藍莓AP2/ERF基因家族鑒定及其在休眠解除過程中的表達分析研究
2022-12-09 07:16:36馬芮李瑞雪鄭煜聆李永強
浙江農業科學 2022年12期
關鍵詞:分析
馬芮,李瑞雪,鄭煜聆,李永強,2*
(1.浙江師范大學 化學與生命科學學院,浙江 金華 321004;2.浙江省特色經濟植物生物技術研究重點實驗室,浙江 金華 321004)
APETALA2/ethylene response factor(AP2/ERF)轉錄因子家族作為植物中最大的轉錄因子家族之一,參與植物體內各種信號傳導和生長發育調節[1-3]。AP2/ERF家族每個成員包含至少一個由60~70個氨基酸組成的AP2結構域,包括AP2、ERF和RAV亞家族,根據AP2結構域的數目和序列的相似程度將AP2/ERF超家族分為AP2家族、ERF家族和RAV家族[4]。其中AP2家族具有兩個AP2/ERF保守結構域,而ERF家族成員包含一個AP2/ERF保守結構域。AP2結構域首次在擬南芥(Arabidopsisthaliana)中被發現AP2蛋白中的重復基序,能夠參與花的發育[5]。RAV家族蛋白包含一個B3結構域和一個AP2/ERF結構域[6]。研究表明,ERF轉錄因子在植物的生長發育以及多種逆境脅迫應答反應中發揮著重要調控作用[7-8]。在乙烯信號轉導級聯反應中一個重要的轉錄因子家族是乙烯響應因子(ERFs)。他們作為AP2/ERF超家族的成員,由EILs基因激活并進一步表達,部分ERFs激活GCC-box靶基因,激活功能基因進行各種代謝反應。前人研究表明,從煙草(Nicotianatabacum)中分離出的含有保守ERF結構域的乙烯響應元件結合蛋白(ERF),它可以與乙烯響應元件GCC-box結合,調控乙烯相關基因的表達[9]。
目前,在番茄中已有多個ERFs被鑒定出來,部分可以正向調控果實成熟,而也有部分乙烯響應因子對成熟調控起到負調控作用[10]。研究表明,AP2/ERF超家族成員廣泛參與植物乙烯信號途徑和花芽萌發以及果實成熟,除此之外還參與植物發育過程中的脅迫、代謝、衰老等。AP2/ERF能夠響應各種生物和非生物脅迫,進而調節植物的生長發育,例如低溫[11]、干旱、高溫[12-13]、鹽脅迫等。