無患子RT-qPCR內(nèi)參基因的篩選與驗(yàn)證
2023-01-05 08:45:32徐圓圓趙國春郝穎穎翁學(xué)煌陳仲賈黎明
生物技術(shù)通報(bào) 2022年10期
徐圓圓 趙國春 郝穎穎 翁學(xué)煌 陳仲,5 賈黎明
(1.北京林業(yè)大學(xué)林學(xué)院省部共建森林培育與保護(hù)教育部重點(diǎn)實(shí)驗(yàn)室,北京 100083;2.北京林業(yè)大學(xué)國家能源非糧生物質(zhì)原料研發(fā)中心,北京 100083;3.北京林業(yè)大學(xué)無患子產(chǎn)業(yè)國家創(chuàng)新聯(lián)盟,北京 100083;4.福建源華林業(yè)生物科技有限公司,三明 354500;5.北京林業(yè)大學(xué)林木分子設(shè)計(jì)育種高精尖創(chuàng)新中心,北京 100083)
實(shí)時(shí)熒光定量PCR(RT-qPCR)是一種研究植物生長發(fā)育和代謝物生物合成過程中關(guān)鍵基因功能的重要技術(shù),由于其準(zhǔn)確性高、特異性強(qiáng)、操作簡(jiǎn)便的特點(diǎn),常被應(yīng)用于基因的表達(dá)分析[1-2]。樣品的RNA完整性、cDNA質(zhì)量、引物特異性及擴(kuò)增效率等因素都會(huì)影響RT-qPCR實(shí)驗(yàn)結(jié)果[1,3],引入1個(gè)或多個(gè)表達(dá)穩(wěn)定的內(nèi)參基因?qū)δ康幕蜻M(jìn)行校正和標(biāo)準(zhǔn)化可以提高RT-qPCR結(jié)果的準(zhǔn)確性[4]。常用的內(nèi)參基因大多是維持細(xì)胞正常生命代謝的管家基因,如與細(xì)胞結(jié)構(gòu)相關(guān)的肌動(dòng)蛋白基因(actin)、微管蛋白基因(tublin),與蛋白質(zhì)降解相關(guān)的泛素基因(ubiquitin),與糖代謝相關(guān)的甘油醛-3-磷酸-脫氫酶基因(GAPDH)以及編碼核糖體RNA的基因18S rRNA、28S rRNA等[5-6]。理想的內(nèi)參基因應(yīng)該是在所有生理狀態(tài)下的樣品、組織類型中均能較穩(wěn)定表達(dá)的基因[7]。然而,研究表明植物中并不存在絕對(duì)穩(wěn)定表達(dá)的基因,不同物種、組織、發(fā)育階段或試驗(yàn)條件下,內(nèi)參基因的表達(dá)穩(wěn)定性存在差異,例如,千里光(Senecio scandens)的內(nèi)參基因ACT1在不同組織和不同脅迫處理下表達(dá)穩(wěn)定[8],而老鴉瓣(Amana edulis)的ACT在不同器官和不同發(fā)育時(shí)期的花芽中的表達(dá)穩(wěn)定性卻較差[9];紅花(Carthamus tinctorius)的UBCE2是最適合用于種子發(fā)育階段基因表達(dá)研究的內(nèi)參基因,而在脫落酸處理下,研究不同紅花品種對(duì)脫落酸信號(hào)響應(yīng)的最佳內(nèi)參基因是UBCE2和EF1β組合[10]。因此,根據(jù)物種、試驗(yàn)條件和試驗(yàn)?zāi)康倪x擇合適的內(nèi)參基因是獲得準(zhǔn)確、可靠的 RT-qPCR 實(shí)驗(yàn)結(jié)果的關(guān)鍵[7,11]。
