon sources,nitrogen sources and inorganic salts on the growth of L.fermentum SS-31
2.2 碳源、氮源、無機鹽質量濃度的確定
探究了不同質量濃度麥芽糖、酵母浸粉、磷酸氫二鉀對發酵乳桿菌SS-31 活菌數的影響,結果見圖2。碳源質量濃度對乳酸菌生長起到關鍵作用,隨著麥芽糖由5 g/L 提高到25 g/L,L.fermentumSS-31活菌數呈現先上升后下降的趨勢,并在15 g/L 時,活菌數達到最大值4.92×109CFU/mL。糖質量濃度過高時,菌株前期大量增殖生長,發酵產生大量酸,導致培養基環境偏酸性化,抑制菌株的生長代謝。因此選擇15 g/L 作為麥芽糖的最適添加量。
氮源為菌體生長代謝提供必要元素,富含氨基酸、無機鹽和維生素。從圖2(b)可知,隨著酵母浸粉由5 g/L 提高到25 g/L,L.fermentumSS-31 活菌數呈現先上升后下降的趨勢,并在添加量為20 g/L時,活菌數達到最大值5.87×109CFU/mL。前人研究認為[19]氮源質量濃度過低時,培養基內營養物質不足,導致菌體生長緩慢;氮源質量濃度過高時,菌體生長過快,導致菌體老化、自溶[20]。因此選擇20 g/L作為酵母浸粉的最適添加量。
無機鹽也是菌體生長代謝的重要因素,能夠構成細胞物質以及調節滲透壓。從圖2(c)可知,隨著磷酸氫二鉀由1 g/L 提高到11 g/L,活菌數呈現先上升后下降的趨勢,并在9 g/L 時,L.fermentumSS-31活菌數達到最大值5.47×109CFU/mL。由于乳酸菌在生長代謝過程中分解糖類產生大量乳酸,隨著時間延長,乳酸大量堆積導致培養基內pH 下降,乳酸菌生長受到抑制。無機鹽可以調節培養基的pH,中和過量的酸,促進菌體的生長,并為其生長補充微量元素。因此選擇9 g/L 作為磷酸氫二鉀的最適添加量。
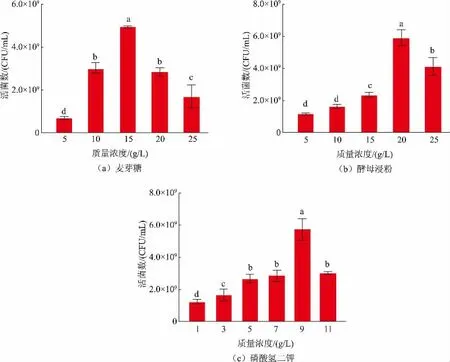
圖2 不同質量濃度麥芽糖、酵母浸粉、磷酸氫二鉀對發酵乳桿菌SS-31 生長的影響Fig.2 Effects of different concentrations of maltose,yeast extract powder and potassium hydrogen phosphate on the growth of L.fermentum SS-31
