紫薇屬植物葉綠體基因組研究進展
2022-12-02 07:54:28黃建睿繆紳裕
廣東農業科學 2022年10期
黃建睿,陳 濤,繆紳裕
(1.廣州大學生命科學學院,廣東 廣州 510006;2.深圳市中國科學院仙湖植物園,廣東 深圳 518004)
葉綠體是質體家族中的一種細胞器,含有豐富的葉綠素,是綠色植物進行能量轉化和光合作用的主要場所,賦予植物在地球生態環境中充當生產者的角色。此外,葉綠體是許多生化過程的基本場所,如氨基酸、核苷酸、脂肪酸、植物激素、維生素的合成以及硫和氮的同化[1]。葉綠體中具有半自主性的細胞器,自身擁有相對獨立的遺傳物質,即葉綠體基因組DNA(cpDNA),其結構一般為雙鏈環狀的DNA 分子,少數為線形,是僅次于核基因組的第二大基因組[2]。與核基因組相比,葉綠體基因組因其相對穩定的基因組結構、基因內容和基因序列,已被證明是用于遺傳多樣性評估的DNA 條形碼的重要數據來源,被廣泛應用于植物系統學研究[3]。葉綠體基因組按功能可分為遺傳系統基因、光合系統基因、合成系統基因、功能未知基因4類[4]。葉綠體基因組由于拷貝數高、母系遺傳、基因結構和排列保守的特點,是研究近緣物種系統發育關系的有力工具[5-7]。在被子植物中,葉綠體的進化速率也極其緩慢[8]。植物葉綠體基因組為系統發育、DNA 條形碼和種群間生物地理學提供了寶貴的資源。基因組學研究在基因組序列的遺傳組成、結構、組織、功能多樣性中,發揮了重要作用,尤其在系統發育研究中,能夠直觀地體現出植物間的進化關系[9-10]。
紫薇屬(Lagerstroemia)隸屬于千屈菜科(Lythraceae),全世界約有60 種紫薇屬植物,目前已培育出500 多個品種[11]。我國現有紫薇屬植物21 種,其中大花紫薇(Lagerstroemia speciosa)、南洋紫薇(L.siamica)、棱萼紫薇(L.turbinate)從東南亞引入[12]。紫薇屬植物多為落葉或常綠灌木或喬木,樹干多光滑,木材堅硬、耐腐,可作家具木材、建筑等使用,分布于亞洲東部、東南部、南部的熱帶、亞熱帶等地區[13]。紫薇(Lagerstroemia indicaL.)原產于中國,至少有1 500 年的種植歷史,隨后被引種到美國南部,采取雜交育種、誘變育種等多種培育方式,開始了紫薇屬植物在國外的育種歷程[14]。大多數紫薇屬植物具有大而美麗的圓錐花序,花期通常在夏季和秋季持續約3 個月或更久。此外,紫薇屬植物具有一定的藥用價值,根和枝葉入藥可用于治療過敏反應,具有止癢功效,而花和葉入藥有清熱解毒、利尿的效果,其葉子還可通過吸收煙霧和灰塵來凈化空氣,因而紫薇屬植物作為觀賞、藥用兼環保的優良花木,在園藝和園林應用中具有重要價值[15-17]。
