解毒消癥飲通過HIF-1/miR-210調節葡萄糖能量代謝抑制肝癌細胞增殖的生物學機制研究
2022-12-22 02:18:56魯琴關建華曾建偉林明和林久茂NathanielWeygant曹治云
福建中醫藥 2022年11期
魯琴,關建華,曾建偉,林明和,林久茂,Nathaniel Weygant,曹治云*
(1.福建中醫藥大學中西醫結合研究院,福建 福州 350122;2.福建省中西醫結合老年性疾病重點實驗室,福建 福州350122)
目前,我國肝癌發病率和病死率在癌癥疾病中位列第三[1]。通過手術、放化療、免疫治療、靶向治療等雖然取得一定的進步,但因復發和轉移導致的病死率仍居高不下。因此,尋找高效且低毒的抗腫瘤藥物,以此來降低復發和減少轉移是目前臨床治療中亟待解決的問題。缺氧是實體腫瘤常見的現象,腫瘤形成過程是腫瘤細胞對缺氧逐漸適應的過程,而低氧誘導因子1(hypoxia inducible factor 1,HIF-1)在這一過程中起著中樞作用[2]。HIF-1可通過形成多血管體系和提高糖酵解速率來提高肝癌細胞對低氧條件的適應性,進而抑制細胞凋亡,促進細胞增殖。miR-210(microRNA-210)是一種低氧誘導的miRNA,直接受HIF-1的調控,它的表達與HIF-1呈正相關性,目前被認為是腫瘤低氧環境下的標志性表達基因[3]。肝癌細胞在低氧的狀態下,通過調控糖酵解途徑的關鍵酶如葡萄糖轉運體蛋白、磷酸甘油酸激酶等,導致糖酵解加快及乳酸生成,降低線粒體氧化磷酸化發生水平,而此過程的調控核心單元為HIF-1/miR-210。深入探討HIF-1/miR-210誘導的能量代謝重編程具體調控機制,將為抗腫瘤藥物研究提供新的治療靶點[4-5]。
解毒消癥飲(JXY)在臨床上長期被應用于消化道腫瘤患者的輔助治療,在圍放、化療期對于消化道腫瘤中熱毒熾盛型的患者,可減少復發和轉移,延長患者生存期[6-9]。該中藥復方還可通過抑制與內皮細胞遷移和血管生成密切相關的基質金屬蛋白酶(matrix metalloproteinases,MMP)-2和MMP-9的表達,實現對低氧條件下以HIF-1為通路核心的肝癌細胞增殖及血管新生的抑制作用[10-12]。但該方通過HIF-1/miR-210回路調控能量代謝重編程具體作用機制尚未明確。本文以HIF-1/miR-210為切入點,考察JXY干預肝癌細胞在能量代謝重編程過程中相關因子的表達情況,從分子水平探討該方調控的具體作用機制。
1 實驗材料
1.1 實驗細胞 人肝癌細胞株HepG2、Hep3B、Huh7,由中國科學院細胞庫提供。