2.3 響應面實驗結果
利用Design-Expert 10.0 軟件對麥芽糖質量濃度(A)、酵母浸粉質量濃度(B)、磷酸氫二鉀質量濃度(C)設計三因素三水平的Box-Behnken 實驗。實驗設計及結果見表2。
通過對表2的實驗結果進行二次項擬合,以L.fermentumSS-31 活菌數作為響應值,對結果進行多元回歸分析,得到回歸方程:活菌數(×109CFU/mL)=6.920+0.559A-0.014B+0.532C+0.786AB+0.636AC-0.467BC-1.440A2-1.534B2-2.028C2。
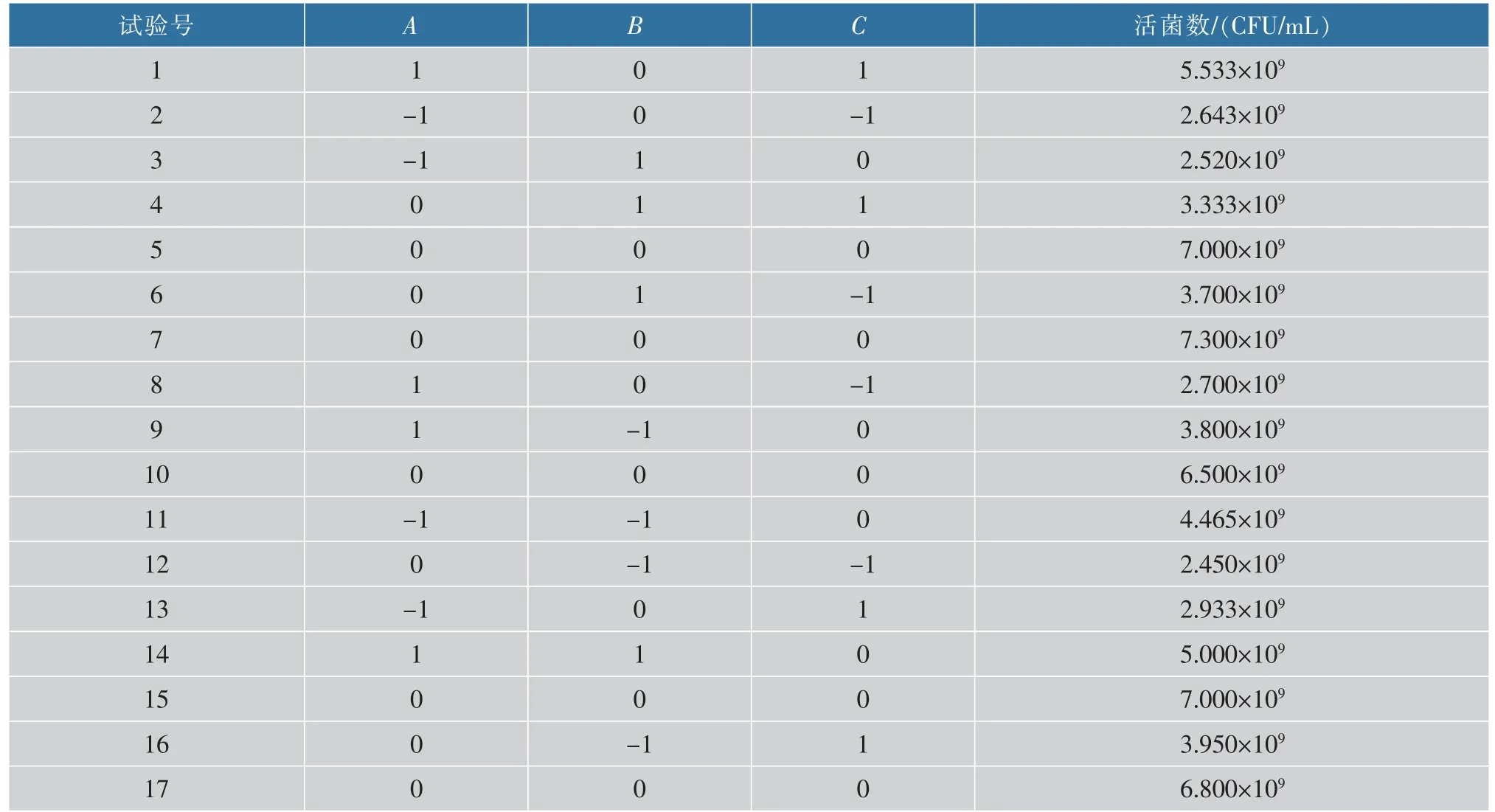
表2 Box-Behnken 實驗設計及結果Table 2 Box-Behnken design and its experimental results
由表3可知模型P<0.000 1,表明該模型建立成功;失擬項為0.148 6,大于0.05,說明失擬項不顯著;模型的R2=0.977 0,Radj2=0.947 5,說明預測值和實驗值的相關性高,可以解釋94.75%的響應值變化。該模型變異系數(0.73%)小,具有較高精確度。綜上,該回歸模型可對L.fermentumSS-31 在培養基的生長情況進行有效分析和預測。3 個因素交互作用的響應曲面及等高線見圖3。
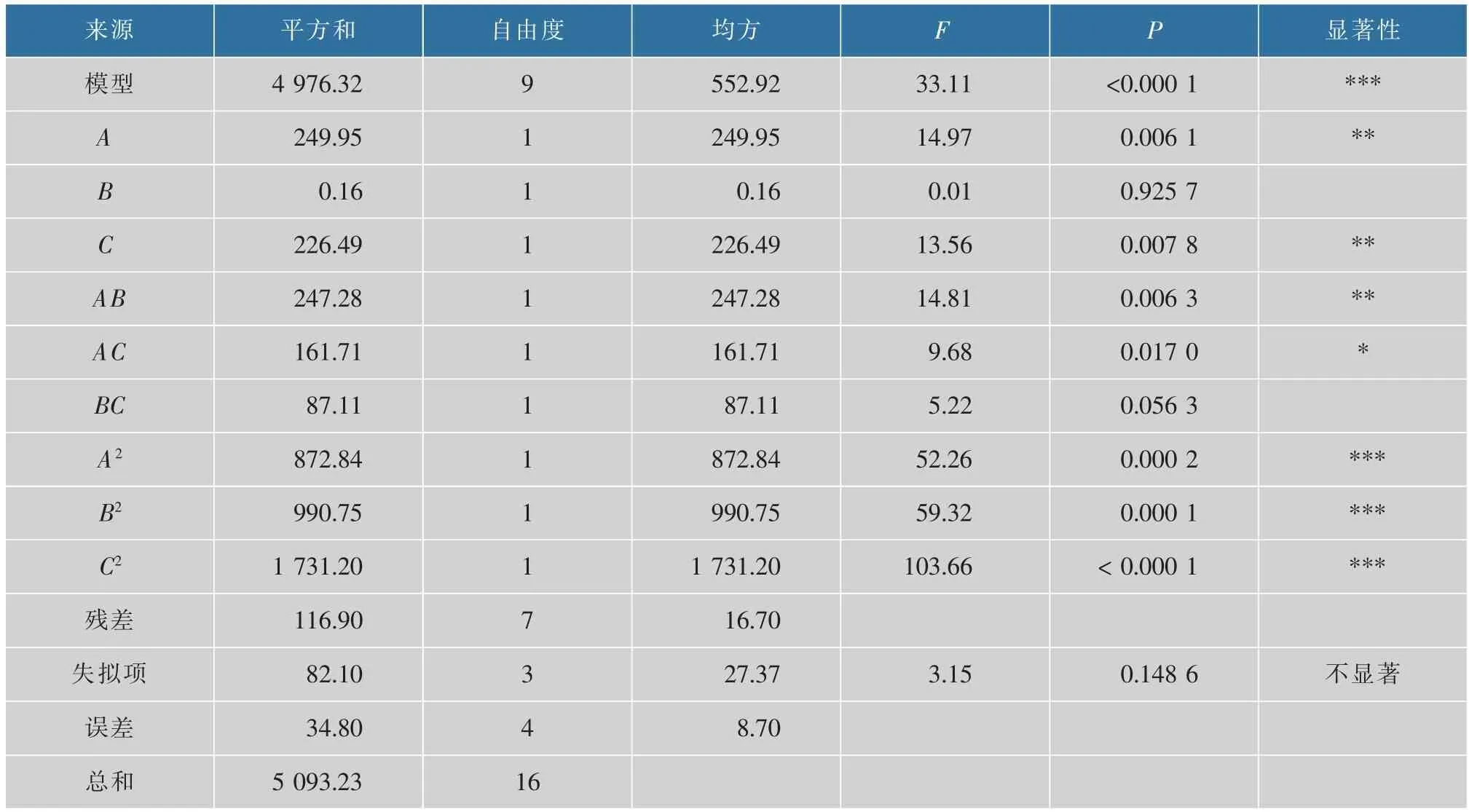
表3 RSM 回歸方程方差分析Table 3 Variance analysis of RSM regression model
2.4 響應面實驗結果的驗證
回歸模型得出Box-Behnken 實驗設計的最優結果見表4。在麥芽糖16.20 g/L、酵母浸粉20.16 g/L、磷酸氫二鉀9.33 g/L 時,在此優化條件下的活菌數預測值為7.03×109CFU/mL。為了驗證模型所得優化結果的準確性,依照優化結果進行培養基的配制及發酵實驗,測定24 h 時L.fermentumSS-31 的活菌數,3 次實驗結果的平均值為6.51×109CFU/mL,和模型預測值接近,驗證了此模型的準確性。

表4 Box-Benhnken 驗證實驗與結果Table 4 Box-Benhnken verification experiments and results