AP2家族中的基因被證實可以決定玉米穗的分生組織[14],AP2/ERF轉錄因子通過轉錄和翻譯等多種機制參與調控生長、發育、激素和應激反應等多種生物學過程的基因表達[15-16]。有研究證明,過表達DREB2C時會導致植物種子推遲萌發[17]。Sl-ERF2轉基因番茄種子比野生型提早萌發,提高了植株對乙烯的敏感性[18]。ERFs基因廣泛存在于高等植物中,可以通過與DNA直接結合,達到調控下游基因轉錄的目的,也可以調控各種信號轉導途徑[19-21]。當山毛櫸(Faguslongipetiolata)處于休眠狀態時,ERF1在種胚中的表達量較低,但休眠解除時,其表達量顯著升高[22]。
藍莓(Vacciniumspp.)是杜鵑花科(Ericaceae)越橘屬(Vaccinium)植物,不僅具有較高的營養價值,還具有較高的經濟價值,在提高免疫力、增強心肺功能等方面作用顯著,被譽為“漿果之王”[23]。由于AP2/ERF轉錄因子在植物生長發育過程中起到重要作用,本研究借助四倍體藍莓品種Draper的基因組數據,對藍莓ERF基因進行鑒定,共得到160個家族成員。進行了多序列比對,保守基序、基因結構、啟動子分析,通過轉錄組數據分析藍莓AP2/ERF基因家族成員在花芽休眠解除過程中的表達情況。研究結果為進一步研究ERF基因在果樹芽休眠調控中的功能奠定基礎,為藍莓產業的發展提供理論基礎。
1 材料與方法
1.1 序列數據獲取
通過ArabidopsisInformation Resource(TAIR)數據庫(http://www.arabidopsis.org),獲取擬南芥AP2/ERF基因家族成員,下載蛋白質序列作為參考序列,從越橘基因組數據庫(GENOME DATABASE FOR VACCINIUM,GDV)中獲取藍莓蛋白數據庫[24]。以擬南芥ERF蛋白序列作參考,使用TBtools軟件對蛋白數據庫進行BlastP比對[25]。
1.2 藍莓AP2/ERF基因家族成員的鑒定
選擇從上述程序獲得的獨特序列作為候選AP2/ERF家族蛋白序列。通過CD-Search(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)排除一些不包含AP2結構域的蛋白序列,通過MEME將篩選出的藍莓ERF蛋白序列進行保守基序分析,確定藍莓AP2/ERF基因家族成員。