1.2 實驗藥物 JXY藥物組成:白花蛇舌草30 g,山慈菇15 g,苦參15 g,夏枯草15 g,購自福建中醫藥大學國醫堂,參照文獻[10]按質量配比制備JXY乙酸乙酯提取物。
1.3 實驗試劑 高糖DMEM培養基(美國GIBCO公司);胎牛血清、胰蛋白酶、MTT(美國Thermo Fisher Scientific公司);Hoechst 33342染色試劑盒、葡萄糖含量檢測試劑盒(北京索萊寶科技有限公司);HIF-1α、人葡萄糖轉運蛋白-1(glucose transporter 1,GLUT-1)、單羧 酸轉 運 蛋 白-4(monocarboxylate transporter 4,MCT-4)、己糖激酶(hexokinase,HK)、磷酸果糖激酶(phosphofructokinase,PFK)抗體(美國Proteintech Group公司);引物構建于浙江尚亞生物技術有限公司。
1.4 實驗儀器 超凈工作臺(蘇州凈化設備公司);低氧細胞工作站(英國Don Whitely Scientific公司);CO2培養箱、-80 ℃超低溫冰箱(美國Thermo公司);形態學顯微圖像分析系統LAS V4.1、倒置顯微鏡系統(德國Leica儀器有限公司);Countes?全自動細胞計數儀(美國Life公司)。
2 實驗方法
2.1 低氧與常氧培養條件 20% O2、5% CO2、37 ℃完全飽和濕度條件下常規培養為常氧培養條件;使用低氧細胞工作站(0.1% O2、5% CO2、94% N2的混合氣),37 ℃條件下培養為低氧條件。
2.2 干預條件篩選
2.2.1 MTT法觀察低氧和常氧條件對肝癌細胞活力的影響 將肝癌細胞株HepG2、Hep3B、Huh7分別按1.0×104個/mL均勻接種于96孔細胞培養板中,并同時分別置于低氧或常氧條件下培養,設置3個復孔。分別干預3、6、9、12、24、36、48 h后,棄去板中上清液,每孔加入10 μL的MTT(初始濃度:0.5 mg/mL)后繼續培養4 h,加入含有1% NH4OH的DMSO溶解晶體至48 h后完全結束該實驗。酶標儀上于492 nm 測定吸光度值。
2.2.2 Western blot法檢測低氧和常氧條件對HIF-1α蛋白表達量的影響 取對數生長期的肝癌細胞接板,分別同時置于低氧或常氧條件下培養,分別在不同時間段(0.5、1、3、6、9 h)收集并提取蛋白。在干預時間結束后提取蛋白,進行電泳、轉膜、一抗冷庫孵育過夜,TBST洗膜,二抗搖床孵育2 h,通過Bio-Rad成像系統對蛋白指標進行成像,并使用Image J軟件對獲得的蛋白數據分析整理。