本文以葉綠體基因組的研究現狀為背景,歸納紫薇屬植物葉綠體基因組的結構特征,總結了紫薇屬葉綠體基因組在DNA 條形碼、簡單重復序列以及系統發育中的應用,并分析了已完成序列測定的22 種植物的系統發育關系,旨在進一步歸納總結紫薇屬葉綠體基因組的研究現狀以及應用前景,為紫薇屬植物種質資源鑒定、分類和系統發育分析等方面的進一步研究提供理論鋪墊,同時為紫薇屬其他物種的葉綠體基因組研究以及物種進化和親緣關系分析等研究提供數據基礎。
1 葉綠體基因組研究概況
1.1 葉綠體全基因組序列測定
1986 年,植物葉綠體基因組的全序列測定最早在煙草(Nicotiana tabacum)中發表[18]。近年來,由于測序技術的不斷發展,測序成本逐漸降低,利用快速發展的第二代測序技術,越來越多物種的葉綠體基因組先后被測序用于系統發育研究,美國國家生物技術中心(The National Center for Biotechnology Information,NCBI)中關于葉綠體全基因組的數據不斷被增加充實。
最初,基因組測序主要使用以Sanger 測序為核心的測序技術,該法需要分離純化葉綠體基因組DNA 或者構建全基因組細菌人工染色體(Bacterial artificial chromosome,BAC)文庫,再利用含有物種葉綠體DNA 片段的載體進行測序,過程復雜且難度大,測序耗時長、成本高,因此未被大范圍使用。直到新一代高通量測序的出現,極大地擴大了測序通量,縮短了測序時長,為大規模葉綠體基因組測序提供了可能[19-21]。隨著測序成本的顯著降低,對整個葉綠體基因組進行測序變得更為便捷。目前,NCBI 數據庫已獲得超過900 個陸地植物完整的葉綠體基因組[22]。此外,在葉綠體全基因組中開發了眾多組裝軟 件,如 GetOrganelle[23]、Fast-Plast[24]、NOVOPlasty[25]、ORG.Asm[26]、chloroExtractor[27]、IOGA[28]、Chloroplast assembly protocol[29]等。
