煙酸促進短乳桿菌合成γ-氨基丁酸的機制
2023-01-05 08:45:44張健才恒申振豪劉仲浩趙娟張巧珍高強
生物技術通報 2022年10期
關鍵詞:分析
張健 才恒 申振豪 劉仲浩 趙娟 張巧珍 高強
(1.天津科技大學生物工程學院,天津 300457;2.天津市工業微生物重點實驗室,天津 300457;3.工業發酵微生物教育部重點實驗室,天津 300457;4.天津科技大學現代分析技術研究中心,天津 300457;5.天津科技大學理學院,天津 300457)
γ-氨基丁酸(GABA)作為重要的抑制神經訊息傳遞的物質[1],能夠參與人體內多種代謝過程,具有較高的生理活性[2-3]。短乳桿菌在發酵過程中產生乳酸,導致生長環境的pH值降低,從而抑制其生長[4]。為了抵抗酸脅迫,短乳桿菌形成了多種抗酸機制,常見的抗酸機制有谷氨酸脫羧酶(GAD)系統[5]、F0-F1-ATPase質子泵、精氨酸脫亞胺酶途徑、生物胺產生系統[6]。GAD系統中,胞外的谷氨酸由轉運蛋白GadC轉運至胞內,并通過磷酸吡哆醛依賴的谷氨酸脫羧酶GadA或GadB脫羧生成GABA和二氧化碳,生成的GABA可由GadC轉運到胞外。該過程消耗胞內H+,外排偏中性的GABA,可有效提高環境的pH值,幫助微生物抵御酸脅迫[7-8]。Lv等[9]通過篩選低F0-F1-ATPase活性的短乳桿菌,來提高GABA的產量,依據是使H+更多的通過GAD系統消耗。受此啟發,我們設想抑制短乳桿菌生物胺產生系統,使H+更多的通過GAD系統消耗而提高GABA的產量。酪胺是生物胺的一種,短乳桿菌可以通過酪氨酸脫羧酶將酪氨酸脫羧生成堿性的酪胺來提高環境pH[10-11]。煙酸是酪氨酸脫羧酶的抑制劑[12],故本實驗計劃添加煙酸來抑制短乳桿菌發酵過程中酪胺的產生,弱化酪胺產生系統,使短乳桿菌更多地通過GAD系統來抵御酸脅迫,從而提高其GABA產量。該思路已經被前期實驗驗證,即在發酵中添加0.15%的煙酸確實能顯著提高GABA的產量。為了闡明煙酸促進短乳桿菌合成GABA的作用機制,本研究采用液相色譜-質譜聯用(LC-MS)的方法探究短乳桿菌添加和不添加煙酸發酵時的差異代謝物[13],分析煙酸對短乳桿菌代謝通路的影響,并采用碘化丙啶(PI)單染色法[14]分析了細胞膜的完整性,旨為發酵法生產GABA提供新的思路。
1 材料與方法
1.1 材料
