鱸魚(Lateolabrax japonicus)營養需求與飼料添加劑研究進展(二)
2023-01-06 02:08:04作者劉敏王卓鐸孫廣文張海濤
廣東飼料 2022年10期
◆作者:劉敏 王卓鐸 孫廣文 張海濤*
◆單位:1.廣東恒興飼料實業股份有限公司,農業農村部華南水產與畜禽飼料重點實驗室 2.湛江恒興動物營養研發有限公司;3.湛江恒興特種飼料有限公司
續上期
2 鱸魚的脂類營養
脂肪是魚類生長發育的重要能量來源,可為魚類提供其自身不能合成的必需脂肪酸,可改善飼料適口性。脂肪是魚類所必需的營養物質,飼料中脂肪含量須適宜。
2.1 適宜脂肪水平
研究表明,體重在9.52~343.3g 的鱸魚的日糧脂肪需求范圍在7.40%~12.91%(表4)。淡水養殖中日糧脂肪水平超過12%會抑制鱸魚生長,同時顯著增加HSI、VSI 和全魚脂肪含量。肝臟脂肪含量直接受日糧脂肪水平的影響,但肌肉脂肪含量保持相對不變(Luo 等,2010)。海水養殖中日糧脂肪水平超過15.8%不能改善實驗魚的生長性能或飼料利用率,但會產生全魚高脂沉積(Xu 等,2011)。此兩項研究均表明DHA 可以選擇性地保留在鱸魚肌肉中。日糧脂肪水平從0.6%提高到7.3%,SGR 顯著提高;從7.3%提高到15.8%,SGR有所下降。隨著日糧脂肪水平的增加,整個魚體和肝臟中的脂肪含量顯著增加。鱸魚成長過程中的脂肪需求、身體成分和脂肪沉積因體型不同而不同(Xu 等,2015)。129.1~178.6 g/kg 的日糧脂肪水平會降低鱸魚的生長性能,并誘導其脂質積累、炎癥和細胞凋亡(Xie 等,2021)。

表4 鱸魚的脂肪需求(%飼料干重)
2.2 脂肪酸需求(見表5)
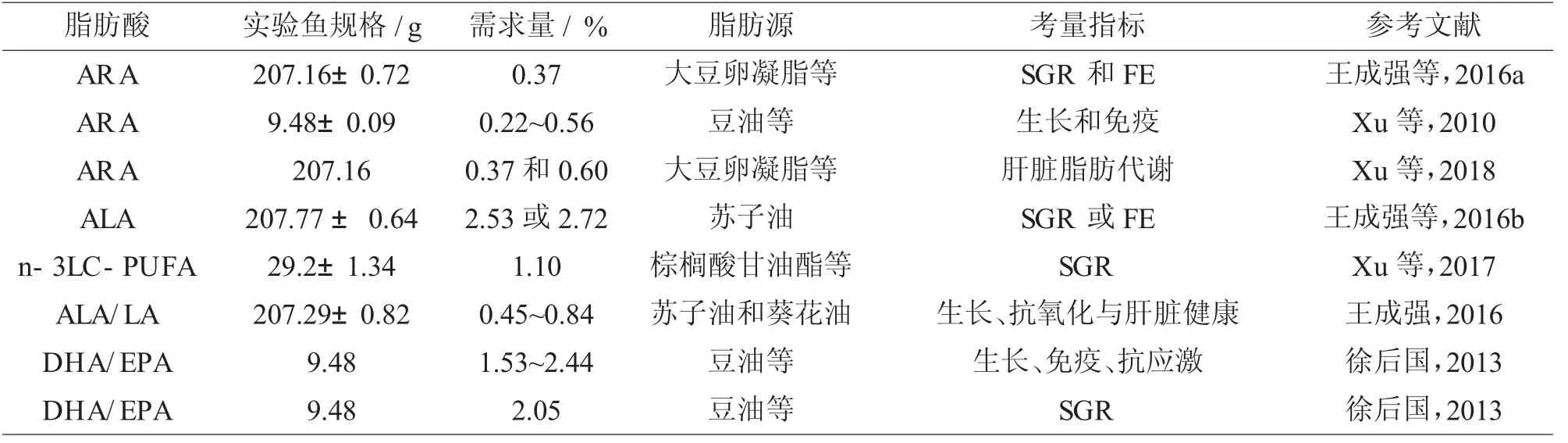
表5 鱸魚的脂肪酸需求(%飼料干重)
研究表明,日糧中0.08%~2.12%的花生四烯酸(ARA)水平,尤其是中等水平(0.22%~0.56%)顯著增強了小規格鱸魚的生長和免疫反應,并改變了全魚和肝臟的化學組成(Xu 等,2010)。日糧中0.05%~2.32%的ARA 對SR、HSI、VSI、肥 滿 度(CF)均無顯著性影響。0.37%和0.60%的ARA 水平可減少大規格鱸魚肝臟中的脂質積累。0.37%的ARA 水平顯著降低了血清谷草轉氨酶和谷丙轉氨酶活性(王成強等,2016a;Xu 等,2018)。王成強等(2016b)同時探討了飼料中不同水平(0.06%、0.99% 、2.03% 、3.18% 、4.12% 、5.08%)的亞麻酸(ALA)對大規格鱸魚生長性能、抗氧化指標和血清生化指標的影響。綜合分析,飼料中適宜含量(2.03%~3.18%)的ALA 能夠促進大規格鱸魚的生長,提高抗氧化能力與肝臟健康水平。Xu 等(2017)有關日糧中1.30~24.13 g/kg 的n-3 LC-PUFA 對鱸魚的影響的研究表明,10.31 g/kg 的日糧n-3 LC-PUFA 獲得了最高SGR,并有利于提高血清中LZM 的活性。隨著日糧n-3 LC-PUFA 的增加,魚體粗脂肪含量顯著降低。同時,24.13 g/kg 的日糧n-3 LC-PUFA 使得肝臟脂質含量顯著增加。肝臟總脂肪酸中n-3 LC-PUFA 含量與日糧中n-3 LC-PUFA 含量密切相關。血清谷丙轉氨酶和谷草轉氨酶活性無顯著差異。有關含不同特征脂肪酸(棕櫚酸、硬脂酸、油酸、α-亞麻酸、n-3 LC-PUFA)的5 種日糧對海水養殖的鱸魚[(29.53±0.86)g] 組織中LC-PUFA 含量和生物合成的影響的研究表明,鱸魚合成LC-PUFA 的能力較低,攝食缺乏LC-PUFA的日糧導致魚組織n-3 LC-PUFA含量顯著降低(Xu 等,2014)。