1.2 葉綠體基因組結構研究
葉綠體全基因組序列由于其相對穩定的基因組結構、基因內容和基因序列,已被廣泛接受為在分子進化方面有價值的數據來源。葉綠體基因組具有高度保守的環狀DNA 結構,少數為線形,是僅次于核基因組的第2 大基因組[30]。葉綠體基因組結構通常為高度保守的四分體結構,通常由1 個大單拷貝區(Large Single Copy,LSC)、1 個小單拷貝區(Small Single Copy,SSC)和2 個反向重復區(Inverted Repeats,IRs)組成。LSC區長約81~90 kbp,SSC 區范圍在18~20 kbp,2 個反向重復區大小介于20~30 kbp[31]。雖然葉綠體基因組的結構和大小高度保守,但IR/SC 邊界區域的擴張和收縮常被認為是造成高等植物葉綠體基因組長度差異的主要原因[32]。大多數被子植物的葉綠體基因組大小在107~218 kbp之間,由大約120 個基因組成,分別編碼rRNA、tRNA和蛋白質[33]。雖然葉綠體基因組結構高度保守,但也會發生基因缺失現象,且可能存在突變熱點[34-35],這為紫薇屬葉綠體基因組的相關研究奠定了基礎。
2 紫薇屬植物葉綠體基因組結構研究現狀
2.1 紫薇屬植物葉綠體基因組測序
隨著基因工程技術的發展,高通量技術的進步降低了測序成本,極大地促進了基因組和系統發育研究的進步。越來越多物種的葉綠體基因組序列被測序,這對紫薇屬植物葉綠體基因組的比較研究提供了一定的分子基礎,有助于提升對其葉綠體基因組應用價值的評價。目前,NCBI 顯示已完成22 種紫薇屬植物的葉綠體全基因組測序。表1 為目前已完成葉綠體全基因組測序的紫薇屬植物和5 個近緣屬的相關信息。
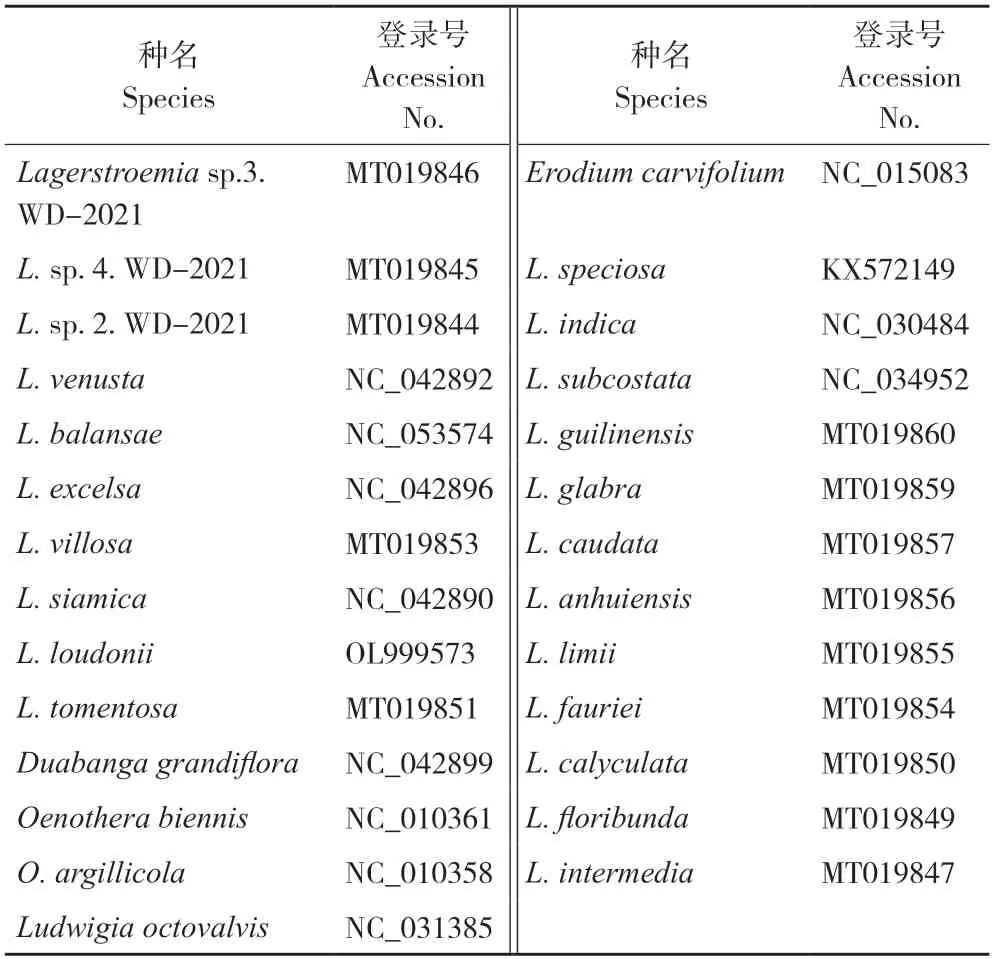
表1 來源于NCBI 的紫薇屬植物和外群葉綠體基因組序列Table 1 Chloroplast genome sequence of Lagerstroemia and outgroups derived from NCBI
2.2 紫薇屬植物葉綠體基因組結構特征