2.2.3 MTT法觀察低氧和常氧條件下不同濃度JXY對肝癌細胞活力的影響 將肝癌細胞株HepG2、Hep3B、Huh7分別按1.0×104個/mL均勻接種于96孔細胞培養板中,同時分別置于低氧或常氧條件下培養,在細胞匯合度到達60%~70%時予以不同濃度JXY(0、0.05、0.1、0.2 mg/mL)干預,并設置3個復孔。分別干預6、9、12、24 h后,棄去板中上清液,每孔加入10 μL的MTT(初始濃度:0.5 mg/mL)后繼續培養4 h,加入含有1% NH4OH的DMSO溶解晶體至48 h后完全結束該實驗。酶標儀上于492 nm測定吸光度A值,按以下公式計算:

2.3 干預實驗
2.3.1 細胞分組 將肝癌細胞分為常氧0 mg/mL組、常 氧0.1 mg/mL組、低 氧0 mg/mL組、低 氧0.1 mg/mL組。在細胞匯合度到達60%~70%時予以不同濃度JXY(0、0.1 mg/mL)干預,同時根據實驗要求設置不同干預條件。
2.3.2 細胞爬片實驗觀察細胞形態 將蓋玻片消毒滅菌后放入6孔板中,按“2.3.1”項下分組干預后,將蓋玻片放置于蘇木素溶液中染色2 min,超純水清洗3遍后,置于自來水中返藍10 min。再將蓋玻片放于伊紅溶液中染色1 min,超純水清洗3遍,晾干,中性樹膠封片,用顯微鏡采集圖片。
2.3.3 劃痕實驗觀察肝癌細胞生長遷移 細胞接種到12孔板中后置于37 ℃、5%CO2飽和濕度的培養箱中培養,第2天當細胞匯合度達到80%~90%時,用無菌200 μL移液槍槍頭尖端從左至右水平輕輕刮擦,經無菌PBS清洗后加入2 mL/孔的完全DMEM培養基,再置于顯微鏡下拍照。按“2.3.1”項下分組干預,每隔24 h拍照1次,直至對照組劃痕全部閉合。
2.3.4 Hoechst染色法觀察肝癌細胞凋亡情況 細胞按照1.0×105個/mL接種在6孔板后置于37 ℃、5%CO2飽和濕度的培養箱中培養,貼壁后按“2.3.1”項下分組干預24 h,棄去上清液,PBS洗2遍后用福爾馬林進行固定,加入400 μL的10 μg/mL Hoechst 33342染色并避光,15 min后PBS洗3遍,用熒光顯微鏡拍照。
