乙酸鈉對油酸誘導的BRL-3A細胞脂肪變性的影響及機制分析
2023-01-03 02:28:42宮彬彬王廣力張源淑
畜牧獸醫學報 2022年12期
關鍵詞:模型
李 林,宮彬彬,王廣力,趙 梅,張源淑
(1.邢臺學院生物科學與工程學院,邢臺 054001;2.河北醫科大學附屬邢臺人民醫院病理科,邢臺 054031;3. 南京農業大學動物醫學院農業部動物生理生化重點開放實驗室,南京 210095)
非酒精性脂肪性肝病(non-alcoholic fatty liver disease,NAFLD)是一種以三酰甘油(triglyceride,TG)在肝蓄積為特征的疾病,近年來,由于脂肪和高糖等攝入增加、激素食品泛濫等因素,使NAFLD發病率逐年增高。NAFLD在雞、狗、豬、牛、羊以及魚等動物也可發生,是各種動物的原發或者繼發性疾病,在雞、鴨的發病率較高,該病不僅影響著養殖業的發展,也會帶來一定的食品安全問題,給禽類養殖業造成嚴重的經濟損失[1]。近年來,NAFLD越來越受到關注,因其發病機制復雜、診斷困難,已成為世界上最常見的慢性肝病。NAFLD通常與高血壓、血脂異常、肥胖、胰島素抵抗、代謝綜合征、2型糖尿病和心血管疾病密切相關[2]。目前,NAFLD沒有有效的治療方案,主要以改變動物的飲食方式為主。
乙酸是一種短鏈脂肪酸(SCFA),它是由攝入的膳食纖維刺激,經動物盲腸和結腸發酵產生[3-4]。據報道,膳食纖維可以降低餐后血糖反應、血漿膽固醇和三酰甘油濃度以及脂肪儲存[5]。并且,Hara等[6]研究發現含有SCFA的無纖維飲食也能降低血清膽固醇。乙酸除了由結腸細菌發酵產生外,許多食物中均含有乙酸的成分。
肝是動物調節脂質代謝和維持脂質穩態的重要器官,通過肝切片發現,肝含有多種細胞類型,如肝細胞、庫普弗細胞、內皮細胞和成纖維細胞[7]。研究表明,乙酸可以使小鼠肝TG含量降低,脂質氧化基因表達增加[8]。此外,乙酸可激活糖尿病KK-A(y)小鼠肝腺苷酸活化蛋白激酶(AMP-activated protein kinase, AMPK)蛋白,而AMPK作為能量的“傳感器”和“調節器”,對肝的脂質代謝的調節具有重要作用[9]。目前,乙酸在調控肝代謝方面的研究較少,且不深入。因此,本試驗通過油酸誘導BRL-3A細胞建立脂肪變性模型,分析乙酸鈉是否對BRL-3A細胞脂肪變性模型具有調控作用,為乙酸鈉對動物攝食調控及改善肥胖、健康方面的研究提供理論依據。
1 材料與方法
1.1 細胞來源
大鼠肝成纖維細胞(BRL-3A)購自中國科學院昆明細胞庫,本實驗室凍存。
1.2 主要試劑
乙酸鈉、油酸購自美國Sigma公司;TG試劑盒、油紅O染色試劑盒、丙氨酸氨基轉移酶、天門冬氨酸氨基轉移酶試劑盒均購自南京建成生物技術有限公司;P-AMPK-兔/AMPK-兔抗體均購自艾博抗上海貿易有限公司;山羊抗兔IgG購自上海生物工程有限公司;BCA蛋白定量試劑盒購自上海碧云天生物技術公司;DMEM培養基、胎牛血清和青霉素/鏈霉素購自南京澤優生物科技有限公司。
1.3 BRL-3A細胞脂肪變性模型建立
1.3.1 BRL-3A細胞的培養與處理 BRL-3A細胞在DMEM高糖培養基中進行培養,內含10%胎牛血清、100 U·mL-1青霉素/鏈霉素,置于含5% CO2的恒溫培養箱(37 ℃)中培養,根據細胞生長狀態,及時更換新鮮培養基,每隔2~3 d將細胞重新傳代1次。當細胞融合度達到80%~90%時,貼壁細胞用含0.25%胰酶和0.02% EDTA胰酶消化液進行消化處理,將細胞接種于6孔板和96孔板中,加入新鮮培養基,置于恒溫培養箱中培養。