從NCBI 數據庫下載紫薇(L.indica)和絨毛紫薇(L.tomentosa)的葉綠體基因組序列,登錄號分別為NC_030484、MT019851,使用OGDRAW 在線軟件生成葉綠體基因組圖譜。由圖1 和圖2 可知,紫薇屬植物的葉綠體基因組呈閉合環狀結構,為高度保守的四分體結構,其葉綠體基因組通常也由4 部分組成,分別為1 個大單拷貝區(LSC)、1 個小單拷貝區(SSC)和2 個反向重復區(IRs),其中2 個反向重復區域的序列相同,但方向相反,通常命名為IRa、IRb,該區域會在紫薇屬葉綠體基因組進化過程中延伸或者縮小;LSC 和SSC 區的變異大于IR 區,非編碼區的分化程度大于編碼區[36]。由表2 可知,22種紫薇屬植物的葉綠體基因組大小約為150 kbp,最大的L.venusta長度為152 521 bp,最小的L.guilinensis長度為151 968 bp,其中LSC 區長度為83~84 kbp,SSC 區長度約為16 kbp,IR 區約為25 kbp,葉綠體基因組中嘌呤(GC)含量為37.6%~37.7%。紫薇屬植物的葉綠體基因組的基因數量大多為112 個,其中包括78 個蛋白編碼基因、30 個tRNA 基因、4 個rRNA 基因,但L.balansae的葉綠體基因組含有130 個基因,包括85 個蛋白編碼基因、37 個tRNA 基因、8 個rRNA 基因。葉綠體基因組雖然在基因結構上高度保守,但IR/SC 邊界區域的擴張和收縮引起的IR/SC 連接位置的變化,通常被認為是造成高等植物葉綠體基因組長度變異的主要機制[37-38]。Zheng等[39]發現13 種紫薇在IR/SC 交界區表現出相似的特征,而Xu等[40]在3 個新測序的紫薇葉綠體基因組中觀察到rpl2 內含子缺失,紫薇rpl2 內含子缺失的發生被認為是千屈菜科中重要的進化事件之一。
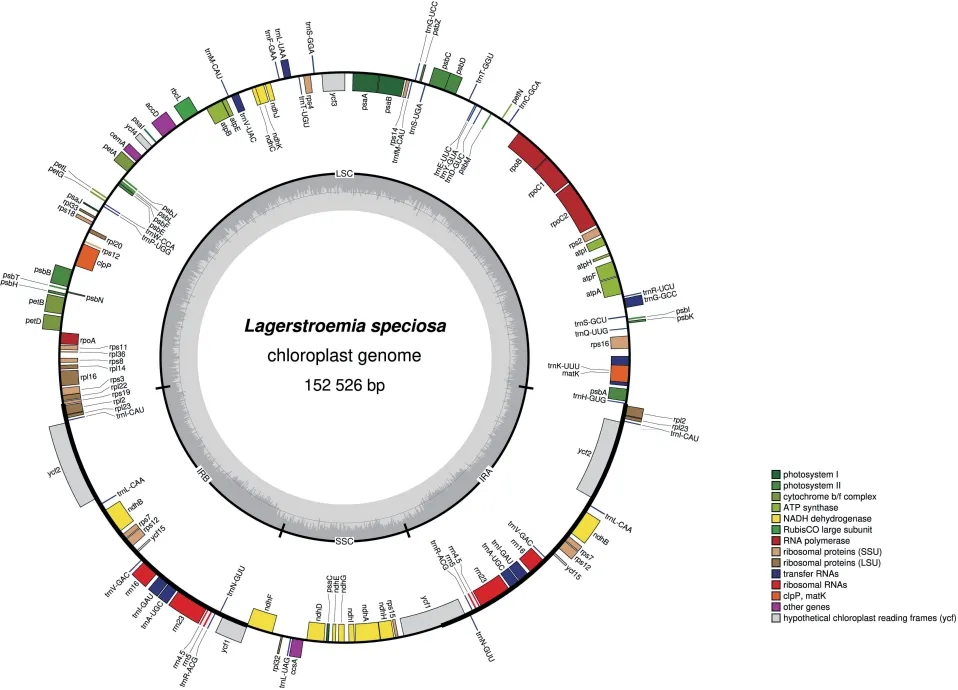
圖1 大花紫薇葉綠體基因組物理圖譜Fig.1 Gene map of the choloroplast genome of Lagerstroemia speciosa
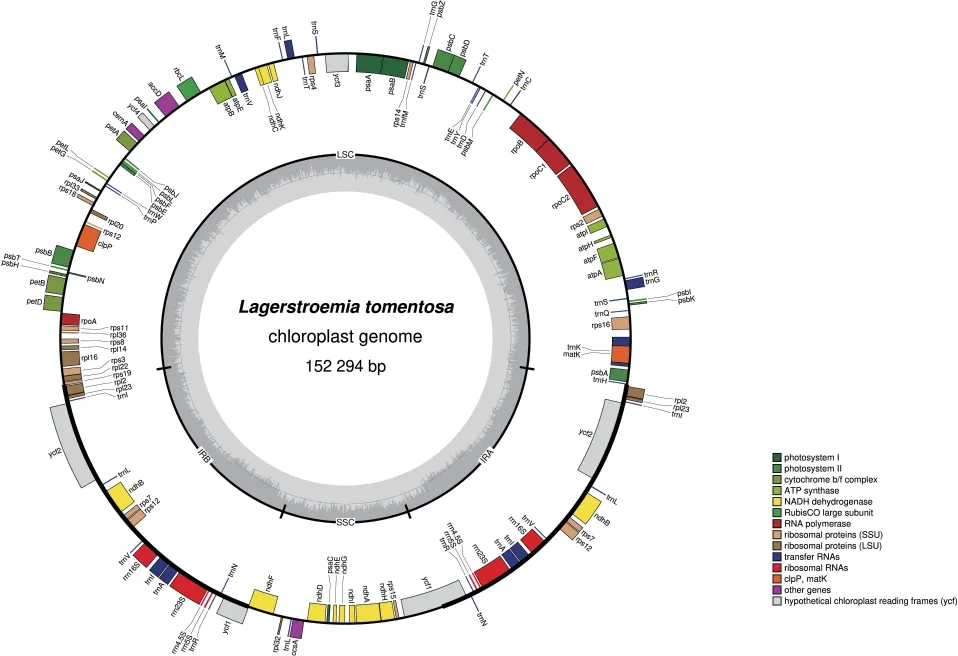
圖2 絨毛紫薇葉綠體基因組物理圖譜Fig.2 Gene map of the choloroplast genome of Lagerstroemia tomentosa
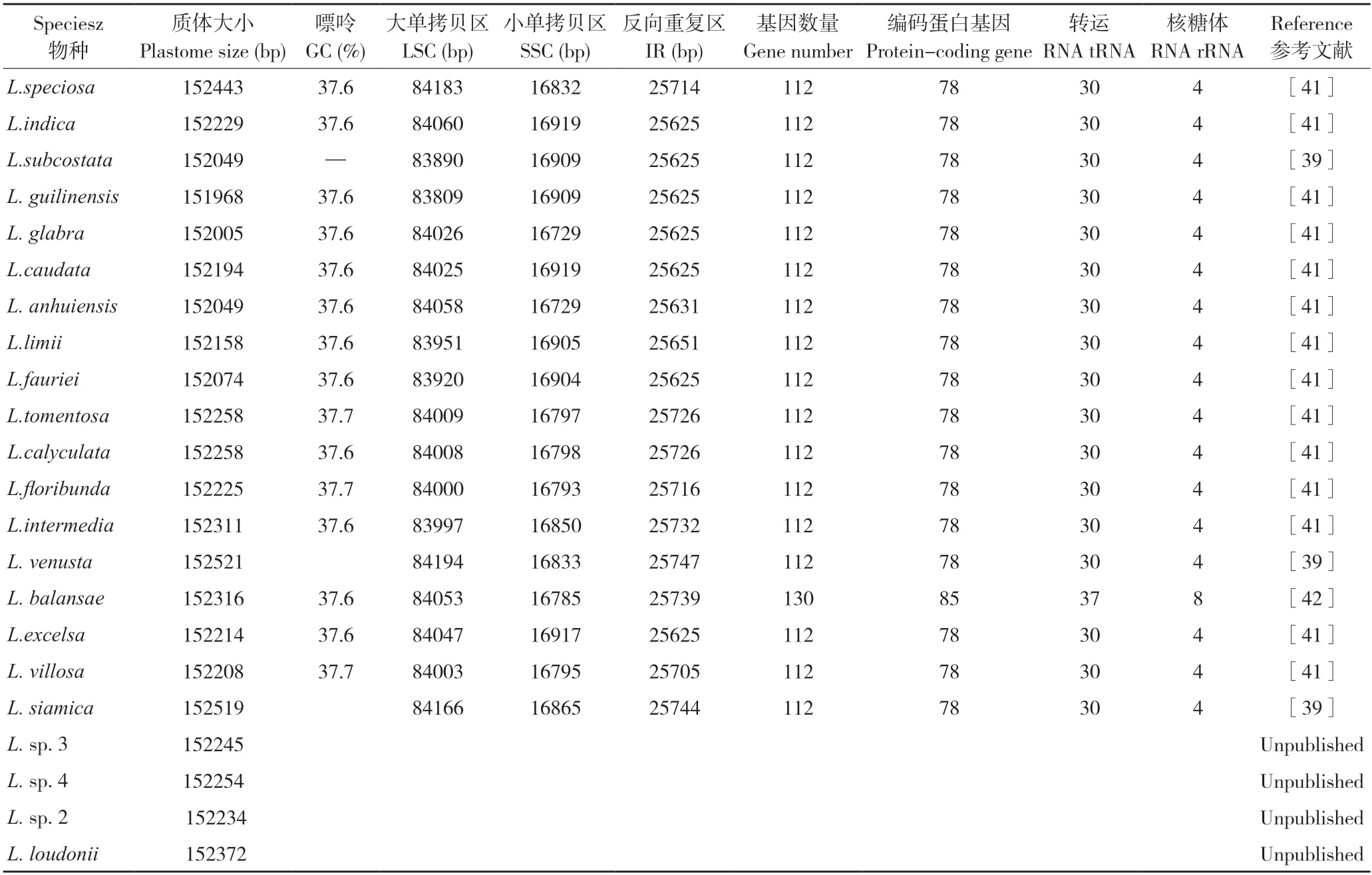