2.3.5 細胞遷移實驗觀察肝癌細胞遷移侵襲能力 取對數生長期細胞消化,使用無血清DMEM培養基重懸后稀釋至所需細胞密度,24孔板下加入600 μL含10%FBS的DMEM培養基,并取等體積細胞懸液加入Transwell小室,按“2.3.1”項下分組干預,孵育24 h后取出小室用PBS清洗2遍,4%多聚甲醛固定30 min后再用PBS清洗2遍,加入0.1%結晶紫染色30 min,最后用PBS清洗2遍并用棉簽輕輕擦去表面未遷移細胞后,置于光學顯微鏡下拍照觀察。
2.3.6 可見分光光度法檢測肝癌細胞葡萄糖濃度 按“2.3.1”項下分組干預24 h,收集細胞到離心管內,離心后棄上清;按照一定比例加入蒸餾水后用超聲波破碎細胞(冰浴,功率20%,超聲3 s,間隔10 s,重復30次),放置于沸水浴中煮沸10 min,冷卻后,25 ℃下8 500 r/min離心10 min,取上清液備用。分光光度計預熱30 min,蒸餾水調零,按照葡萄糖含量檢測試劑盒說明書,在樣本中加入提前配置好的試劑混勻后,置于37 ℃水浴保溫15 min,于505 nm波長處讀取吸光值。
2.3.7 Agilent Seahorse XF能量代謝檢測肝癌細胞能量代謝水平 按照XF ATP RATE ASSAY檢測步驟說明書進行實驗,按“2.3.1”項下分組干預后,利用Seahorse XFe9檢測系統,檢測三磷酸腺苷(ATP)水平。
2.3.8 Western blot法檢測肝癌細胞GLUT1、MCT4、HK、PFK蛋白表達量 取對數生長期的肝癌細胞按照1×105個/mL的密度接種于6孔板中,當細胞匯合度達到50%~60%時,按“2.3.1”項下分組干預24 h。干預后提取蛋白,進行電泳、轉膜、一抗冷庫孵育過夜,TBST洗膜,二抗搖床孵育2 h,通過Bio-Rad成像系統對蛋白指標進行成像,并使用Image J軟件對獲得的蛋白數據分析整理。
2.3.9 熒光定量PCR法檢測肝癌細胞HIF-1α、miR-210 mRNA表達水平 取對數生長期的細胞進行接板并置于37 ℃、5%CO2飽和濕度的培養箱中培養,待細胞貼壁后,按“2.3.1”項下分組干預24 h,收集肝癌細胞,按經典RNA提取試劑盒說明書提取總RNA,并用常規逆轉錄方法將mRNA轉錄成cDNA,SYBR Green I做為熒光染料,而后根據Light CyelerDNA Master SYBR Green I說明書進行熒光定量PCR擴增。HIF-1α正引:GCAGCAACGACACA GAAACT,反引:TGCAGGGTCAGCACTACTTC;miR-210正引:AATAATCGCTGTGCGTGTGAC,反引:AG TGCAGGGTCCGAGGTATT。檢測HIF-1α和miR-210 mRNA水平的表達。