無患子Sapindus mukorossi Gaertn.為無患子科(Sapindaceae)無患子屬(Sapindus Linn.)植物,是我國傳統(tǒng)的洗護(hù)用植物和藥用植物,集日用化工、生物醫(yī)藥、生物質(zhì)能源、綠化美化和生態(tài)修復(fù)于一體,具有較高的綜合利用價(jià)值[12]。我們前期研究表明無患子根、莖、葉、花和果皮中均含有三萜皂苷[13],三萜皂苷起泡性能和去污能力強(qiáng),可以用于制作手工皂、洗發(fā)水等洗護(hù)用品[14]。此外,無患子三萜皂苷及其皂苷元還具有抑菌、抗腫瘤、降血糖、降血脂、保肝和殺蟲等多種生物活性,可用于醫(yī)藥、食品保健和農(nóng)業(yè)等領(lǐng)域[15]。目前,對(duì)無患子的研究主要集中在種質(zhì)資源收集和評(píng)價(jià)[14,16]、開花結(jié)實(shí)習(xí)性[17],原料林高效培育[18]及化學(xué)成分的提取、分離、鑒定與應(yīng)用[19-20]等方面,極少涉及分子生物學(xué)和功能基因組學(xué)方面的研究,無患子內(nèi)參基因篩選方面的研究也未見報(bào)道。課題組已完成了無患子根、莖、葉、芽、花和不同發(fā)育時(shí)期果皮的轉(zhuǎn)錄組測(cè)序工作,并從中挖掘了一系列無患子三萜皂苷生物合成候選基因。本研究從轉(zhuǎn)錄組測(cè)序數(shù)據(jù)中初步篩選了8個(gè)常見的候選內(nèi)參基因,通過RT-qPCR技術(shù),結(jié)合GeNorm、NormFinder、BestKeeper三個(gè)分析軟件和RefFinder在線分析工具,對(duì)它們?cè)跓o患子不同器官和不同發(fā)育時(shí)期果皮中的表達(dá)穩(wěn)定性進(jìn)行評(píng)估,以期篩選適用于無患子三萜皂苷生物合成相關(guān)基因表達(dá)研究的內(nèi)參基因,同時(shí)對(duì)無患子三萜皂苷生物合成過程中的6個(gè)編碼關(guān)鍵酶的基因(乙酰輔酶A酰基轉(zhuǎn)移酶基因,SmAACT;1-脫氧-木酮糖-5-磷酸合酶基因,SmDXS;法呢基焦磷酸合成酶基因,SmFPS;β-香樹素合成酶基因,SmbAS1;細(xì)胞色素P450酶基因,SmCYP716A-5;糖基轉(zhuǎn)移酶基因,SmUGT73C-14)和2個(gè)編碼轉(zhuǎn)錄因子的基因(SmbHLH8和SmERF25)的表達(dá)模式進(jìn)行分析,可為后續(xù)深入開展無患子三萜皂苷生物合成機(jī)理研究奠定基礎(chǔ),也可為無患子及近緣植物的其他生物學(xué)過程中的基因表達(dá)研究提供參考。
1 材料與方法
1.1 材料
試驗(yàn)材料采自福建省三明市建寧縣無患子原料林(116°52′E,26°49′N)。于 2018 年 3-11 月進(jìn)行試驗(yàn)材料的采集。在果園中選取3株樹體相近、生長健康、結(jié)果量穩(wěn)定的優(yōu)樹進(jìn)行果實(shí)樣品采集,以單株為小區(qū),3次重復(fù)。于3月4日采集芽(bud,B),4月 25日采集根(root,R)、莖(stem,ST)、葉(leaf,L),5月25日采集雄花(male flower,MF)和雌花(female flower,MF);6月10日-11月10日采集完整果實(shí)發(fā)育周期的果皮樣品,涵蓋果實(shí)發(fā)育全程的8個(gè)關(guān)鍵時(shí)期[17,21],分別為:子房發(fā)育早期(花后15 d,S1);果實(shí)最大體積的30%(花后45 d,S2);果實(shí)最大體積的70%(花后75 d,S3);果實(shí)最大體積的80%(花后90 d,S4);果實(shí)最大體積的90%(花后105 d,S5);果實(shí)開始成熟(花后120 d,S6);果皮發(fā)生較大變化(花后135 d,S7);果實(shí)充分發(fā)育和成熟(花后150 d,S8)(圖1)。