1.3.2 不同濃度油酸處理BRL-3A細胞相對活力檢測 接種于96孔板內的BRL-3A細胞融合度達到70%~80%時,棄去培養液。PBS洗滌后,分別加入含有不同濃度油酸(0.03、0.06、0.12、0.24、0.48 mmol·L-1)的無血清培養基;在5% CO2的恒溫培養箱(37 ℃)中培養24 h。隨后每孔細胞加入含0.5 mg·mL-1MTT的無胎牛血清培養液20 μL,繼續培養4 h 后,棄上清液,再加入150 μL DMSO 充分溶解,用酶標儀檢測各孔的吸光值,檢測其細胞相對活力。
1.3.3 不同濃度油酸處理BRL-3A細胞總脂滴面積、TG、AST和ALT指標檢測 將細胞按照合適密度接種于6孔板中,待細胞融合至80%時,棄去培養液,對細胞進行饑餓處理后,加入含有不同濃度油酸(0.03、0.06、0.12、0.24 mmol·L-1)的無血清培養基,在37 ℃、5% CO2條件下培養24 h后,PBS沖洗3遍,10%福爾馬林將細胞固定30 min,室溫條件下,PBS沖洗3遍,用油紅O染液染色10 min,PBS緩沖液洗2次;倒置顯微鏡觀察細胞內脂滴染色情況,并進行統計分析。另外,在37 ℃、5% CO2條件下培養24 h后收集細胞,棄上清進行TG含量、AST和ALT活性檢測。
1.4 不同濃度乙酸鈉處理BRL-3A細胞凋亡率指標檢測
將細胞培養于6孔板中,當細胞融合度達到80%~90%時,用低(2 mmol·L-1)、中(4 mmol·L-1)、高(8 mmol·L-1)濃度的乙酸鈉處理細胞24 h。收集細胞后加入200 μL的Annexin V-異硫氰酸熒光素(FITC)原液重懸細胞,4 ℃避光孵育30 min,再加入10 μL碘化丙啶(PI),避光孵育15 min,采用美國BD FACS Calibur型流式細胞術檢測熒光強度,計算細胞凋亡率。
1.5 乙酸鈉對油酸誘導的BRL-3A細胞脂肪變性模型的影響
1.5.1 不同濃度乙酸鈉處理脂肪變性細胞模型胞內脂滴變化檢測 將細胞按照合適密度接種于6孔板中,待細胞融合至80%時,棄去培養液,對細胞進行饑餓處理后,將細胞分為油酸處理組(加入含有終濃度為0.12 mmol·L-1油酸的無血清培養基),2 mmol·L-1乙酸鈉+油酸處理組,4 mmol·L-1乙酸鈉+油酸處理組和8 mmol·L-1乙酸鈉+油酸處理組。在2 mmol·L-1乙酸鈉+油酸處理組、4 mmol·L-1乙酸鈉+油酸處理組和8 mmol·L-1乙酸鈉+油酸處理組中,均為2 h后加入乙酸鈉。在37 ℃、5% CO2條件下培養24 h后, PBS沖洗3遍,10%福爾馬林將細胞固定30 min,室溫條件下,PBS沖洗3遍,用油紅O染液染色10 min,PBS緩沖液洗2次;倒置顯微鏡觀察細胞內脂滴染色情況,并進行統計分析。
1.5.2 不同濃度乙酸鈉處理脂肪變性細胞模型TG、AST和ALT指標檢測 接種于6孔板的細胞經過上述同樣處理后,在37 ℃、5% CO2條件下培養24 h后,收集細胞,棄上清,進行TG含量、AST和ALT活性檢測,操作步驟按照說明書進行。提取細胞總蛋白,BCA蛋白濃度測定試劑盒測定并計算出蛋白濃度。
1.5.3 不同濃度乙酸鈉處理脂肪變性細胞模型AMPK通路蛋白檢測 Western blot檢測各處理細胞中P-AMPK/AMPK蛋白的表達變化,參照李林等[10]的方法進行操作。用ECL發光液進行Western blot圖像處理。用Image Lab 6.0分析條帶灰度值。