表2 紫薇屬植物葉綠體基因組序列特征匯總Table 2 Summary of chloroplast genome sequence characteristics of Lagerstroemia
2.3 22 種紫薇屬植物的系統發育關系
為了確定紫薇屬植物的系統發育關系,根據表1 中由NCBI 下載的FASTA 序列,使用軟件PhyloSuite,采用貝葉斯法(Bayesian inference,BI)構建22 種紫薇屬植物的系統發育樹,另設置5 個近源外群Duabanga grandif lora、Oenothera biennis、O.argillicola、Ludwigia octovalvis、Erodium carvifolium。由圖3 可知,L.calyculata、L.loudonii、L.sp.2、L.tomentosa、L.sp.3、L.floribunda、L.balansae、L.intermedia、L.siamica、L.speciosa、L.venusta、L.anhuiensis、L.glabra、L.caudata、L.excelsa、L.indica、L.guilinensis、L.sp.4、L.limii、L.subcostata、L.fauriei、L.villosa共22 種紫薇屬植物單獨為一支,為單系群,具有較大的支持率。

圖3 基于27 個物種的葉綠體全基因組用貝葉斯法(Bayes)構建的系統發育樹Fig.3 Phylogenetic tree based on the 27 species chloroplast genome sequences with Bayes’method
3 紫薇屬植物葉綠體基因組的應用研究進展
3.1 DNA 條形碼研究
DNA 條形碼(DNA barcode)是指生物體內能夠代表該物種的、標準的、有足夠變異的、易擴增且相對較短的DNA 片段。該技術是利用生物體DNA 中一個或幾個保守片段對物種進行快速準確鑒定的新興生物技術。葉綠體上的DNA序列片段(如matK、rbcL、trnH-psbA、rpoC1、rpoB、accD、ycf5 等)在植物DNA 條形碼被廣泛應用。