每個(gè)時(shí)期于晴天上午10點(diǎn)進(jìn)行采樣,每株樹在相同位置上隨機(jī)采摘果實(shí),果實(shí)采摘后立即取下果皮,用液氮速凍后,置于-80℃保存?zhèn)溆谩?/p>

圖1 無患子采樣材料Fig.1 Sample materials of S.mukorossi
FinePure Plant RNA提取試劑盒購自北京濟(jì)凡生物科技有限公司;Goldenstar RT6 cDNA Synthesis Kit Ver 2、2× T5 Fast qPCR Mix等購自北京擎科生物科技有限公司。
1.2 方法
1.2.1 候選內(nèi)參基因的選取 基于無患子根、莖、葉、芽、花和果皮的轉(zhuǎn)錄組數(shù)據(jù),以基因表達(dá)量FPKM(fragments per kilobase of transcript per million mapped reads)值大于10和不同器官或果皮發(fā)育時(shí)期間FPKM的差異倍數(shù)小于2為篩選標(biāo)準(zhǔn),選取8個(gè)常用的內(nèi)參基因作為候選基因進(jìn)行穩(wěn)定性分析,包括18S核糖體RNA甲基轉(zhuǎn)移酶基因(18S rRNA(guanine1575-N7)-methyltransferase,Sm18S)、 肌動(dòng)蛋白基因(actin-related protein 8,SmACT)、轉(zhuǎn)錄延伸因子基因(elongation factor 1-alpha,SmEF-1α)、大亞基核糖體蛋白基因(large subunit ribosomal protein L1,SmRPL1)、小亞基核糖體蛋白基因(small subunit ribosomal protein S26e,SmRPS26)、微管蛋白特異性伴侶蛋白基因(tubulin-specific chaperone C,SmTBCC)、泛素綴合酶基因(ubiquitin-conjugating enzyme E2 M,SmUBC12)、E3泛素蛋白連接酶基因(E3 ubiquitin-protein ligase BAH,SmUBP)。根據(jù)候選內(nèi)參基因的核苷酸序列,使用Primer Premier 5.0軟件設(shè)計(jì)引物(表1),引物由北京擎科生物科技有限公司合成。
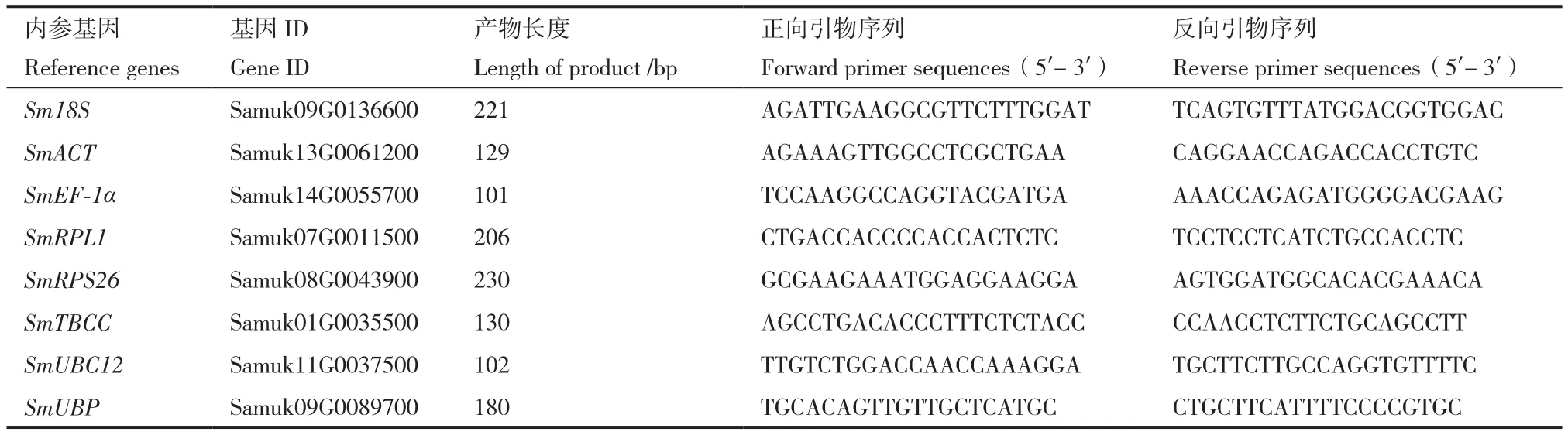
表1 候選內(nèi)參基因的引物Table 1 Primers of candidate reference genes
1.2.2 候選內(nèi)參基因的RT-qPCR分析 根據(jù)FinePure Plant RNA提取試劑盒說明書提取無患子樣品的總RNA,并采用1%瓊脂糖凝膠電泳檢測(cè)每個(gè)樣品的RNA質(zhì)量,然后使用NanoDrop 2000分光光度計(jì)檢測(cè)RNA的濃度及純度,要求A260/A280值在1.8-2.1之間。使用反轉(zhuǎn)錄試劑盒Goldenstar RT6 cDNA Synthesis Kit Ver 2合成cDNA。