1.5.4 不同濃度乙酸鈉處理脂肪變性細胞模型脂代謝相關關鍵基因檢測 將收集的細胞采用TRIzol法直接提取總RNA,利用BioPhotometer測定樣品總RNA濃度,分析OD260 nm/OD280 nm值,判斷提取總RNA的純度,OD260 nm/OD280 nm需要為1.8~2.0。取1 μg總RNA進行反轉錄得到cDNA,詳細操作步驟參照說明書進行。
參照GenBank序列,用Primer Premier 5軟件自行設計,設計完成后送上海Sangon公司合成,引物序列見表1。

表1 目的基因及β-Actin引物序列
1.6 數據統計與分析
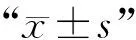
2 結 果
2.1 不同濃度油酸對BRL-3A細胞相對活力的影響
圖1所示,向細胞中添加不同濃度(0.03、0.06、0.12、0.24、0.48 mmol·L-1)的油酸后,發現與對照組(0 mmol·L-1油酸)相比,0.48 mmol·L-1油酸對細胞有明顯的抑制作用,且差異顯著(P<0.05)。其他濃度的油酸處理細胞后與對照組相比,均無差異。結果提示:0.03~0.24 mmol·L-1的油酸對BRL-3A細胞無毒副作用。
2.2 不同濃度油酸對BRL-3A細胞總脂滴的影響
圖2所示,與對照組相比,0.12和0.24 mmol·L-1油酸處理后細胞總脂滴面積均極顯著上升(P<0.01)。
2.3 不同濃度油酸對BRL-3A細胞三酰甘油(TG)、天門冬氨酸氨基轉移酶(AST)及丙氨酸氨基轉移酶(ALT)的影響
表2所示,向細胞中添加不同濃度(0.03、0.06、0.12、0.24 mmol·L-1)的油酸。發現與對照組相比,0.06 mmol·L-1油酸處理后,TG含量顯著上升(P<0.05);并且在0.12和0.24 mmol·L-1油酸處理后,TG含量極顯著上升(P<0.01),提示造成了BRL-3A細胞的脂質蓄積。與對照組相比,0.12和0.24 mmol·L-1油酸處理后,BRL-3A細胞AST和ALT活性顯著上升(P<0.05);其他濃度的油酸處理細胞后,AST和ALT活性均無顯著差異。
綜合以上細胞活力、細胞總脂滴面積、TG含量和AST、ALT活性測定結果,本試驗用不同濃度油酸誘導建立BRL-3A細胞體外脂肪變性模型的條件: 0.12 mmol·L-1油酸,培養時間為24 h。因此,后續選取0.12 mmol·L-1油酸進行誘導脂肪變性細胞模型處理試驗。

與對照組(0 mmol·L-1)相比,*. P<0.05Compared with control group(0 mmol·L-1),*. P<0.05圖1 油酸對BRL-3A細胞相對活力分析Fig.1 Analysis of the relative activity of oleic acid on BRL-3A cells

與對照組相比,**表示差異極顯著(P<0.01)Compared with control group,**.P<0.01圖2 油酸處理BRL-3A細胞總脂滴面積結果分析Fig.2 Analysis of total lipid droplet area of BRL-3A cells treated with oleic acid

表2 油酸對BRL-3A細胞三酰甘油(TG)、天門冬氨酸氨基轉移酶(AST)及丙氨酸氨基轉移酶(ALT)的影響分析