2.4 統計學方法 采用SPSS 24.0軟件包進行數據分析。計量資料符合正態分布以(±s)表示,多組間比較采用單因素方差分析,兩兩比較采用Student-Newman-Keuls法。P<0.05為差異有統計學意義。
3 結 果
3.1 不同干預時間對低氧和常氧條件下肝癌細胞增殖活性的影響 MTT結果顯示,在低氧條件下培養的3株肝癌細胞增殖活性較常氧條件下明顯增強,并隨著培養時間的增長而不斷升高。其中,Huh7和HepG2肝癌細胞在低氧條件下24 h細胞增殖活性達到最高,Hep3B細胞增殖活性在24 h仍在緩慢上升,至36 h達到峰值。見圖1。
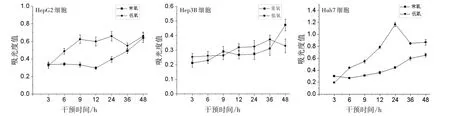
圖1 不同干預時間對低氧和常氧條件下肝癌細胞增殖活性的影響
3.2 不同干預時間對低氧和常氧條件下肝癌細胞HIF-1α蛋白表達的影響 Western blot結果顯示,3株肝癌細胞在低氧條件下HIF-1α蛋白表達量較常氧條件下明顯上調,且隨著培養時間的延長蛋白表達量也逐漸升高,說明低氧細胞模型已成功建立。因此結合MTT實驗結果,后續實驗將選取低氧24 h為JXY干預時間點。見圖2。
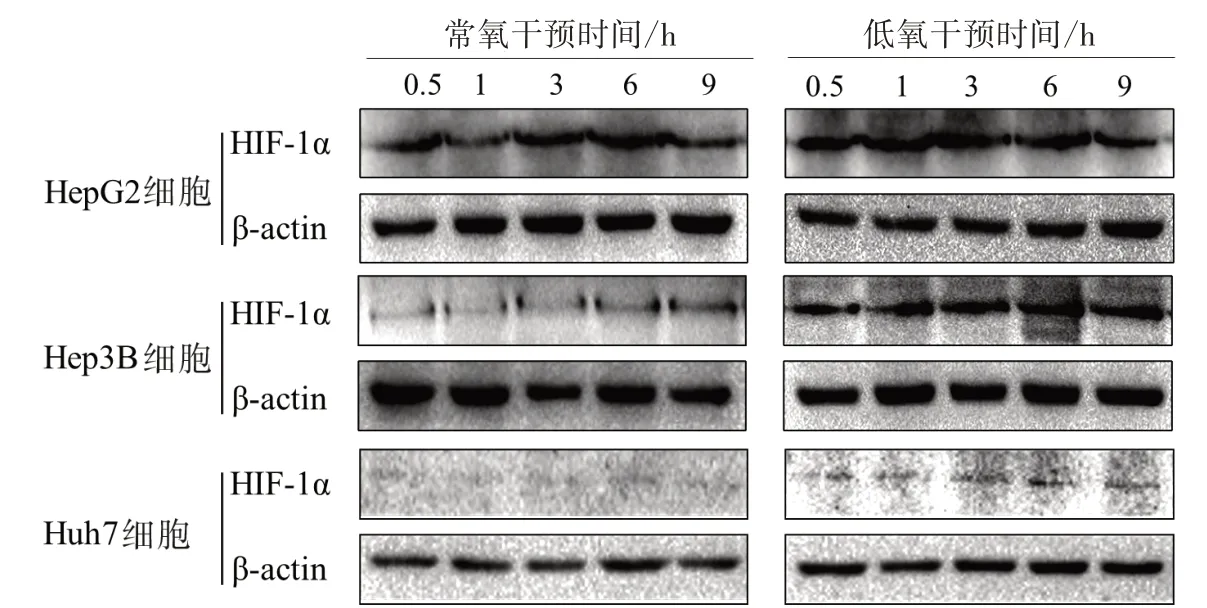
圖2 不同干預時間對低氧和常氧條件下肝癌細胞HIF-1α蛋白表達的影響
3.3 不同濃度JXY和不同干預時間對低氧和常氧條件下肝癌細胞活力的影響 JXY干預后在低氧和常氧條件下均能抑制肝癌細胞增殖,其在低氧條件下的抑制效果顯著高于常氧條件組,并且呈時間-劑量依賴性,見圖3。
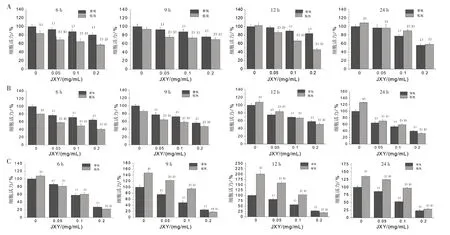
圖3 不同濃度JXY和不同干預時間對低氧和常氧條件下肝癌細胞活力的影響
3.4 常氧和低氧條件下JXY對肝癌細胞形態的影響 細胞予以不同濃度JXY干預后置于不同條件下培養,結果發現在0 mg/mL濃度下,細胞在低氧條件下較常氧條件形態發生明顯改變;而在相同條件下,高濃度JXY干預可明顯抑制低氧條件導致的肝癌細胞的形態改變。見圖4。
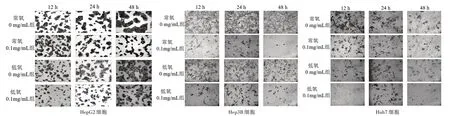
圖4 4組肝癌細胞不同干預時間細胞爬片圖(×200)
3.5 常氧和低氧條件下JXY對肝癌細胞轉移能力的影響 劃痕實驗結果表明,予以0 mg/mL JXY濃度下的Hep3B細胞遷移能力在不同條件下無明顯變化,HepG2和Huh7細胞在常氧條件下的遷移能力較低氧條件下更強;而高濃度能顯著抑制2種條件下3株肝癌細胞的遷移能力。見圖5。