1.1.1 菌種 短乳桿菌(Lactobacillus brevis)CGMCC 3414[15],本實驗室分離保藏于中國微生物菌種保藏中心。
1.1.2 儀器與試劑 Agilent1200高效液相色譜儀(Angilent科技有限公司);色譜柱:ZORBAX SBC184.6 mm×250 mm,5 μm;高速臺式離心機(上海安亭科學儀器廠);BX53正置熒光顯微鏡(日本Olympus);320-S型 pH 計(Mettler Toledo);LDZX-75KBS立式壓力蒸汽滅菌鍋(上海申安醫療器械廠);電子分析天平(Mettler Toledo);生化培養箱(廣東省醫療器械廠);SBA-40C生物傳感分析儀(山東省科學院生物研究所);AB Triple TOF 6600質譜儀(AB SCIEX);Agilent1290InfinityLC超高壓液相色譜儀(Agilent);色譜柱:Waters,ACQUITY UPLC BEH Amide 1.7 μm,2.1 mm×100 mm。
乙腈(色譜純,天津市科密歐化學試劑有限公司);乙酸銨(Sigma);谷氨酸鈉(99%,紅玫瑰味精);2,4-二硝基氟苯(DNFB,分析純,Sigma公司);乙酸鈉(分析純,生工生物工程股份有限公司);酵母粉(分析純,北京奧博生物技術有限公司);硫酸鎂(分析純,天津市北方天醫化學試劑廠);硫酸錳(分析純,天津市北方天醫化學試劑廠);葡萄糖(分析純,天津市化學試劑一廠);吐溫-80(分析純,天津市江天化工技術有限公司);煙酸(分析純,天津市北方天醫化學試劑廠)。衍生緩沖液:取碳酸鈉4.2 g用超純水定容至100 mL;衍生劑:0.1 mL DNFB用乙腈定容至50 mL;定容緩沖液:取磷酸氫二鉀3.4 g溶于145.5 mL的0.1 mol/L的氫氧化鈉溶液,超純水定容至1 000 mL。
1.1.3 培養基 種子培養基:蛋白胨10 g/L,酵母粉5 g/L,葡萄糖10 g/L,乙酸鈉2 g/L,硫酸錳0.05 g/L,硫酸鎂0.2 g/L,吐溫80 1 mL/L。
發酵培養基:葡萄糖30 g/L,酵母粉35 g/L,谷氨酸鈉(MSG)50 g/L,硫酸亞鐵0.05 g/L,硫酸鎂0.2 g/L,硫酸銨2 g/L,pH 5.5。
1.2 方法