使用LineGene 9600 Plus熒光定量聚合酶反應(yīng)(PCR)檢測(cè)系統(tǒng)(杭州博日,中國)進(jìn)行熒光定量檢測(cè)。擴(kuò)增體系和反應(yīng)程序如下:反應(yīng)體系為20 μL,其中T5 Fast qPCR Mix(SYBR Green I)(2×)10 μL,正反向引物各0.8 μL,cDNA 模板 1 μL,ddH2O 7.4 μL。PCR 程序 :95℃預(yù)變性 1 min;95℃變性 15 s,60℃退火 34 s,72℃延伸30 s,40個(gè)循環(huán);反應(yīng)結(jié)束后通過熔解曲線檢測(cè)產(chǎn)物特異性:從60℃緩慢升溫(0.11℃/s)至95℃,每升溫1℃連續(xù)采集5次熒光信號(hào)。每個(gè)樣本設(shè)置3次技術(shù)重復(fù)。最后基于擴(kuò)增結(jié)果制作內(nèi)參基因的標(biāo)準(zhǔn)曲線,并計(jì)算相關(guān)系數(shù)。
1.2.3 候選內(nèi)參基因的穩(wěn)定性分析 使用Microsoft Excel 2016軟件對(duì)實(shí)時(shí)熒光定量PCR產(chǎn)生的原始Ct值進(jìn)行整理,計(jì)算每個(gè)樣本的平均Ct值。使用 geNorm[22]、NormFinder[23]和 BestKeeper[24]軟件對(duì)8個(gè)候選內(nèi)參基因的穩(wěn)定性進(jìn)行分析。其中,geNorm和NormFinder軟件以2-△Ct進(jìn)行分析,△Ct=Ct樣本-Ctmin;geNorm軟件通過計(jì)算候選內(nèi)參基因的平均表達(dá)穩(wěn)定值(M)對(duì)其穩(wěn)定性進(jìn)行評(píng)估,該值與內(nèi)參基因的穩(wěn)定性呈負(fù)相關(guān)關(guān)系,一般認(rèn)為M值小于1.5,內(nèi)參基因表達(dá)穩(wěn)定[22]。另外,geNorm軟件還可以通過候選內(nèi)參基因標(biāo)準(zhǔn)化因子的配對(duì)差異分析(Vn/Vn+1)得出最佳內(nèi)參基因的配對(duì)組合。軟件默認(rèn)的Vn/Vn+1值為0.15,當(dāng)Vn/Vn+1<0.15時(shí),說明n個(gè)內(nèi)參基因已滿足穩(wěn)定歸一化;當(dāng)Vn/Vn+1>0.15時(shí),則需要n+1個(gè)內(nèi)參基因才能滿足穩(wěn)定歸一化。NormFinder軟件則以表達(dá)穩(wěn)定值(Stability value,SV)對(duì)候選內(nèi)參基因的表達(dá)穩(wěn)定性進(jìn)行評(píng)價(jià),該值越小,說明基因表達(dá)越穩(wěn)定[23]。Bestkeeper軟件以樣本的原始Ct值進(jìn)行分析,通過Ct值的標(biāo)準(zhǔn)偏差(SD)、變異系數(shù)(CV)和相關(guān)系數(shù)(r)來評(píng)價(jià)候選內(nèi)參基因的表達(dá)穩(wěn)定性,SD和CV越小,r越大,候選內(nèi)參基因的穩(wěn)定性越好[24]。然后,根據(jù)Delta Ct方法計(jì)算各內(nèi)參基因表達(dá)量的平均標(biāo)準(zhǔn)偏差(STDEV)。基于geNorm、Normfinder、BestKeeper和Delta Ct四種分析結(jié)果,通過RefFinder在線分析工具(https://www.heartcure.com.au/reffinder/)計(jì)算其幾何平均數(shù)以獲得候選內(nèi)參基因穩(wěn)定性的綜合指數(shù)排名[25],綜合指數(shù)越小,說明基因的表達(dá)穩(wěn)定性越高。