1.3 藍莓ERF蛋白的理化性質
利用ProtParam在線軟件預測藍莓中ERF蛋白的氨基酸長度、分子量(MW)、理論等電點(pI)、親水性(GRAVY)。應用WoLF PSORT軟件分析ERF蛋白序列,進行亞細胞定位預測[26]。
1.4 多序列比對及進化分析
通過MEGA5.0分析軟件對篩選得到的160個藍莓ERF全長蛋白序列進行多序列比對并構建系統發育樹,方法為:鄰接法(Neighbor-Joining method)Bootstrap 1 000次重復[27],借助在線工具EvolView(https://www.evolgenius.info/evolview/#/)繪制進化樹圖[28]。
1.5 保守基序分析
利用MEME(http://meme-suite.org/tools/meme)在線工具進行蛋白保守結構域預測分析,將保守基序(motif)大小設定為默認參數,輸出結構域最大數為5,motif寬度最長為60,其余為默認[29]。
1.6 染色體定位及共線性分析
根據藍莓基因組信息將160個VcERF基因定位到染色體上。通過運行BlastP來檢索藍莓基因組中的同源基因對(E< 10-5,前五個匹配項)。然后將同源對用作MCScanX的輸入文件,分析基因復制的類型[30-31]。采用Circos軟件作圖[32]。
1.7 基于轉錄組的VcERFs表達模式分析
根據藍莓花芽休眠時期的轉錄組表達數據,獲得VcERFs在花芽內休眠解除、生態休眠解除的表達量數據,將FPKM值(fragments per kilobases per millionreads)進行log2a標準化處理,使用TBtools軟件的“HeatMap”功能,構建藍莓ERF基因的表達熱圖[25]。
1.8 啟動子順式作用元件分析