楊瑞冬等對Lactobacillus buchneriIMAU80233培養基中碳源、氮源、無機鹽體系進行Box-Behnken實驗,后經發酵培養條件優化后,菌體活菌數僅有3.81×109CFU/mL,約為優化前的1.59 倍[21];李納對鼠李糖乳桿菌RG2 培養基成分及培養條件優化后,活菌數達到7.1×109CFU/mL,是未優化前活菌數的7 倍[22];高志敏采用正交法結合響應面法優化Lactobacillus fermentumIMAU 10129 的培養基,優化后活菌數為4.21×109CFU/mL,為優化前活菌數的6 倍[23]。而在該實驗中,優化后L.fermentumSS-31活菌數為6.51×109CFU/mL,是優化前MRS 基礎培養基活菌數(4.67×108CFU/mL)的13.94 倍,活菌數提升了一個數量級,高于目前文獻已報道的活菌數水平,說明效果較好。
2.5 高密度培養條件的優化
2.5.1 接種體積分數的選擇菌體接種體積分數的多少通常影響其生長到穩定期的時間以及濃度[24]。作者探究了不同接種體積分數對L.fermentumSS-31活菌數的影響,結果見圖4。在接種體積分數為1%~5%時,L.fermentumSS-31 的活菌數隨接種體積分數升高呈現先上升后下降的趨勢,在接種體積分數為3%時,活菌數達到最大值6.83×109CFU/mL。這可能是初始接種體積分數過大,菌體大量繁殖消耗更多營養物質,導致后期培養基內營養物質不足,從而呈現下降趨勢。綜上,選擇3%為L.fermentumSS-31 的接種體積分數。
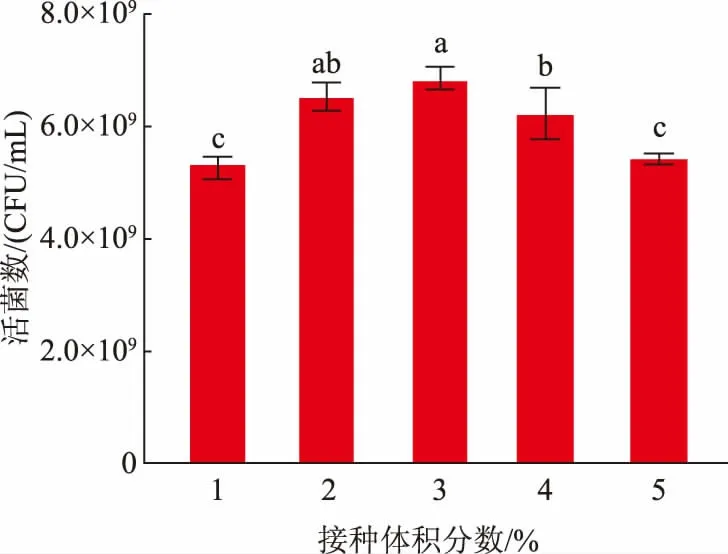
圖4 接種體積分數對發酵乳桿菌SS-31 生長的影響Fig.4 Effects of inoculum on the growth of L.fermentum SS-31
2.5.2 初始pH 的選擇乳酸菌的最適生長pH 一般為5.0~7.0,不同種類乳酸菌有不同的最適生長pH。作者探究了不同初始pH 對L.fermentumSS-31活菌數的影響,結果見圖5。隨著初始pH 的增加,L.fermentumSS-31 的活菌數呈現先增加后降低的趨勢,在pH 為6.8 時,活菌數達到最大值7.30×109CFU/mL。在過酸的環境下,會破壞細胞膜滲透壓平衡和干擾細胞內外酶的活性,限制乳酸菌的生長;當pH 過高時,也不利于其生長[25-26]。綜上,選擇pH 6.8 為初始pH。