1.2.4 內(nèi)參基因的驗(yàn)證 以篩選出來的內(nèi)參基因作為內(nèi)參,使用RT-qPCR對(duì)無患子三萜皂苷生物合成相關(guān)的6個(gè)編碼關(guān)鍵酶的基因(SmAACT、SmDXS、SmFPS、SmbAS1、SmCYP716A-5 和 SmUGT73C-14)和2個(gè)編碼轉(zhuǎn)錄因子的基因(SmbHLH8和Sm-ERF25)的相對(duì)表達(dá)量進(jìn)行測(cè)定,以驗(yàn)證篩選出來的內(nèi)參基因的可靠性。采用Ct(2-ΔΔCt)法計(jì)算相對(duì)表達(dá)量,再計(jì)算各樣本相對(duì)于果皮S1時(shí)期的表達(dá)量。各目的基因的引物見表2。使用Microsoft Excel 2016軟件對(duì)數(shù)據(jù)進(jìn)行處理,使用Origin 2017 SR2繪制圖表。

表2 目的基因的引物Table 2 Primers of objective genes
2 結(jié)果
2.1 RT-qPCR分析結(jié)果
RT-qPCR結(jié)果顯示8個(gè)候選內(nèi)參基因的擴(kuò)增效率在93.727%-100.206%之間,相關(guān)系數(shù)在0.997-0.999之間(表3),符合熒光定量PCR實(shí)驗(yàn)要求。如圖2所示,在無患子根、莖、葉、芽、花和果皮等器官中,8個(gè)內(nèi)參基因的平均Ct值在21.37(SmEF-1α)-28.04(SmTBCC) 之 間,SmEF-1α的變幅較大。
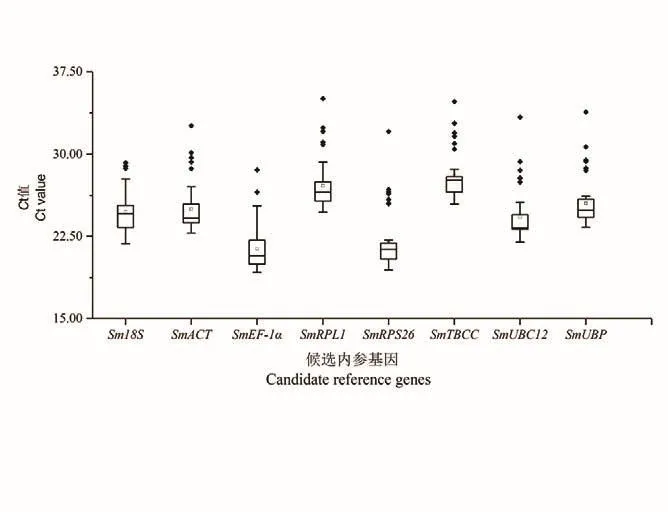
圖2 候選內(nèi)參基因的Ct值Fig.2 Ct values of candidate reference genes
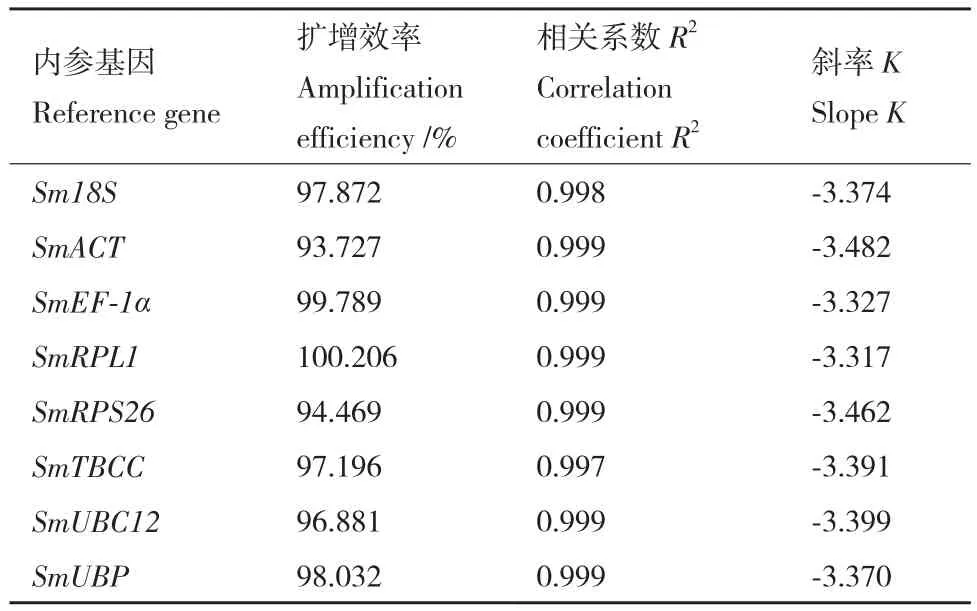
表3 候選內(nèi)參基因擴(kuò)增效率和標(biāo)準(zhǔn)曲線參數(shù)Table 3 Amplification efficiency and standard curve parameters of candidate reference genes
2.2 候選內(nèi)參基因的穩(wěn)定性分析
geNorm軟件分析結(jié)果表明,在無患子不同器官和不同發(fā)育時(shí)期果皮中,8個(gè)候選內(nèi)參基因的平均表達(dá)穩(wěn)定值(M)均小于1.5(表4)。其中,SmACT和SmRPL1的M值最小,為0.373;其次為SmUBC12,M值為0.508。候選內(nèi)參基因標(biāo)準(zhǔn)化因子的配對(duì)差異分析結(jié)果顯示V2/V3(0.182)大于0.15,V3/V4(0.126)小于 0.15( 圖3)。NormFinder軟件結(jié)果顯示8個(gè)候選內(nèi)參基因的穩(wěn)定性排序?yàn)镾mACT>SmUBP>SmRPL1>SmUBC12>SmTBCC>SmRPS26> Sm18S> SmEF-1α(表4)。BestKeeper軟件分析結(jié)果表明,SD值和CV值最小的基因分別為 Sm18S和 SmTBCC,SmACT、SmRPL1和 SmUBP的 r值最大(r=0.985)(表4)。