Xu等[40]從6 種紫薇屬植物的葉綠體基因組中選擇12 個相對較高的變異區(trnK-rps 16、trnStrnG、trnG-trnR-atpa、trnE-trnT、rbcLaccd、psbL-psbF-psbE、trnP-psaJ-rpl33、rrn16-trni、ccsa、ndhG-ndhI、rps15-ycf1 和ycf1)作 為cp DNA 標記,推測它們在物種和品種水平上經歷了更快的核苷酸替換,可以作為分子標記應用于紫薇屬植物的系統發育分析和植物鑒定;Zheng等[39]從13 種紫薇屬植物中選擇識別度最高的7 個基因片段(ndhF、ycf1、trnK-rps16、psbKPSBI、trnR-ucu-atpa、rpl32-trnL 和rrn16-trni)作為DNA 條形碼,結果發現這7 個條形碼能有效鑒別13 個種。Dong等[41]比較了20 種紫薇屬植物中的4 個高變量標記和國際DNA 通用條形碼(rbcL、matK、psbA-trnH),發現4 個高變量標記的進化速率是DNA通用條形碼的2.5倍。馬麗[43]針對國際DNA 條形碼對紫薇屬植物的鑒定進行研究,結果發現trnH-psbA 在13 個紫薇屬物種中變異最大,但是鑒定成功率僅為38.46%;MatK 和rbcL 變異太小不適合用作紫薇屬DNA 條形碼;而組合片段rbcL+trnH-psbA 和mat K+rbc L+trn H-psb A 鑒定能力高,可用作紫薇屬特異性條形碼;構建NJ 樹對15 個高變片段進行評估,發現其 中5 個片段 petA-psbJ、ndh F-rpl32、ndhGndhI、trnS-trnG 和trnR-atpA 的鑒定能力最強,因此建議將它們作為紫薇屬的候選DNA 條形碼。前人從紫薇屬葉綠體基因組中開發的DNA 標記能彌補國際通用條形碼在紫薇屬中的鑒定缺陷,對紫薇種質資源的種間鑒定、定向育種以及系統發育關系提供了強有力的分子手段。
3.2 SSR 研究
簡單重復序列(Simple sequence repeat,SSR)是由1~6 個核苷酸組成的簡單重復的串聯序列,SSR 在基因組的不同位置不同分布,長度一般在200 bp 以下,通常有6 種核苷酸類型,即單核苷酸、二核苷酸、三核苷酸、四核苷酸、五核苷酸以及六核苷酸,且SSR 在真核和原核生物中廣泛分布[44-45]。SSR 分子標記具有在基因中覆蓋率高、重復次數多、多態性豐富、穩定性好等優點,能夠檢測出品種親緣關系之間的細小差異,現已經廣泛應用于品種間親緣關系及遺傳多樣性的研究[46]。