2.4 不同濃度乙酸鈉對脂肪變性細胞模型總凋亡率的影響
分別用低(2 mmol·L-1)、中(4 mmol·L-1)、高(8 mmol·L-1)濃度的乙酸鈉作用BRL-3A細胞24 h后,通過流式細胞術檢測細胞凋亡率,發現不同濃度的乙酸鈉對細胞凋亡率均無影響(圖3),證明乙酸鈉對細胞無毒害作用,可以進行下一步試驗。

圖3 乙酸鈉對BRL-3A細胞總凋亡率分析Fig.3 Analysis of sodium acetate on total apoptosis rate of BRL-3A cells
2.5 不同濃度乙酸鈉對脂肪變性細胞模型脂滴的影響
圖4所示,用0.12 mmol·L-1油酸分別和2、4、8 mmol·L-1的乙酸鈉共同孵育細胞24 h后,與油酸處理組相比,4 mmol·L-1乙酸鈉+油酸處理組和8 mmol·L-1+油酸處理組細胞總脂滴面積和每平方毫米脂滴數極顯著下降(P<0.01)。提示,4和8 mmol·L-1的乙酸鈉可以顯著降低脂肪變性細胞模型的脂質堆積。

與油酸處理組相比,**. P<0.01Compared with oleic acid treatment group,**.P<0.01圖4 乙酸鈉對脂肪變性細胞模型脂滴影響分析Fig.4 Analysis of sodium acetate on lipid droplets in steatosis cell models
2.6 不同濃度乙酸鈉對脂肪變性細胞模型三酰甘油(TG)、天門冬氨酸氨基轉移酶(AST)及丙氨酸氨基轉移酶(ALT)的影響
表3所示,向細胞中添加不同濃度的乙酸鈉后,發現與油酸處理組相比,4 mmol·L-1乙酸鈉+油酸處理后,TG含量、AST活性和ALT活性均顯著下降(P<0.05);并且在8 mmol·L-1乙酸鈉+油酸處理后,與對照組相比,TG含量極顯著下降(P<0.01),ALT、AST活性均顯著下降(P<0.05)。

表3 乙酸鈉對脂肪變性細胞模型三酰甘油(TG)、天門冬氨酸氨基轉移酶(AST)及丙氨酸氨基轉移酶(ALT)的影響分析
2.7 不同濃度乙酸鈉對脂肪變性細胞模型AMPK信號蛋白的影響
進一步檢測細胞AMPK相關信號通路,結果發現(圖5),與油酸處理組相比,4 mmol·L-1乙酸鈉+油酸處理組P-AMPK表達水平顯著上調(P<0.05);8 mmol·L-1乙酸鈉+油酸處理組P-AMPK表達水平極顯著上調(P<0.01)。提示,乙酸鈉激活了脂肪變性細胞模型的AMPK信號通路。

與油酸處理組相比,*.P<0.05, **.P<0.01Compared with oleic acid treatment group, *.P<0.05, **.P<0.01圖5 脂肪變性細胞模型P-AMPK蛋白表達水平變化Fig.5 The P-AMPK protein expression level in steatosis cell models
2.8 不同濃度乙酸鈉對脂肪變性細胞模型脂合成代謝關鍵基因的影響
通過檢測與AMPK信號通路下游相關的關鍵脂合成代謝基因,圖6A發現,與油酸處理組相比,4 mmol·L-1乙酸鈉+油酸處理組和8 mmol·L-1乙酸鈉+油酸處理組ACC mRNA表達含量均顯著下降(P<0.05);圖6B發現,與油酸處理組相比,8 mmol·L-1乙酸鈉+油酸處理組FAS mRNA表達含量顯著下降(P<0.05),而其他乙酸鈉+油酸處理組與油酸處理組相比,均無顯著差異;圖6C發現與油酸處理組相比,2、4、8 mmol·L-1乙酸鈉+油酸處理組SCD-1 mRNA表達含量均顯著下降(P<0.05)。