1.2.1 發酵過程中菌體生長、葡萄糖消耗和GABA合成 從甘油管中取100 μL菌液接于50 mL種子培養基中,在30℃、搖床轉速50 r/min培養24 h 后,以5%(V/V)的接種量轉接于下一級種子培養基中,30℃、靜置培養12 h。將種子菌液以 5%(V/V)的接種量轉接于50 mL發酵培養基中,發酵培養基中添加0.15 %煙酸的為實驗組,不含煙酸的為對照組,在30℃靜置培養72 h。在發酵過程中每6 h取1 mL菌液,稀釋10倍后用分光光度計在600 nm下測定吸光度,計為OD600,反映菌體生長情況;然后將稀釋10倍后的菌液8 000 r/min離心10 min取上清液,使用生物傳感分析儀測量葡萄糖含量。另外,在發酵24 h、36 h、48 h、60 h和72 h時,各取1 mL發酵液,8 000 r/min離心10 min取上清,蒸餾水稀釋10倍后取10 μL于1.5 mL離心管中,依次加入衍生緩沖液100 μL、衍生劑100 μL、定容緩沖液790 μL,振蕩混勻,60℃水浴1 h。反應后用0.22 μm醋酸纖維膜過濾,用高效液相色譜儀測定GABA的產量[16]。
1.2.2 代謝組學樣品的制備 取對數后期發酵液對照組和實驗組各6組平行,通過分光光度計調到相同的OD600值,定容后在4℃下8 000 r/min離心10 min取菌體沉淀,快速用生理鹽水在4℃下8 000 r/min離心10 min的條件下洗滌菌體兩次得到菌體樣本,加入預冷甲醇/乙腈/水溶液(2∶2∶1,V/V/V),渦旋混合,低溫超聲30 min,-20℃靜置10 min,4℃下12 000 r/min離心20 min,取上清真空干燥,質譜分析時加入100 μL乙腈水溶液(乙腈∶水=1∶1,V/V)復溶,渦旋,4℃下12 000 r/min 離心15 min,取上清液進樣分析[17]。
1.2.3 LC-MS中液相色譜條件 樣品采用超高效液相色譜系統(ultra performance liquid chromatography,UPLC)HILIC色譜柱進行分離;柱溫25℃;流速0.5 mL/min;進樣量2 μL;流動相組成A:水+25 mmol/L乙酸銨+25 mmol/L氨水,B:乙腈;梯度洗脫程序如下:0-0.5 min,95% B;0.5-7 min,B從95%線性變化至65%;7-8 min,B從65%線性變化至40%;8-9 min,B維持在40%;9-9.1 min,B從40%線性變化至95%;9.1-12 min,B維持在95%;整個分析過程中樣品置于4℃自動進樣器中。為避免儀器檢測信號波動而造成的影響,采用隨機順序進行樣本的連續分析。樣本隊列中插入QC樣品,用于監測和評價系統的穩定性及實驗數據的可靠性[18]。