采用 Delta Ct法對(duì)候選內(nèi)參基因的表達(dá)穩(wěn)定性進(jìn)行分析,結(jié)果表明SmACT(0.67) 的 STDEV 最 小,SmEF-1α(1.07)的STDEV最大(表4)。RefFinder綜合分析結(jié)果顯示,8個(gè)候選內(nèi)參基因在無患子不同器官和不同發(fā)育時(shí)期果皮中的表達(dá)穩(wěn)定性大小順序?yàn)镾mACT、SmRPL1、SmUBP、SmUBC12、Sm18S、SmTBCC、SmRPS26、SmEF-1α,其中SmACT是最理想的內(nèi)參基因,其次為SmRPL1和SmUBP(表4)。
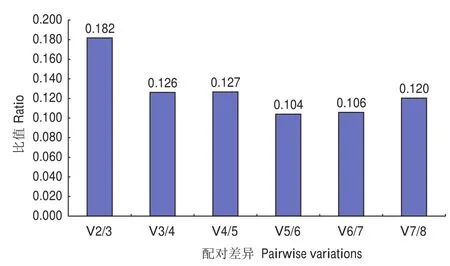
圖3 geNorm軟件分析內(nèi)參基因最適數(shù)目Fig.3 Analysis of optimal number of reference genes for normalization by geNorm

表4 GeNorm,NormFinder和BestKeeper分析結(jié)果和排名Table 4 GeNorm,NormFinder and BestKeeper analysis results and rankings
2.3 內(nèi)參基因穩(wěn)定性的驗(yàn)證
分別以篩選出來的3個(gè)穩(wěn)定性較好的內(nèi)參基因(SmACT、SmRPL1和SmUBP)和一個(gè)穩(wěn)定性較差的內(nèi)參基因(SmEF-1α)作為內(nèi)參,對(duì)8個(gè)皂苷合成相關(guān)基因在不同器官和不同發(fā)育時(shí)期果皮中的表達(dá)情況進(jìn)行研究。結(jié)果如圖4所示,SmAACT4在果皮S6-S8時(shí)期和雄花中有較高相對(duì)表達(dá)量;SmDXS4在果皮S5-S7時(shí)期和根中高表達(dá);SmFPS和SmCYP716A-5在果皮S1-S4時(shí)期和雌花中有較高相對(duì)表達(dá)量;SmbAS1和SmUGT73C-14的相對(duì)表達(dá)量隨著果皮生長先升高后下降,且均在S3時(shí)期達(dá)到最高;SmbHLH8和SmERF25在果皮S1時(shí)期和根、莖、葉、芽和雌花等器官中高表達(dá)。以SmACT、SmACT+SmRPL1組合和SmACT+SmRPL1+SmUBP組合進(jìn)行數(shù)據(jù)標(biāo)準(zhǔn)化,目的基因的相對(duì)表達(dá)量基本上保持一致;同時(shí),8個(gè)目的基因在不同器官和不同發(fā)育時(shí)期果皮中的表達(dá)規(guī)律與轉(zhuǎn)錄組測(cè)序結(jié)果相一致,進(jìn)一步驗(yàn)證了轉(zhuǎn)錄組測(cè)序結(jié)果的可靠性;以穩(wěn)定性較差的候選內(nèi)參基因SmEF-1α為內(nèi)參,會(huì)高估目的基因的相對(duì)表達(dá)水平。
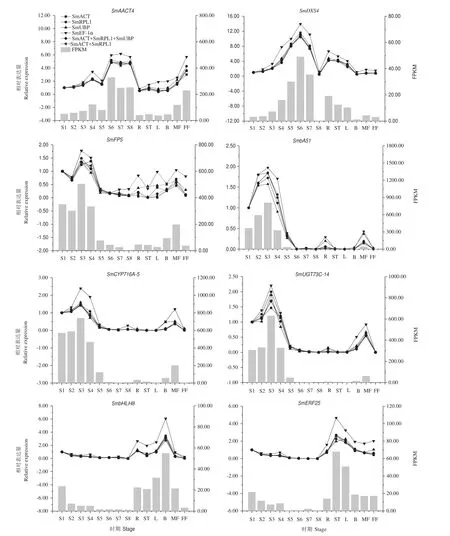
圖4 無患子三萜皂苷生物合成途徑中相關(guān)基因的RT-qPCR結(jié)果Fig.4 RT-qPCR expression pattern of genes related to the triterpenoid saponin biosynthesis of S.mukorossi