隨著生物學技術的發展,分子標記已成為紫薇屬植物在遺傳多樣性及親緣關系分析等研究的重要手段。例如,王獻等[47]設計并優化了20 個紫薇和南紫薇的AFLP 銀染色反應系統,該技術還應用于分析30 個紫薇品種和2 個近緣種的親緣關系;顧翠花[48]在王獻等[47]的基礎上針對紫薇建立AFLP 實驗反應體系,篩選出可用于分析紫薇種質資源的引物,并用于評價13 個紫薇群體的親緣關系;徐靜靜等[49]利用ISSR 技術,基于4個紫薇群體和48 個不同花色的紫薇品種,分析了紫薇屬品種的花色遺傳多樣性;Wang等[50]利用78 個SSR 標記分析評價了51 個紫薇品種和5 個屋久島紫薇品種的遺傳多樣性,驗證了現有品種與已鑒定但未開發種質資源的品種之間的關系。這些研究在紫薇品種鑒定、分類、遺傳多樣性評價等方面取得了一定進展。
葉綠體基因組中的簡單重復序列在種內水平上可能是高度可變的,因此經常被用作群體遺傳學和進化研究中的遺傳標記。Gu等[36]分析了22 種千屈菜科植物的SSR 位點,其中包含14種紫薇屬植物,研究發現每個種均有211~332 個SSR,長度包含8~16 個堿基,共發現單核苷酸、二核苷酸、三核苷酸、四核苷酸、五核苷酸和六核苷酸6 種SSR,以單核苷酸的重復序列最常見,數量在123~212 個,其中,紫薇屬植物中只有L.siamica和L.intermedia 存在六核苷酸。SSR 位于31 個編碼基因和57 個基因間隔區中,結果表明葉綠體基因組變異較大的SSR 可用于近緣物種的鑒定和系統發育研究[35]。Xu等[40]分析了6 種紫薇屬植物的葉綠體基因組中的SSR,發現SSR 的長度在10~15 個堿基之間,對6 個紫薇基因組序列的比較分析表明,共檢測到5 類SSR(單核苷酸、二核苷酸、三核苷酸、四核苷酸和五核苷酸)重復。Zheng等[39]通過比較13 種紫薇屬植物的葉綠體基因組中SSR 的分布和數量,發現了從單核苷酸到六核苷酸的SSR,前5 種SSR 分別存在于紫薇屬13 種植物中,六核苷酸重復序列也僅存在于Lagerstroemia siamica和L.intermedia的葉綠體基因組中,發現SSR 序列分布在31 個基因編碼區和57 個基因間隔區,這與Gu 在22種千屈菜科的14 種紫薇屬植物的SSR 分布研究結果一致,SSR 在葉綠體基因組中分布不均的現象特征,有助于SSR 分子標記技術應用到種下層面的系統發育分析。馬麗[43]研究了13 種紫薇的66 個SSR 位點,選擇了7 個SSR 數量最高的基因,發現同源 SSR 沒有顯著差異,這也從側面說明了SSR 的多態性。在紫薇屬物種中鑒定SSR 位點為多態性SSR,為紫薇屬SSR 標記的開發提供參考序列。