圖5 4組肝癌細胞劃痕愈合圖(×100)
3.6 低氧和常氧條件下不同濃度JXY對肝癌細胞凋亡能力的影響 選取0.1 mg/mL作為后續生物學機制研究的濃度,檢測常氧和低氧條件細胞凋亡,染色結果表明,在低氧和常氧條件下JXY干預均能促進肝癌細胞凋亡,低氧條件下發生凋亡的細胞數量更多。見圖6。
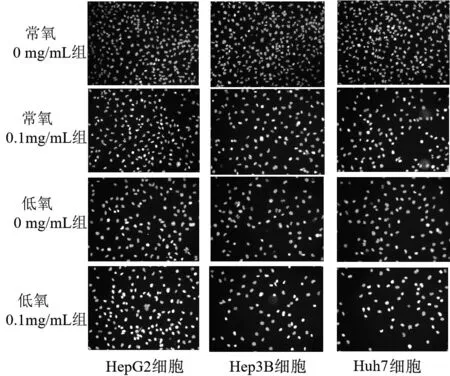
圖6 4組不同肝癌細胞Hoechst染色圖(×200)
3.7 常氧和低氧條件下JXY對肝癌細胞轉移能力的影響 細胞遷移實驗表明,高濃度JXY對肝癌細胞侵襲遷移能力具有顯著抑制作用。見圖7。

圖7 4組肝癌細胞遷移作用圖(×100)
3.8 4組肝癌細胞HIF-1α蛋白表達及HIF-1α、miR-210 mRNA表達水平比較 Western blot結果顯示,在0 mg/mL JXY濃度下,2株肝癌細胞在低氧條件下產生的HIF-1α顯著高于常氧條件組,而高濃度JXY能顯著下調在不同條件下HIF-1α的蛋白表達,見圖8。同時熒光定量PCR結果顯示,在0 mg/mL JXY濃度下,低氧條件下HIF-1α的累積促進了miR-210 mRNA水平的表達,而高濃度JXY在抑制HIF-1α的同時也抑制了miR-210 mRNA水平的表達,兩者呈正相關。見圖9。

圖8 4組肝癌細胞HIF-1α蛋白表達比較
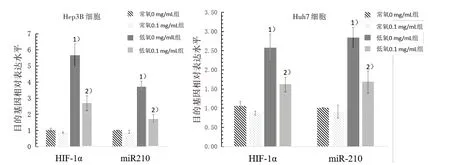
圖9 4組肝癌細胞HIF-1α和miR-210 mRNA表達水平比較
3.9 4組肝癌細胞葡萄糖含量和細胞能量代謝比較 葡萄糖含量檢測實驗和細胞能量代謝檢測結果顯示,在0 mg/mL JXY濃度下,低氧條件組較常氧條件組葡萄糖攝取含量和糖酵解途徑的ATP產量增加;而JXY干預對其具有顯著的抑制效果。見圖10。

圖10 4組肝癌細胞葡萄糖含量和細胞能量代謝比較
3.10 4組肝癌細胞GLUT-1、HK、PFK和MCT-4蛋白表達比較 Western blot結果表明,低氧條件下由于能量代謝重編程引起的GLUT-1、HK、PFK和MCT-4等相關蛋白的高表達,JXY干預后均能起到顯著的抑制作用。見圖11。
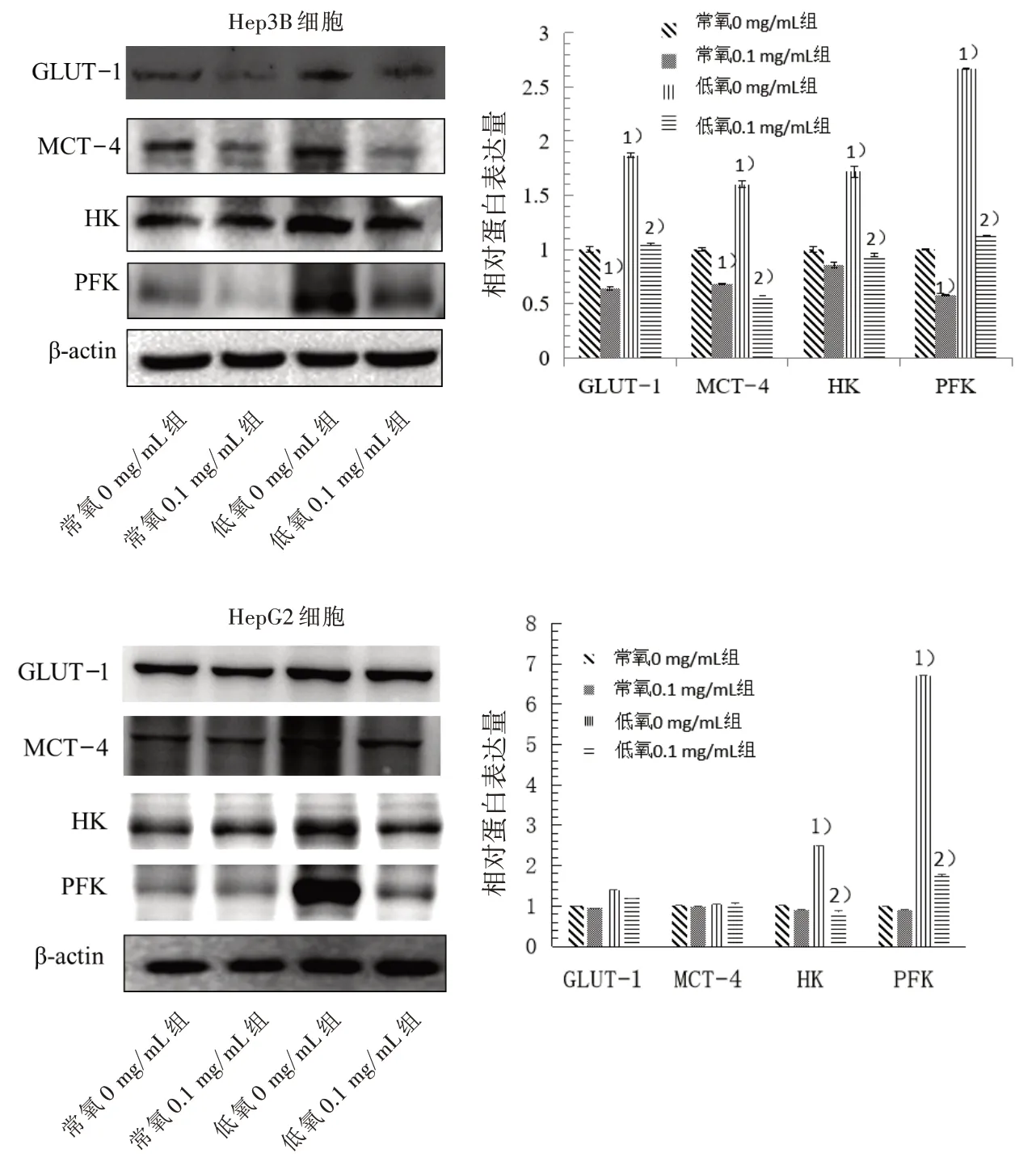
圖11 4組肝癌細胞GLUT-1、HK、PFK和MCT-4蛋白表達比較
4 討 論