本研究對(duì)無患子的8個(gè)候選內(nèi)參基因在不同器官和不同果皮發(fā)育時(shí)期的表達(dá)量進(jìn)行RT-qPCR分析,結(jié)果顯示各基因的Ct值均有所變化,結(jié)合3種軟件對(duì)其穩(wěn)定性進(jìn)行評(píng)價(jià),結(jié)果表明不同軟件得出的穩(wěn)定性排序存在一定差異,geNorm、NormFinder軟件和Delta Ct法結(jié)果均顯示SmACT的穩(wěn)定性最好;而BestKeeper軟件分析表明SD值最小的基因?yàn)镾m18S,CV值最小的基因?yàn)镾mTBCC,r值最大的基因?yàn)镾mACT、SmRPL1和SmUBP。可見,GeNorm和NormFinder軟件的分析結(jié)果較為接近,而與BestKeeper軟件分析結(jié)果略有不同,這與蔣婷婷等[26]、Dudziak 等[27]、曹映輝等[28]、章穎佳等[29]的研究結(jié)果相似,造成這種差異的原因可能是各軟件所使用的算法不同[30]。ACT是一類球狀微絲結(jié)構(gòu)蛋白,在細(xì)胞中占總蛋白質(zhì)含量的5%以上,氨基酸序列高度保守[31]。ACT在植物組織中表達(dá)量大且基本穩(wěn)定,常被用于植物三萜皂苷生物合成途徑中相關(guān)基因的表達(dá)分析研究,如刺五加(Eleutherococcus senticosus)[32]、刺楸(Kalopanax septemlobus)[33]和細(xì)風(fēng)輪菜(Clinopodium gracile)[34]等。本研究結(jié)果也表明SmACT在無患子根、莖、葉、芽、花和果皮中表達(dá)最穩(wěn)定,是最適合作為三萜皂苷生物合成相關(guān)基因表達(dá)分析的內(nèi)參基因。另外,根據(jù)geNorm軟件得到的V2/V3>0.15、V3/V4<0.15的結(jié)果,說明3
3 討論
個(gè)穩(wěn)定基因的組合能增加定量結(jié)果的可靠性,所以SmACT、SmRPL1、SmUBP是理想的內(nèi)參基因組合。
本研究結(jié)果顯示以SmACT、SmACT+SmRPL1組合和SmACT+SmRPL1+SmUBP組合進(jìn)行數(shù)據(jù)標(biāo)準(zhǔn)化,無患子皂苷合成途徑中的6個(gè)關(guān)鍵酶編碼基 因(SmAACT4、SmDXS4、SmFPS、SmbAS1、SmCYP716A-5和SmUGT73C-14)和2個(gè)轉(zhuǎn)錄因子編碼基因(SmbHLH8和SmERF25)的相對(duì)表達(dá)量基本上保持一致,說明了這些內(nèi)參基因及內(nèi)參基因組合的可靠性。Xiao等[35]在羊躑躅(Rhododendron molle)不同發(fā)育階段和不同組織下內(nèi)參基因篩選的研究中也得到了類似的結(jié)論,即采用單個(gè)或兩個(gè)內(nèi)參的組合對(duì)目的基因進(jìn)行表達(dá)驗(yàn)證時(shí),其表達(dá)趨勢(shì)一致。所以,在無患子果皮RT-qPCR實(shí) 驗(yàn) 中, 使 用SmACT、SmACT+SmRPL1組 合、SmACT+SmRPL1+SmUBP組合作為內(nèi)參基因?qū)δ康幕蜻M(jìn)行校準(zhǔn)均可得到準(zhǔn)確和可靠的定量結(jié)果。