此外, 飼料中適量的ALA/LA 比值能顯著提高鱸魚的生長、抗氧化能力與肝臟健康水平,過低或過高ALA 和LA 含量均會一定程度影響肝臟脂肪酸合成和氧化相關基因的表達(王成強,2016)。飼料中適量的DHA/EPA 比例能顯著提高鱸魚幼魚的生長、免疫性能和抗應激能力,并遵循DHA 優先積累于魚組織的規律(徐后國,2013;Xu等,2016)。
此外,呈現鱸魚肌肉中脂肪酸的組成(表6),以供研究者參考(吳燕燕等,2016)。
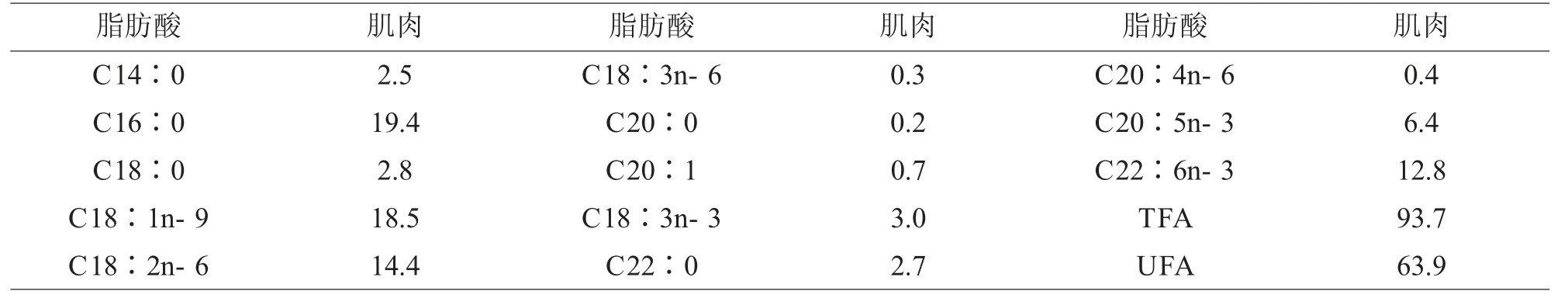
表6 鱸魚肌肉的脂肪酸組成(%)
2.3 脂肪源替代(見表7)

表7 鱸魚配合飼料中的魚油替代研究
潘瑜等(2016)研究了裂殖壺菌粉等DHA 含量替代0%、50%和100%的魚油,并補充豆油,對鱸魚生長性能、肌肉常規組成及抗氧化能力的影響。綜合分析發現,裂殖壺菌粉能夠替代50%的魚油而不影響生長,但完全替代魚油會造成魚體生長受阻,并危及魚體健康。脂肪水平為10%時,分別以6 種替代脂肪源[豬油、牛油、雞油、豆油、玉米油、混合脂肪(60%牛脂+20%豆油+20%魚油)] 替代對照組中50%的魚油,不會降低鱸魚生長性能,但魚片的脂肪酸結構會發生改變,并且與日糧的脂肪酸結構密切相關(Xue 等,2006)。綜合WG、SGR、FCR、脂肪酸組成等指標分析發現,棕櫚油可以替代鱸魚幼魚日糧中50%的新鮮魚油。雖然用棕櫚油替代氧化魚油(POV:26 meq/kg日糧)可減少魚腸道和肌肉中的脂質過氧化,但不能改善魚的生長性能(Gao等,2012)。用亞麻籽油或豆油替代1/3、2/3 和3/3 的魚油不會降低鱸魚的生長性能,但會降低免疫反應和LC-PUFA 濃度。就鱸魚的生長性能和免疫反應而言,相比豆油,亞麻籽油是更好的脂質來源(Xu 等,2015)。在滿足鱸魚對日糧n-3 LC PUFA 需求的情況下,以亞麻籽油和豬油(1∶1)完全替代日糧魚油,對SR、體成分、體指標、非特異性免疫或應激耐受性均沒有顯著影響,但顯著提高了SGR 和FER,顯著降低了血清抗氧化酶活性和肝組織抗氧化相關基因表達(Xue等,2021)。
此外,Tan 等(2017)的研究發現,分別用植物油(豆油∶亞麻 籽 油 =1 ∶1) 替 代 鱸 魚(18.60±0.36 g)日糧中0%、50%、100%的魚油,降低了鱸魚酸性磷酸酶活性、抗病性和肝臟抗氧化能力,加劇了炎癥反應。Xu 等(2019) 研究了脂肪源分別為10%葵花籽油(富含亞油酸)或10%紫蘇油(富含α- 亞麻酸)的日糧,結果發現,鱸魚組織亞油酸和α- 亞麻酸組成與其所攝食日糧的亞油酸和α- 亞麻酸組成相對應,但兩組間鱸魚的生長性能和身體近似成分沒有顯著差異。α- 亞麻酸和亞油酸不同程度地調節了鱸魚中一系列與脂質和蛋白質代謝相關的生物過程。高水平的日糧α- 亞麻酸傾向于在基因表達水平上抑制鱸魚肝臟中的脂質運輸和蛋白質生物合成過程。魚油氧化會顯著降低鱸魚[(8.01±0.76)g]的生長速度,補充維生素E(600IU/kg)可以減輕或補償氧化魚油的負面影響,提高SR 和生長性能。飼料中魚油氧化的程度會對維生素E 的作用產生影響,在研究鱸魚飼料中維生素E 最適添加量時,需要考慮飼料脂類珺氧化情況(王珺,2014)。