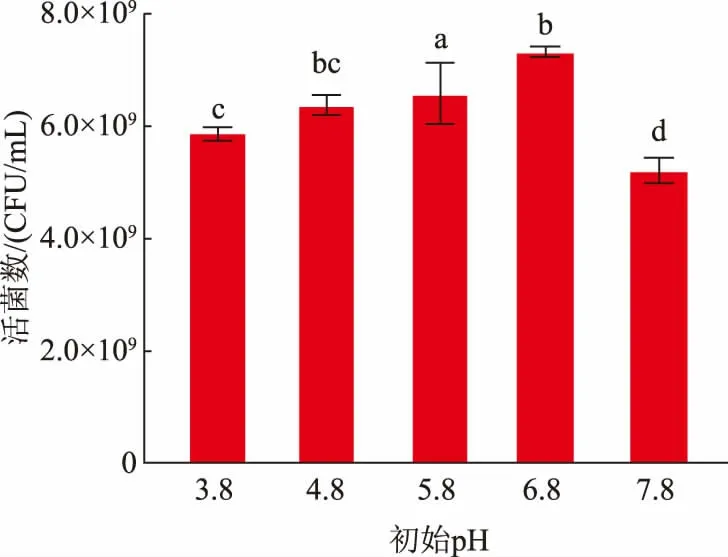
圖5 初始pH 對發酵乳桿菌SS-31 生長的影響Fig.5 Effects of initial pH on the growth of L.fermentum SS-31
2.5.3 溫度的選擇溫度的變化對乳酸菌的影響較大,溫度過高或過低均不利于其生長繁殖。作者探究了不同溫度對L.fermentumSS-31 活菌數的影響,結果見圖6。隨著溫度升高,L.fermentumSS-31的活菌數呈現顯著增加后降低的趨勢,在溫度為37 ℃時,活菌數達到最大值8.15×109CFU/mL。這可能因為在低溫條件下菌體失活,高溫使其酶失活,從而影響其生長速度。綜上,選擇37 ℃為發酵乳桿菌SS-31 的最佳培養溫度。

圖6 溫度對發酵乳桿菌SS-31 生長的影響Fig.6 Effects of temperature on the growth of L.fermentum SS-31
2.5.4 中和劑的選擇分批培養菌體時,抑制菌體生長的主要原因是菌體自身代謝會產生較多的有機酸,大量有機酸的累積會形成酸抑制,從而使菌體生長較為緩慢。但是通過在發酵過程中加入中和劑調節pH,保持恒定pH 則可以解除酸抑制[27-28]。作者利用15 L 發酵罐探究了不同中和劑對L.fermentumSS-31 活菌數的影響,結果見圖7。
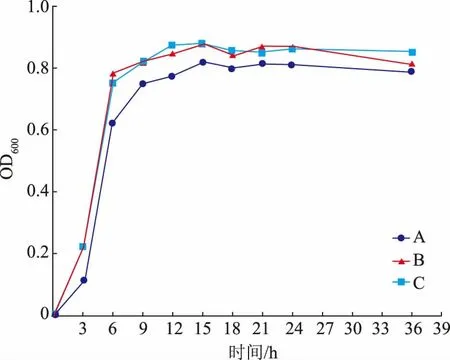
圖7 中和劑對發酵乳桿菌SS-31 生長的影響Fig.7 Effects of neutralizer type on the growth of L.fermentum SS-31
如圖7所示,與未添加中和劑組相比,當添加碳酸鈉和氨水,使發酵液pH 恒定為6.8 后,L.fermentumSS-31 菌液OD600顯著增加(P<0.05);但氨水和碳酸鈉作為中和劑時,二者對L.fermentumSS-31 生長的影響無顯著性差異 (P>0.05)。氨水作為中和劑時,既可以調節培養基內的pH,又可以為菌體生長提供氮源,用量比碳酸鈉少,且氨水較為廉價,經濟實惠。因此,選用氨水作為L.fermentumSS-31 高密度培養的中和劑,此時37 ℃培養24 h 后活菌數達到1.19×1010CFU/mL。