根據藍莓ERF基因各轉錄組表達情況,篩選可能有重要調控作用的藍莓ERF候選基因。通過藍莓基因組文件提取VcERFs編碼區上游2 000 bp的啟動子序列,使用PlantCARE(http://bioinfoatics.psb.ugent.be/webtools/plantcare/html/)網站分析相關藍莓ERF基因啟動子的順式作用元件,將得到的順式作用元件進行篩選和統計,用熱圖展示順式作用元件的類型及數量[33]。
2 結果與分析
2.1 藍莓ERF鑒定及蛋白理化性質
通過序列比對及結構域分析,最終篩選獲得160個藍莓ERF蛋白(表1)。使用ExPasy網站在線工具對VcERF蛋白的理化特性進行分析,結果顯示,ERF蛋白的氨基酸數量均在154~786個,分子量17.6~86.5 ku,等電點4.71~10.75。不穩定系數變化范圍在32.58~74.54,ERF蛋白大部分處于不穩定形態,親水性值在-1.000至-0.312,說明均為親水蛋白。亞細胞定位預測分析發現,VcERF蛋白定位在細胞的各個部位,主要定位在細胞核,少數成員定位在細胞質、葉綠體、線粒體和細胞基質。
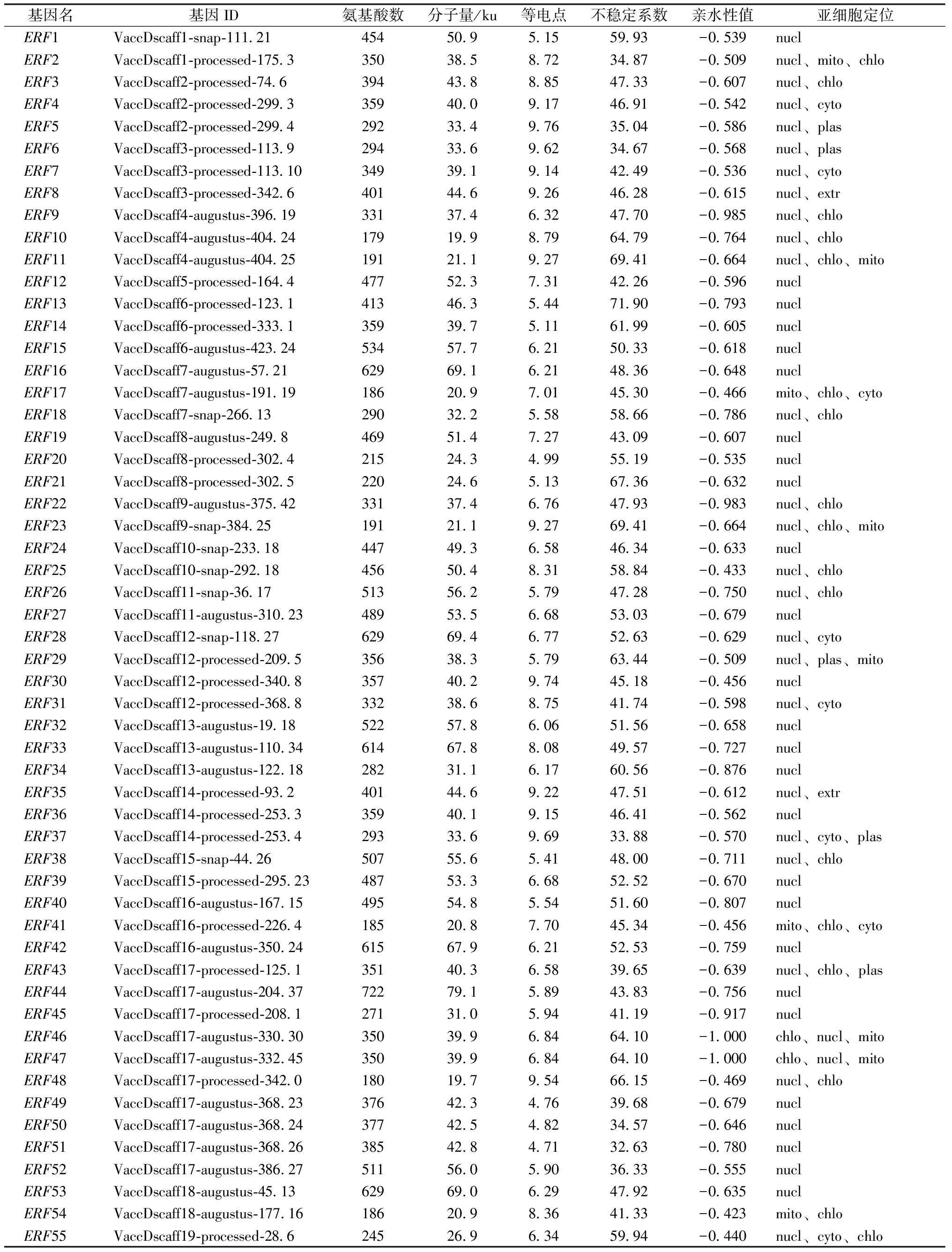
表1 藍莓ERF蛋白基本信息
2.2 藍莓AP2/ERF系統發育進化樹
為更深入地了解藍莓AP2/ERF轉錄因子同源進化關系,通過MEGA5.0軟件構建了160個藍莓ERF蛋白成員的系統進化樹,系統進化樹分枝顯示(圖1),AP2/ERF家族成員被分為3個亞族,藍莓ERF分布在AP2家族、RAV家族和ERF家族中,AP2家族含有54個ERF蛋白,ERF家族所含的藍莓成員最多,共82個,RAV家族中有24個ERF蛋白,此類成員含有一個AP2結構域和一個B3結構域。
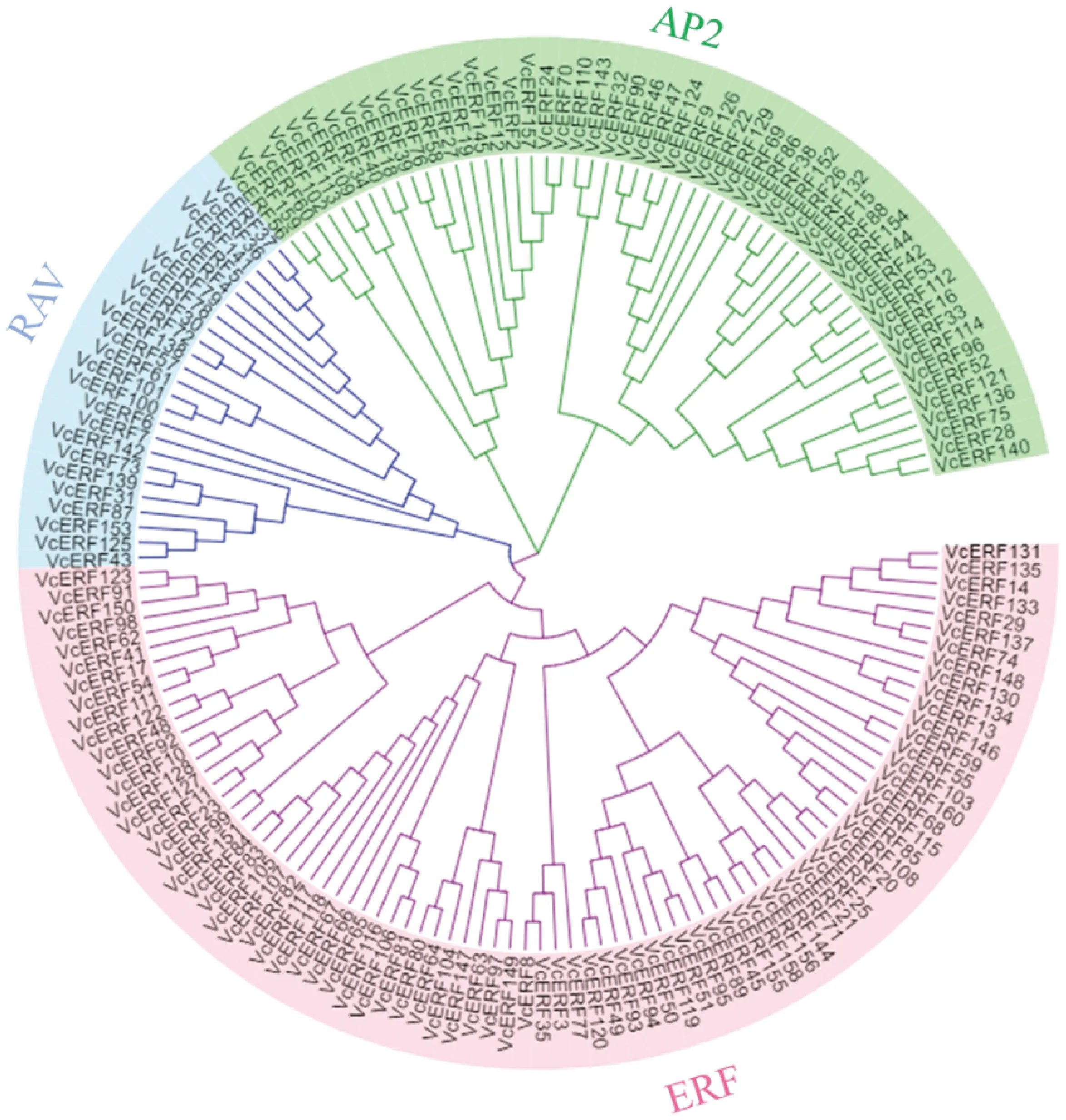
圖1 藍莓AP2/ERF基因家族成員系統進化樹分析
2.3 藍莓ERF蛋白結構和保守基序分析
為更深入地了解VcERFs中基序(motif)組成的多樣性,利用MEME預測藍莓ERF蛋白質保守基序,共得到5個保守基序(圖2),結果表明,所有VcERF蛋白均存在高度保守的AP2結構域(motif 1),motif 1作為DNA的結合模塊,對ERF蛋白的功能至關重要。根據保守基序的分布特點,可將藍莓ERF蛋白分為3組,Ⅰ組成員最多,含有82個ERF蛋白成員,Ⅱ組含有54個,Ⅲ組含有24個。從圖2中可知,每條ERF蛋白序列所含有的保守基序的數量以及類型都存在或多或少的差異,其在分布上也存在差異,可能表明每個基序的功能有所差異,其中Ⅲ組成員含有的保守域數量最多,Ⅱ組成員除特有的AP2結構域外,還含有一個特殊的B3結構域(motif 5)。