乙酰輔酶A酰基轉(zhuǎn)移酶(acetyl-CoA C-acetyltransferase,AACT)、1-脫氧D-木酮糖-5-磷酸合成酶(1-deoxy-D-xylulose-5-phosphat synthetase,DXS)、法呢基焦磷酸合酶(farnesyl pyrophosphate synthetase,F(xiàn)PS)、β-香樹素合成酶(β-amyrin synthase,β-AS)、細(xì)胞色素P450依賴性單加氧酶(Cytochrome P450-dependent monooxygenases,CYP450s)和 糖基轉(zhuǎn)移酶(uridine diphosphate(UDP)-glucosyltransferases,UGTs)是三萜皂苷生物合成途徑中的關(guān)鍵酶類,編碼這些酶的基因在無患子三萜皂苷生物合成過程中均起著重要作用[36-38]。課題組前期研究表明無患子果皮中的三萜皂苷含量最高(13.26%),其次為花(5.68%-6.21%)和葉(3.30%-5.35%)[13];此外,在果皮發(fā)育過程中,總皂苷含量總體上呈先增加后降低、然后保持在較高水平的趨勢(shì),在S3和S4時(shí)期有較高的總?cè)圃碥蘸浚?9]。本研究 發(fā) 現(xiàn) SmAACT4、SmDXS4、SmFPS、SmbAS1、SmCYP716A-5、SmUGT73C-14、SmbHLH8 和SmERF25等編碼基因在根、莖、葉、花等部分器官或果皮的部分發(fā)育時(shí)期中高表達(dá),說明其在無患子三萜皂苷生物合成中可能發(fā)揮著重要作用。另外,SmFPS、SmbAS1、SmCYP716A-5、SmUGT73C-14 的相對(duì)表達(dá)量在果皮發(fā)育前期(S1-S4時(shí)期)較高,SmbHLH8和SmERF25在果皮S1時(shí)期也有較高的相對(duì)表達(dá)量,說明這些關(guān)鍵基因或轉(zhuǎn)錄因子在調(diào)控?zé)o患子皂苷合成與積累的過程中可能存在著協(xié)同作用,具體機(jī)制有待進(jìn)一步研究。研究表明親緣關(guān)系相近的物種中同一類同源基因的保守性一般會(huì)更高,所以本研究篩選出的內(nèi)參基因SmACT、SmRPL1和SmUBP不僅可用作無患子三萜皂苷合成相關(guān)基因表達(dá)量分析的內(nèi)參基因,也可以應(yīng)用于無患子及近緣植物的其他生物學(xué)過程中的基因表達(dá)研究。然而,本研究僅對(duì)無患子根、莖、葉、芽、花和不同發(fā)育時(shí)期果皮進(jìn)行了研究,篩選出的內(nèi)參基因在各類脅迫處理的無患子植株中的穩(wěn)定性有待進(jìn)一步驗(yàn)證。
4 結(jié)論
從無患子8個(gè)候選內(nèi)參基因中,篩選得到SmACT、SmRPL1和SmUBP等3個(gè)表達(dá)穩(wěn)定性較好的內(nèi)參基因,以這些基因?yàn)閮?nèi)參對(duì)三萜皂苷生物合成途徑中的8個(gè)候選基因進(jìn)行校準(zhǔn)所得的表達(dá)量基本上保持一致,且相對(duì)表達(dá)量結(jié)果與轉(zhuǎn)錄組結(jié)果基本保持一致,表明SmACT、SmACT+SmRPL組合和SmACT+SmRPL+SmUBP組合可作為無患子三萜皂苷生物合成途徑中相關(guān)基因表達(dá)研究的內(nèi)參基因。
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年18期)2018-11-14 01:48:24
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