1.2.4 質譜條件 分別采用電噴霧電離正離子和負離子模式進行檢測。ESI源條件如下:離子源溫度600℃,噴霧電壓(ion sapary voltage floating,ISVF)±5 500 V(正負兩種模式);TOF MS掃描范圍(m/z):60-1 000 Da,產物離子掃描范圍(m/z):25-1 000 Da,TOF MS 掃描累積時間 0.20 s/spectra,產物離子掃描累積時間0.05 s/spectra;二級質譜采用information dependent acquisition(IDA)獲得,并且采用高靈敏模式,Declustering potential(DP):±60 V(正負兩種模式),碰撞能量Collision Energy:(35±15)eV。
1.2.5 數據分析 質譜結果通過標準品數據庫(inhouse database(Shanghai Applied Protein Technology))中代謝物的保留時間、分子質量等信息進行匹配,對樣本中的代謝物進行結構鑒定,并對鑒定結果進行嚴格核對。對實驗檢測到的代謝物結果進行預處理后,使用SIMCA軟件(V15.0.2)進行數據分析。數據分析內容包括正交偏最小二乘判別分析(OPLSDA)等分析,在最大程度保留原始信息的基礎上對采集的多維數據進行降維分析,然后進行差異代謝物篩選、差異代謝物相關性分析、KEGG 通路分析等內容[19]。
1.2.6 碘化丙啶單染色 細菌的細胞膜不完整時,碘化丙啶(propidium iodide,PI)能夠通過細胞膜進入細胞與DNA結合,并且發出更亮的紅色熒光,PI不能進入擁有完整細胞膜的胞內,用來檢測細胞膜的完整性[20]。將發酵對數生長前期的實驗組和對照組菌液的OD600調至0.8,PBS溶液清洗菌體3次并用PBS溶液懸浮菌體,加入PI使其終濃度為5 μg/mL,輕輕搖勻菌液在黑暗下靜置15 min,最后吸取合適的菌液滴于載玻片上通過正置熒光顯微鏡觀察來判定細胞膜的通透性和完整性。
2 結果
2.1 發酵過程中菌體生長、葡萄糖消耗和GABA合成