3 鱸魚的碳水化合物營養
碳水化合物是最廉價的能源,在飼料中合理使用可降低飼料成本,改善飼料蛋白的利用,部分糖類可改善飼料性狀。
竇兵帥等(2014)關于飼料中不同碳水化合物水平(0、6%、12%、18%、24%、30%) 對鱸魚[(34.26±0.37)g]生長、飼料利用、血糖水平和糖酵解酶活力的影響的研究表明,在不影響鱸魚正常生長、消化與機體代謝的情況下,SGR 經二次曲線模型分析得出鱸魚飼料中碳水化合物的適宜 水 平 為17.75% 。 程 鎮 燕(2010)通過在基礎飼料中分別添加不同水平的糊化玉米淀粉,配制成糖實際含量分別為1.86%、7.34%、13.56%、19.40%、24.82%、31.76%的6 種鱸魚[(8.00±0.20)g]實驗飼料。結果表明,飼料中糖含量對鱸魚SR 沒有顯著影響,但顯著提高了鱸魚粗蛋白和粗脂肪含量,并顯著降低了魚體水分和灰分含量。肝糖原和血糖含量隨著飼料糖含量的增加而呈現顯著的先上升后平穩的趨勢,肌糖原含量并沒有受到飼料糖水平的影響。飼料中添加糖顯著提高了鱸魚肝臟蛋白酶和淀粉酶活性,對己糖激酶活力沒有顯著影響。隨著飼料中糖含量的增加,鱸魚對飼料蛋白質、脂肪和糖的消化率顯著降低,當飼料中糖含量大于19.40%時,鱸魚對飼料蛋白質、脂肪和糖的消化率顯著低于對照組。以SGR 經二次曲線模型分析得出鱸魚飼料中糖的適宜含量約為19.8%。
Zheng 等(2015)研究了六個水平(0%、6%、12%、18%、24%、30%)的玉米淀粉對生長期鱸魚(初重343g)生長性能、體成分、肝臟/肌肉糖元等指標的影響。體蛋白和脂質含量、血清中總膽固醇、甘油三酯均隨日糧玉米淀粉水平的增加先增加,然后達到穩定。隨著日糧玉米淀粉的增加,灰分含量先降低,然后達到一個穩定值;HSI 和肝糖元顯著增加,而肌糖元不受日糧玉米淀粉水平的影響;血糖先升高后降低。以WG 為考量依據,經二次多項式模型分析得出鱸魚日糧中玉米淀粉的最佳水平為27.0%。Peng 等(2020)研究了2種碳水化合物水平(15%、39%)以及39%的碳水化合物水平下添加0.1%的縮合單寧對鱸魚[(2.3±0.01)g]的影響。結果表明,日糧碳水化合物達39%時會降低鱸魚的生長性能、增強抗氧化和免疫能力以及提高血糖。添加0.1%的縮合單寧可改善生長性能并降低血糖濃度,這主要是因為縮合單寧對鱸魚肝糖酵解酶和糖異生酶的活性和基因表達有積極影響。
(未完,待續見下期)
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小讀者(2021年2期)2021-03-29 05:03:48
火花(2019年12期)2019-12-26 01:00:28
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
人大建設(2019年12期)2019-05-21 02:55:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