腫瘤細胞為了適應在低氧環境下對能量的需求,會對能量進行代謝重編程,即弱化線粒體的氧化磷酸化途徑,取而代之選擇糖酵解,主要表現為葡萄糖攝取量顯著提高、糖酵解活性明顯增強及乳酸大量堆積,又被稱為Warburg效應。腫瘤細胞代謝重編程的間接表現是葡萄糖攝取量增加,因為葡萄糖不能通過細胞膜,在很大程度上依賴于葡萄糖轉運體的運輸,腫瘤為攝取葡萄糖會應激性高表達GULT,其中GULT-1和GULT-3是HIF-1轉錄調節的靶基因,其轉錄水平表達升高與HIF-1直接相關。糖酵解過程中的3個限速酶HK、PFK和PK是糖酵解途徑的3個關鍵調節點,其轉錄水平和激酶活性受HIF-1的直接調控[5]。而且糖酵解不僅可為細胞增殖提供不可缺少的中間代謝產物,如6-磷酸葡萄糖、6-磷酸果糖、磷酸烯醇式丙酮酸及丙酮酸等,作為細胞合成核酸、氨基酸、脂肪酸等細胞結構生物大分子不可或缺的前體物質,又可快速產生一定量的ATP用于維持細胞能量穩態,使腫瘤細胞達到生長與代謝的空前平衡[13],而此過程的調控核心單元為HIF-1/miR-210回路。
實體腫瘤組織內部處于低氧狀態,這時細胞通常會通過啟動相關信號調節通路,從而改變分子的DNA轉錄模式以應對不良微條件,應對這種改變的核心分子是HIF-1。HIF-1由對氧敏感的HIF-1α亞基和對氧不敏感的HIF-1β亞基組成,在正常氧分壓下HIF-1β是具有活性的分子,而HIF-1α則通過PHD羥化其分子上的脯氨酸后進入蛋白酶體降解通路最終被酶解代謝[14-15]。在腫瘤低氧的微環境中,HIF-1α的降解途徑被阻斷,直接導致HIF-1α水平的累積。
人miR-210位于染色體1lp15.5,序列為“CUGU GCGUGUGACAGCGGCUGA”。常氧條件下,內源性miR-210水平維持在非常低的狀態[16]。低氧條件下,HIF-1α的表達應激性上調,HIF-1α啟動子區域的缺氧反應元件可和miR-210序列(6~8個核苷酸)配對,抑制miR-210降解的同時上調miR-210轉錄產物[17],過表達miR-210又可削弱PHD mRNA的翻譯水平,降低HIF-1α的羥化,抑制其降解。實驗表明腫瘤細胞在缺氧處理48~72 h后,上調的HIF-1α能夠促進VEGF和miR-210的表達,而抑制miR-210 mRNA水平則會降低HIF-1α的蛋白水平[18]。這表明HIF-1α與miR-210之間是一個正反饋回路的調控關系[19]。