短乳桿菌發酵過程中的GABA產量如圖1-A所示,培養48 h后,實驗組(experimental group,EG)的GABA產量開始顯著高于對照組(control group,CG),至72 h時實驗組比對照組的GABA產量提高44%。發酵過程中菌體OD值(OD600)和葡萄糖含量的變化如圖1-B所示,添加0.15%煙酸的實驗組中菌體生長較之對照組明顯減緩,相應的,發酵前24 h實驗組中的葡萄糖消耗也更慢,但發酵30-42 h,實驗組的葡萄糖消耗比對照組更多,此后實驗組和對照組的葡萄糖消耗趨于一致。
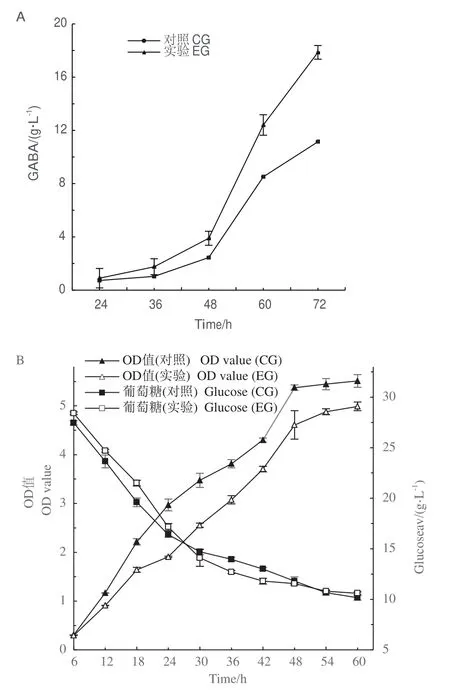
圖1 實驗組(煙酸)和對照組在不同發酵時間下的GABA產量、OD值和葡萄糖的變化Fig.1 Changes in GABA yield,OD and glucose in experimental groups(nicotinic acid)and control groups at different fermentation times
2.2 多變量統計分析
根據 OPLS-DA方法對數據進行分析,OPLS-DA是一種多因變量對多自變量的回歸建模方法,最大特點是可以去除自變量和分類變量無關的異常值,根據得分圖可以揭示數據離散程度,發現異常值。圖2-A和圖2-B顯示的得分圖上對照組和實驗組的組間分別位于兩側,則兩組間的代謝輪廓具有顯著差異或者是差異代謝物物的含量具有顯著性變化。
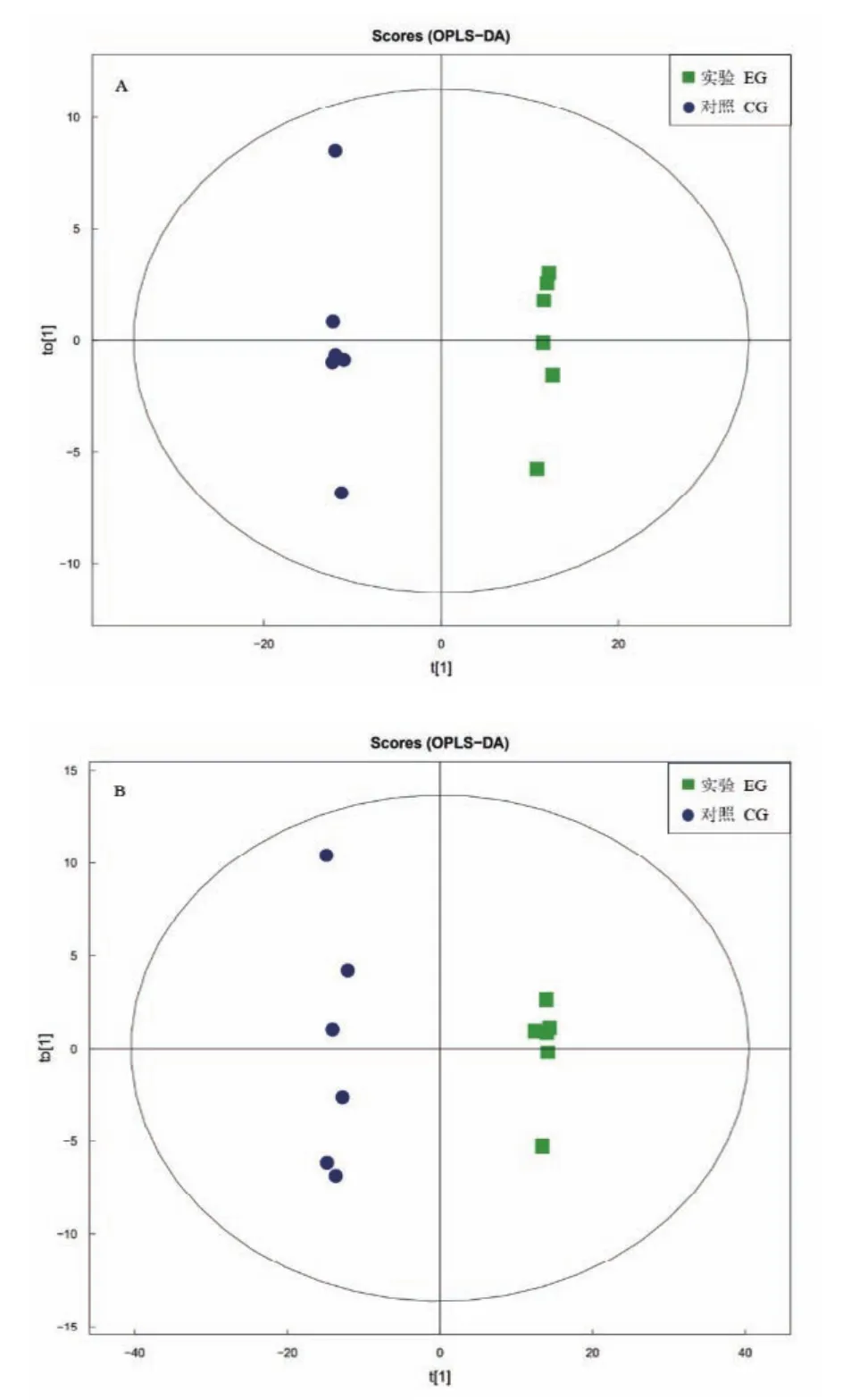
圖2 正(A)負(B)離子模式OPLS-DA得分圖Fig.2 OPLS-DA score of positive(A)and negative(B)ion mode
2.3 差異代謝物的篩選
正離子模式下鑒定的代謝物數量為204種,負離子模式下鑒定出120種代謝物,其中主要為氨基酸、脂質、有機酸及其衍生物,大量且豐富的代謝物數據有利于全面檢測出代謝通路的變化。基于OPLS-DA模式下VIP>1篩選兩組間差異代謝物,篩出兩組間具有顯著性的變量(P<0.05),通過HMDB和KEGG數據庫比對,篩選出33個顯著性差異代謝物如表1所示,其中調節趨勢是根據實驗組與對照組的差異代謝物含量的比值為依據(↑*:1-2;↑ **:2-5;↑ ***:>5;↓ *:0.6-0.9;↓ **:0.3-0.6;↓***:<0.3)。