與油酸處理組相比,*.P<0.05Compared with oleic acid treatment group,*.P<0.05圖6 脂肪變性細胞模型脂合成代謝基因表達Fig.6 The expression levels of lipid anabolism genes in steatosis cell models
2.9 不同濃度乙酸鈉對脂肪變性細胞模型脂分解代謝關鍵基因的影響
通過檢測與AMPK信號通路下游相關的關鍵脂分解代謝基因,圖7A發現與油酸處理組相比,4 mmol·L-1乙酸鈉+油酸處理組和8 mmol·L-1乙酸鈉+油酸處理組CPT-1 mRNA表達量均顯著上升(P<0.05);圖7B發現,與油酸處理組相比,8 mmol·L-1乙酸鈉+油酸處理組CPT-2 mRNA表達量顯著上升(P<0.05);而圖7C發現,2、4、8 mmol·L-1乙酸鈉+油酸處理組ACO mRNA表達量,與油酸處理組相比,均無顯著差異。

與油酸處理組相比,*.P<0.05Compared with oleic acid treatment group,*.P<0.05圖7 脂肪變性細胞模型脂分解代謝基因表達Fig.7 The expression levels of lipid catabolism genes in steatosis cell models
3 討 論
3.1 油酸誘導脂肪變性細胞模型的建立
脂肪肝的發生與動物肝脂質代謝的異常有關。NAFLD是一種以肝細胞內脂質過度積聚為特征的疾病。如果動物沒有得到及時治療,NAFLD就可能從單純性肝脂肪變性發展為非酒精性脂肪性肝炎,或者發展為肝纖維化、肝硬化甚至肝細胞癌。動物超重、胰島素抵抗、圈養、飲食模式改變、遺傳因素以及腸道屏障功能的紊亂都有可能導致NAFLD[11]。
目前,脂肪肝動物模型的構建通常通過高脂、高糖、四環素等毒物或藥物構建[12-13];而體外脂肪變性模型的構建通常采用油酸、脂肪酸、棕櫚酸等處理方法[14-15]。本試驗首先用不同濃度的油酸誘導BRL-3A細胞,經細胞活力測定分析發現,向細胞中添加(0.03、0.06、0.12、0.24、0.48)mmol·L-1油酸后,與對照組相比,只有0.48 mmol·L-1油酸對細胞有明顯的抑制作用,證明0.48 mmol·L-1油酸對BRL-3A細胞毒性較大,并且通過顯微鏡觀察發現細胞出現受損死亡等情況,而其他濃度的油酸對BRL-3A細胞無毒副作用。通過油紅O染色進一步檢測細胞總脂滴面積,發現與對照組相比,0.12和0.24 mmol·L-1油酸處理后細胞總脂滴面積極顯著上升。轉氨酶是肝損傷的主要標志物,主要由AST和ALT組成。其中,AST主要分布于心肌、肝、腎等組織,而ALT則主要分布于肝組織中[16]。醫學上當超過5%脂肪堆積在肝中時,就稱為脂肪肝。本試驗油酸濃度為0.06 mmol·L-1時,TG含量就增加了一倍;并且在0.12和0.24 mmol·L-1油酸處理后,TG含量極顯著上升,同時AST和ALT活性皆升高,提示肝細胞出現損傷。所以結合上述細胞總脂滴面積結果,當油酸濃度為0.12 mmol·L-1,刺激時間為24 h,即可成功構建脂肪變性細胞模型。