另外,孫媛媛對發酵乳桿菌FXJCJ6-1 最適生長pH 進行研究,結果顯示在pH 恒定為6.0 并分批培養時,活菌數達到9.5×109CFU/mL,較初始MRS培養基中活菌數(1.7×109CFU/mL)提高了4.6 倍[27];李江等對發酵乳桿菌L1 高密度培養條件進行優化,結果顯示通過流加中和劑NaOH,維持恒定pH,較分批培養時的活菌數(1.55×1010CFU/mL)顯著提高18.52 倍[29];聶黔麗等采用單因素實驗和正交實驗對發酵乳桿菌LF-8001 進行高密度培養優化后,活菌數達到了1.08×1010CFU/mL,提高了5.32 倍[24]。作者對L.fermentumSS-31 進行培養基和培養條件優化后,活菌數達到1.19×1010CFU/mL,相比優化前活菌數(4.67×108CFU/mL)提高了25.48 倍,活菌數提升了兩個數量級,高于目前大部分已報道的活菌數水平,說明效果良好。
3 結 語
實現乳酸菌高密度培養的核心是發酵培養基的優化,作者以活菌數為指標,通過對L.fermentiumSS-31 發酵培養基組成和發酵條件進行系統優化,最終確定最佳發酵培養基成分為:麥芽糖16.20 g/L、酵母浸粉20.16 g/L、磷酸氫二鉀9.33 g/L、硫酸錳0.50 g/L、硫酸鎂1.00 g/L、吐溫80 1.00 g/L;最佳發酵條件為:接種體積分數3%、初始pH 6.8,期間添加氨水保持發酵液pH 穩定,在37 ℃下培養24 h后,活菌數可達到1.19×1010CFU/mL,是優化前4.67×108CFU/mL 的25.48 倍。發酵乳桿菌SS-31 菌體濃度顯著提高,為今后大規模短期快速生產提供了技術支持。
猜你喜歡
房地產導刊(2022年5期)2022-06-01 06:20:14
建材發展導向(2021年12期)2021-07-22 08:06:48
建材發展導向(2021年7期)2021-07-16 07:07:52
中學生數理化(高中版.高二數學)(2021年12期)2021-04-26 07:43:48
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化(高中版.高考數學)(2021年12期)2021-03-08 01:28:50
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