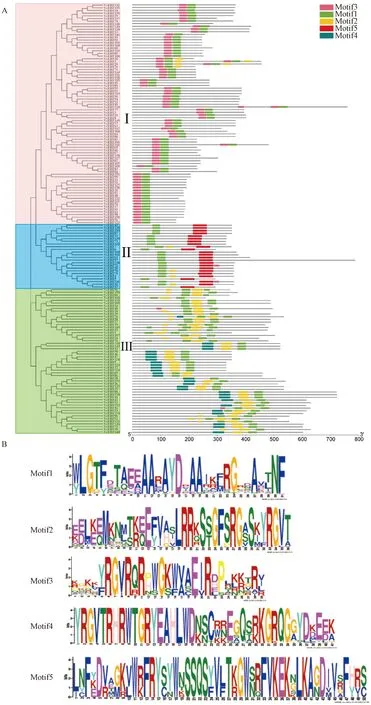
A—藍莓VcERF進化關系及保守基序;B—藍莓VcERF Motif序列。
2.4 ERF基因的共線性分析
為了進一步探討藍莓AP2/ERF基因家族的起源和可能的進化機制,將藍莓自身蛋白比對的結果輸入MCScanX,輸出同源配對文件,分析VcERFs的組內同源配對情況發現,VcERF的同源配對非常復雜,除VcERF5、VcERF6、VcERF15、VcERF21、VcERF23、VcERF26、VcERF27、VcERF38、VcERF39、VcERF50、VcERF51、VcERF52、VcERF58等30個基因沒有同源配對外,其他基因家族成員均存在線性關系,見圖3中紅色連線。藍莓AP2/ERF基因家族共有45條染色體上存在共線性基因,其中17、27號染色體上共線性基因最多,均有7對共線性基因,其次21、26號染色體上均有6對共線性基因。
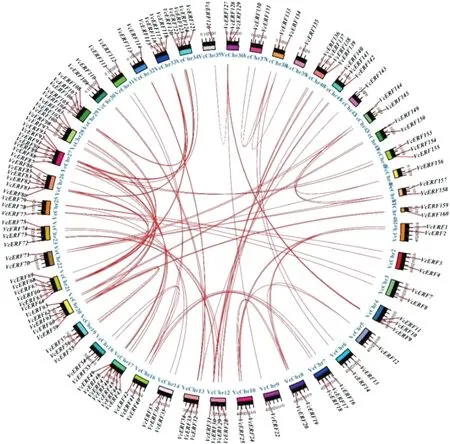
圖3 藍莓ERF基因在染色體上的分布和共線性
2.5 基于轉錄組數據分析VcERF基因的表達特性
基于已有的藍莓轉錄組數據分析AP2/ERF基因家族成員在花芽休眠解除下的表達模式(圖4),相同亞家族VcERF在休眠解除過程中的表達模式并不相似,暗示其可能在花芽休眠解除過程中發揮不同的作用。ERF家族成員VcERF51、VcERF115、VcERF119、VcERF74、VcERF95、VcERF14、VcERF29、VcERF137、VcERF48、VcERF49、VcERF148、VcERF103在休眠解除過程中表達量較高;VcERF146、VcERF50、VcERF99、VcERF122、VcERF133、VcERF82、VcERF81、VcERF105、VcERF117在內休眠初期表達量水平很高,在內休眠解除過程中下調;其中VcERF81、VcERF105、VcERF117、VcERF65、VcERF106在生態休眠解除過程中的表達量升高,最終也沒有恢復到內休眠初期的表達量水平。