結果顯示實驗組中酪氨酸脫羧酶的產物酪胺含量顯著降低,有關脂質代謝的十八碳二烯酸、二十二碳一烯酸、二十二碳二烯酸、20-羥二十烷四烯酸和棕櫚油酰磷脂酰甘油等不飽和脂肪酸的含量均顯著高于對照組,而對應的飽和脂肪酸中單棕櫚酰甘油含量降低;氨基酸代謝中組脯氨酸、谷氨酸、組氨酸、天冬氨酸、絲氨酸和鳥氨酸的含量都是實驗組顯著高于對照組;實驗組中與能量代謝有關的AMP和2-磷酸甘油酸含量均顯著下降,證明了煙酸對菌體生長的抑制。
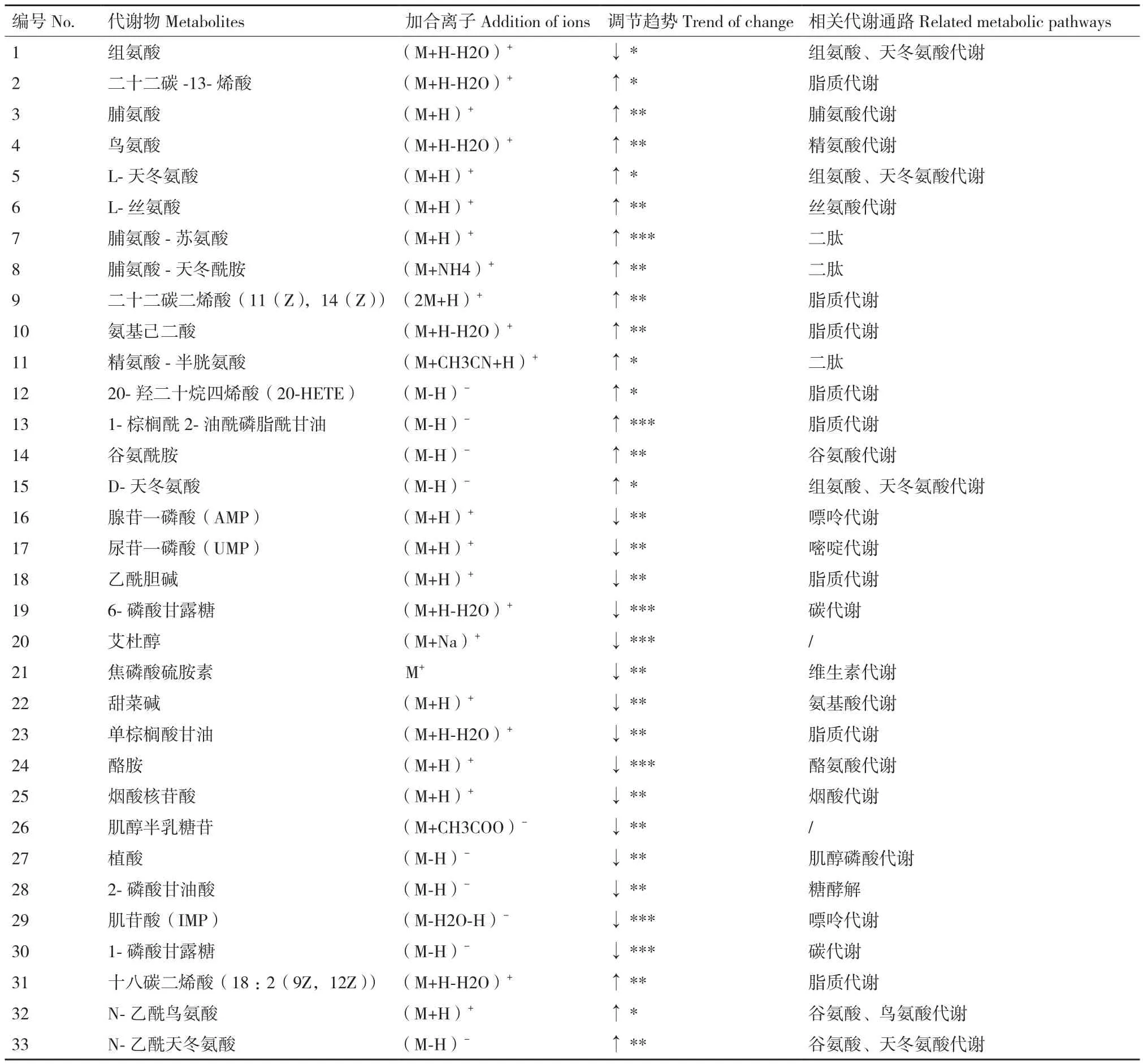
表1 正負離子模式下顯著性差異代謝物物Table 1 Significant difference metabolites in positive and negative ion mode
2.4 碘化丙啶(PI)單染色結果
實驗組和對照組對數生長前期的PI染色結果如圖3所示,實驗組中具有紅色熒光的短乳桿菌細胞明顯多于空白實驗組中的細胞,說明實驗組中短乳桿菌細胞膜的通透性顯著高于對照組細胞。

圖3 煙酸對短乳桿菌的細胞膜通透性和完整性影響Fig.3 Effect of nicotinic acid on cell membrane permeability and integrity of L.brevis
3 討論
3.1 脂質代謝
脂質不僅是能量代謝產物,還是細胞膜的重要組成成分。細胞在經受環境脅迫時會調節飽和脂肪酸和不飽和脂肪酸的比例,來影響細胞膜通透性,當不飽和脂肪酸含量增多時細胞膜的通透性會加大[21-22]。代謝組學結果表明,實驗組不飽和脂肪酸含量增加且飽和脂肪酸含量減少,暗示實驗組細胞膜通透性增加,后續的PI染色結果證實了這一點。細胞膜的通透性增加,使得底物谷氨酸更容易進入胞內進行脫羧反應,同時生成的GABA也更易排出細胞。
3.2 氨基酸代謝
由于乳酸菌缺乏必要的生物代謝途徑,不能合成生長所必需的氨基酸,乳酸菌必須從培養基中獲得這些生物活性物質才能滿足其快速生長需要。乳酸菌依靠胞外蛋白酶水解培養基中的蛋白質形成寡肽和氨基酸,這些水解產物通過轉運系統進入細胞,最后由細胞內的各種肽酶裂解形成游離氨基酸供細胞合成代謝[23]。脯氨酸、谷氨酸、組氨酸、絲氨酸均是短乳桿菌的必需氨基酸,鳥氨酸和天冬氨酸能夠通過谷氨酸代謝合成,屬于短乳桿菌的非必需氨基酸。實驗組中上述氨基酸含量均高于對照組,證明實驗組中肽轉運蛋白活性更強,或者胞內肽酶活性更強,或者細胞通透性增加導致胞內必需氨基酸含量增加。雖然胞內必需氨基酸增加,但是實驗組的菌體生長更慢,顯然是其它原因抑制了實驗組菌體的生長。另外,實驗組中酪胺含量的顯著下降,證明胞內酪胺的合成受到抑制,煙酸對酪氨酸脫羧酶的抑制是有效的。