短鏈脂肪酸是腸道菌群膳食纖維的主要代謝產物。乙酸、丙酸和丁酸是人類腸道中最豐富的短鏈脂肪酸[17]。短鏈脂肪酸不僅是代謝底物,也是調節肝代謝的信號分子[18]。乙酸是動物機體內最主要的短鏈脂肪酸之一,占體循環內短鏈脂肪酸含量的一半以上。乙酸除了能作為能量底物和碳源參與機體內代謝活動外,還對機體脂肪代謝、食欲調節和胰島素抵抗等方面具有重要作用[19-21]。白鴿等[22]研究發現,向牛肝細胞中添加乙酸鈉,會加速肝脂肪酸的氧化。本試驗通過流式細胞術檢測細胞凋亡率,發現不同濃度的乙酸鈉對正常的BRL-3A細胞凋亡率均無影響,證明乙酸鈉對細胞無毒副作用,可用于后續研究。進一步用4和8 mmol·L-1乙酸鈉處理脂肪變性細胞模型后,均使細胞總脂滴面積、每平方毫米脂滴數、TG含量下降,AST和ALT活性皆下調,這與前人的研究結果一致。提示乙酸鈉可以顯著降低脂肪變性細胞模型的脂質堆積,緩解肝細胞損傷,具體機制有待進一步研究證實。
肝是調節脂質代謝、維持脂質穩態的重要器官。AMPK是肝細胞中能量的“傳感器”和“調節器”。AMPK可調控脂質代謝酶的表達來調節肝脂質代謝。因此,AMPK信號通路在肝脂質代謝中起中心作用[23]。研究表明,乙酸可激活AMPK,AMPK進而上調肝中脂質氧化基因的表達,減少脂肪堆積[9,24-25]。小鼠肝特異性AMPK缺失導致血漿TG含量和肝脂肪生成增加[26]。AMPK還可以通過調控骨骼肌中葡萄糖攝取和游離脂肪酸氧化,抑制肝中的糖異生、糖酵解、脂肪生成和膽固醇形成[27]。本試驗發現,用不同濃度乙酸鈉處理脂肪變性細胞模型后,4和8 mmol·L-1乙酸鈉均可激活AMPK信號通路關鍵蛋白。
TG是細胞內和血漿中脂肪酸儲存和運輸的主要形式。肝是脂肪酸代謝的中心器官,肝細胞通過從血漿中攝取和從頭生物合成的方式在肝中積累脂肪酸[28]。脂肪酸的從頭合成主要受相關關鍵酶所調控,其中,乙酰輔酶A羧化酶(acetyl-CoA carboxylase,ACC)可以將乙酰輔酶A羧化生成丙二酰輔酶A[29];脂肪酸合成酶(fatty acid synthetase,FAS)是催化脂肪酸從頭生成的最后一步[30];SCD-1是單不飽和脂肪酸合成的關鍵調控因子[31]。而脂肪酸的分解代謝受CPT-1、CPT-2等酶的調控,主要是將脂肪酸轉運到線粒體進行氧化分解,這是脂肪酸在β-氧化過程中的限速步驟[32]。本研究發現,用不同濃度的乙酸鈉處理脂肪變性細胞模型后,4和8 mmol·L-1乙酸鈉均會導致細胞ACC mRNA表達量顯著下降;8 mmol·L-1乙酸鈉會導致FAS mRNA表達量顯著下降;而2、4、8 mmol·L-1乙酸鈉均可使SCD-1 mRNA表達量顯著下降。4和8 mmol·L-1乙酸鈉會導致與脂肪酸分解代謝的相關酶CPT-1 mRNA表達量顯著上調;而8 mmol·L-1乙酸鈉會導致CPT-2 mRNA表達量顯著上調。提示,乙酸鈉主要通過CPT-1促進了脂肪變性細胞模型胞液中脂肪酸轉運到線粒體進行氧化分解,并且通過SCD-1進一步抑制了飽和脂肪酸催化合成單不飽和脂肪酸,進而降低了細胞中脂質的含量。以上結果表明,乙酸鈉可通過AMPK信號通路調控脂肪變性細胞模型脂代謝關鍵酶的活性,促使脂肪酸的從頭合成途徑被抑制,激活脂肪酸的分解途徑,進而減少BRL-3A細胞TG的合成。
4 結 論
油酸誘導的脂肪變性細胞模型,會造成肝細胞受到一定程度的損傷。而用不同濃度乙酸鈉會通過AMPK信號通路促進脂肪酸的分解代謝,進而減少TG的合成,同時可以抑制肝細胞損傷,從而緩解脂肪變性所帶來的負面影響,為今后NAFLD的治療提供了新的理論依據和切入點。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