3.3 系統發育關系研究
系統發育也稱為系統發展,它是指某一個類群的形成和發展過程。通過建立系統發育樹,能更直觀地分析類群的親緣關系。Xu等[40]使用最大簡約(MP)、最大似然(ML)和貝葉斯推斷(BI)方法,基于4 個葉綠體基因組全序列、編碼區、非編碼區和12 個高變區,在高Bootstrap 支持下完全區分了所有6 個紫薇屬分類群,獲得了較大的支持率。Zheng等[39]基于32 種物種(其中包括13 種紫薇)的66 個共享蛋白質編碼基因構建的系統發育樹,揭示了紫薇屬內物種的系統發育關系及其在桃金娘目中的系 統發育位置。紫薇屬植物的系統發育,包括更具代表性的物種和大量的分子標記,對于了解紫薇屬植物的進化史、新品種的選育和紫薇種質資源的保護至關重要[50]。
4 結語與展望
紫薇屬植物具有花期長、花色艷麗且抗污能力強的特點,是我國夏季重要的園林觀賞植物。本文在分析葉綠體基因組各 結構中,由于L.loudonii、L.sp.2 WD-2021、L.sp.3 WD-2021、L.sp.4 WD-2021 的參考文獻未發布,暫只對19種紫薇屬植物的葉綠體基因組結構進行概述,且已進行葉綠體基因組測序的物種僅占已有物種的1/3,因此對紫薇屬植物葉綠體基因組結構的概括不夠系統。進一步完成未進行葉綠體基因序列的測定,有利于紫薇屬內親緣關系以及屬內基因組的比較研究。
相比利用基因片段作為DNA 標記,簡短的片段無法準確評估物種在系統發育樹中的位置,因此亟需通過完整的葉綠體基因組序列判定物種親緣關系,利用完整的葉綠體基因組序列進行基因組比較研究更具有說服力[51-53]。新測序的紫薇屬植物葉綠體全基因組將利于提高對紫薇屬植物葉綠體基因組的認識,并有助于對該物種開展資源保護工作。通過比較完整的葉綠體基因組,提高對葉綠體基因組進化、物種鑒定和系統發育關系的理解,同時有助于提升對葉綠體基因組應用價值的評價,為紫薇屬植物后續的種質資源鑒定、分類和系統發育分析等方面的進一步研究提供理論鋪墊。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