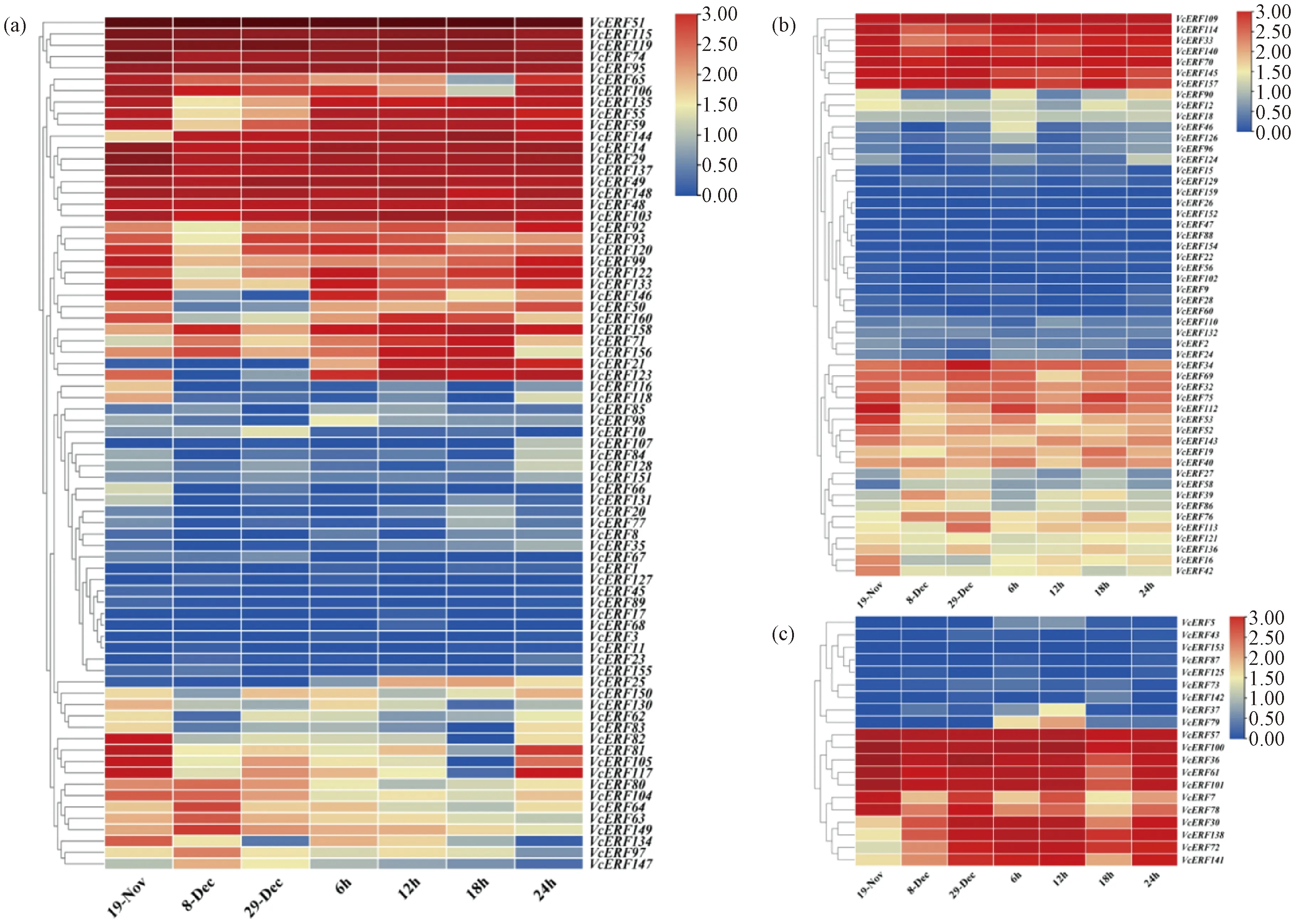
a—ERF家族;b—AP2家族;c—RAV家族。
AP2家族成員VcERF109、VcERF114、VcERF33、VcERF140、VcERF70、VcERF145、VcERF157在休眠解除過程中表達量較高;VcERF112、VcERF53的表達量在內休眠解除過程中下調。
第Ⅲ類RAV家族成員VcERF5、VcERF43、VcERF153、VcERF87、VcERF125、VcERF73、VcERF142在內休眠及生態休眠解除過程中幾乎不表達;VcERF30、VcERF138、VcERF72、VcERF141隨著內休眠的解除表達量上升;VcERF30、VcERF141的表達量在生態休眠解除過程中先下降后上升。
2.6 啟動子順式作用元件分析
為進一步分析藍莓AP2/ERF家族基因在啟動子區域的調控元件特點,對10個ERF基因上游2 000 bp片段進行啟動子順式作用元件分析。結果顯示(圖5),藍莓ERF啟動子區域含有大量光響應元件和激素響應元件,包括光反應(Box 4、ATCT-motif、MRE、SP1、G-box)等,脫落酸(ABRE)、赤霉素(P-box、TATC-box、GARE-motif)、生長素(TGA-element)、茉莉酸甲酯(CGTCA-motif、TGACG-motif)、水楊酸(TCA-element)等,表明藍莓ERF基因可能在光響應和激素響應中起重要作用。除此之外,藍莓ERF基因還含有脅迫響應元件,如低溫(LTR)、防御應激(TC-rich repeats)。
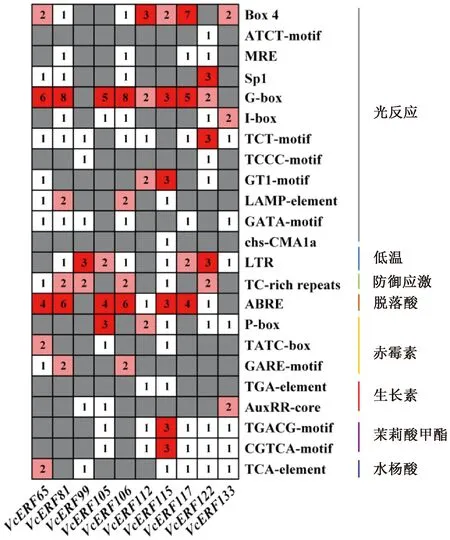
圖5 藍莓ERF基因啟動子的順式作用元件分析
3 討論