研究表明夏枯草的主要成分夏枯草總黃酮可同時抑制線粒體氧化磷酸化和糖酵解途徑抑制肝癌誘導肝癌細胞SMMC-7721凋亡,苦參素抑制HepG2細胞糖酵解過程抑制肝癌細胞增殖,此外紅豆杉、白頭翁、穿心蓮、黃芩、半枝蓮等多種清熱解毒中藥都具有抑制腫瘤糖酵解途徑中關鍵酶的作用[20-22]。這為中藥抗腫瘤藥物研究提供了新的治療靶點。而清熱解毒中藥通過HIF-1α/miR-210正反饋回路調控腫瘤細胞能量代謝重編程過程具體作用機制目前尚未見相關報道。
中醫學認為癌癥多為內虛邪擾,陰陽嚴重失調,瘀、毒、虛、六淫等諸多因素均與癌癥相關。治療上一般采取扶正祛邪,調理各臟腑機能,增強體質和免疫力。而解毒消癥飲入肝經,方中白花蛇舌草、夏枯草、山慈菇、苦參四藥性味均屬苦、甘、寒,是清熱解毒法中藥驗方,可通過疏泄癌腫堵塞的肝經,化解日久化熱所致小便短赤、大便干結、舌紅苔厚等熱證。
本研究通過建立低氧和常氧細胞模型,對比模擬肝癌細胞生長環境,證明低氧和常氧條件下腫瘤細胞能量代謝重編程的應答機制在形態及功能方面的顯著差異,同時探討JXY通過HIF-1/miR-210回路調控肝癌細胞能量代謝重編程的具體作用機制。結果顯示肝癌細胞在低氧條件下的增殖活性較常氧條件下明顯增強,并隨著培養時間的增長而不斷升高。其中,Huh7和HepG2肝癌細胞在低氧條件下24 h細胞增殖活性達到最高,Hep3B細胞增殖活性在24 h仍在緩慢上升,至36 h達到峰值。因此后續實驗選取低氧24 h為JXY干預時間點。肝癌細胞在低氧條件下HIF-1α蛋白表達量較常氧條件下明顯上調,且隨著培養時間的延長蛋白表達量也逐漸升高。而在JXY干預后,其在低氧和常氧條件下均能抑制肝癌細胞增殖,且在低氧條件下的抑制效果顯著高于常氧條件組,呈時間和劑量依賴性。同樣地,JXY干預在低氧條件下促進肝癌細胞凋亡作用顯著。低氧條件下JXY干預對肝癌細胞的形態學改變、侵襲遷移能力和能量代謝重編程過程中所需的葡萄糖攝取,以及相關因子HIF-1α、GLUT-1、HK、PFK和MCT-4蛋白表達量和HIF-1α調控的miR-210 mRNA表達水平均有顯著的抑制作用。由此猜想,JXY可能通過抑制HIF-1/miR-210正反饋回路,抑制腫瘤細胞糖酵解過程,從而抑制糖酵解過程中所需葡萄糖攝取和ATP生成,進而抑制GLUT-1、HK、PFK、MCT-4的表達。最終調整整個能量代謝重編程過程,達到抑制腫瘤細胞增殖、侵襲、轉移等目的。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
中成藥(2016年8期)2016-05-17 06:08:14
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