3.3 碳代謝
短乳桿菌是典型的兼性厭氧菌且進行異型乳酸發酵。1-磷酸甘露糖和6-磷酸甘露糖是核苷酸糖的主要來源,而核苷酸糖主要應用于細胞結構物質的合成,本研究中1-磷酸甘露糖和6-磷酸甘露糖的含量在添加煙酸后顯著降低,說明在實驗組代謝過程中核苷酸糖類合成較少,不利于細胞生長繁殖。焦磷酸硫胺素是丙酮酸脫氫酶的輔因子,在實驗組中含量顯著降低,可能會使丙酮酸脫氫酶的活性變低,減少乙酰CoA生成從而不利于脂肪酸的合成而影響細胞膜的形成。
3.4 其他差異代謝物
氨基己二酸是不飽和脂肪酸的氧化產物,實驗組中其含量上升意味著細胞膜完整性遭到破壞。肌醇半乳糖苷是典型的抗逆物質與多種耐逆響應有關[24],甜菜堿是一種生物堿化學名稱為N,N,N-三甲基甘氨酸,艾杜醇作為一種糖醇類物質,其兩者作用與脯氨酸相似都能起到維持細胞滲透壓的作用,甜菜堿和艾杜醇含量降低的原因是在生長過程中微生物會優先從外界轉運能夠維持滲透壓的溶質[25];通過代謝通路分析發現煙酰胺只會單向合成煙酸,煙酸含量的增加可能會抑制煙酰胺產物的生成,從而抑制NAD+(煙酰胺腺嘌呤二核苷酸)的合成,導致煙酸核苷酸的含量降低,抑制多種脫氫酶的活性,進而抑制脂肪酸合成和菌體生長。植酸是一種非離子表面活性劑,當積累在細胞膜表面可以改善通透性[26],實驗組中植酸含量降低可能是因為細胞膜的完整性受到破壞。
短乳桿菌發酵中添加煙酸能顯著提高GABA的產量,但煙酸促進短乳桿菌GABA合成的機制未知。利用非靶標代謝組學可以無偏向性地分析短乳桿菌發酵后盡可能多的代謝產物,通過分析添加煙酸與不添加煙酸發酵后的差異代謝物,可能從全局角度理解煙酸促進短乳桿菌GABA合成機制,將會對GABA的生物合成提供理論基礎和新的思路。
4 結論
本研究利用LC-MS非靶向代謝組學方法分析了添加煙酸和不添加煙酸時,短乳桿菌細胞內的差異代謝物。得到的33種具有顯著性差異的代謝物能夠看出添加煙酸后,酪胺和核苷酸糖的含量減少的同時,不飽和脂肪酸含量增加。通過PI單染色實驗,發現添加煙酸后的短乳桿菌細胞膜的通透性顯著高于不添加煙酸的對照組。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