本研究中采用生物信息學方法從藍莓基因組數據庫中鑒定出160個ERF基因,分布于藍莓48條染色體中的45條上,亞細胞定位預測顯示ERF基因主要定位于細胞核。通過生信分析發現,最長的VcERF基因編碼786個氨基酸,最短的編碼154個氨基酸,分子量分布在17.6~86.5 ku,對VcERF基因的結構分析表明,雖然VcERF基因的長度存在較大差異,但總體上同一亞類的基因具有相似的基因結構,這說明VcERF基因在進化上是保守的。為了進一步研究藍莓與擬南芥中AP2/ERF家族的同源進化關系,利用鄰接法構建AP2/ERF基因家族系統發育樹,并根據其分類特點將這些基因分為3組。擬南芥中的一些AP2/ERF基因家族成員功能已經得到驗證,根據進化關系中基因的同源性,推測與擬南芥位于同一組的藍莓ERF基因也具有與其相同或相類似的功能。藍莓基因組中之所以存在如此豐富的AP2/ERF基因家族成員,一方面可能是由于其屬于多倍體物種,本研究用于AP2/ERF基因家族生物信息學預測的基因組來源于四倍體藍莓品種Draper,多倍體化會影響基因家族的分布和大小;另一方面可能與基因組加倍事件有關,藍莓基因組經歷了至少1次全基因組重復[24]和大量串聯重復,串聯重復和片段重復是基因家族擴張和進化的重要因素。系統發育分析也表明,許多藍莓AP2/ERF家族成員具有高度相似性,而一些VcERF基因的相鄰染色體位置為串聯復制提供了進一步的支持。
通過對ERF基因在藍莓休眠及解除過程中的轉錄水平進行研究,發現多數VcERF基因在內休眠及生態休眠解除過程中均能表達。其中VcERF51、VcERF115、VcERF119、VcERF74、VcERF95、VcERF14、VcERF29等基因在內休眠及生態休眠解除過程中表達量較高;VcERF116、VcERF118、VcERF123、VcERF82、VcERF81等基因隨著內休眠的解除表達量下降;不同的基因在不同暖溫處理時長下表達存在差異。啟動子順式作用元件分析結果顯示,10個ERF基因啟動子區域存在光響應、激素響應(脫落酸、赤霉素、生長素、茉莉酸甲酯、水楊酸)和脅迫響應元件(低溫、防御應激),說明藍莓ERF可能通過光、激素和脅迫信號調控藍莓花芽的生長發育和休眠。本研究可為藍莓ERF的研究提供一定的理論基礎。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
