定向進化肌醇加氧酶MIOX4提高葡萄糖二酸的產量
2022-11-29 04:14:26蘇蕊芳李詩韻毛銀趙運英鄧禹
食品與發酵工業 2022年22期
蘇蕊芳,李詩韻,毛銀,趙運英*,鄧禹,*
1(江南大學, 糧食發酵與食品生物制造國家工程研究中心,江蘇 無錫,214122)2(江南大學 生物工程學院,江蘇 無錫,214122) 3(江南大學,江蘇省生物活性產品加工工程研究中心,江蘇 無錫,214122)
D-葡萄糖二酸(glucaric acid, GA)是葡萄糖的衍生物,是一種天然的二元酸,存在于許多水果和蔬菜中,特別是在十字花科蔬菜、西柚、柑橘等植物中葡萄糖二酸的含量較為豐富。葡萄糖二酸不僅在醫藥中有抗癌、治療糖尿病的作用[1],而且因其能合成羥基化尼龍,在紡織服裝方面也有廣泛的應用[2]。因此,它在2004年被美國能源部確認為“最具價值的生物煉制產品”之一[3]。
目前,葡萄糖二酸主要通過葡萄糖的化學氧化方法合成,該方法以硝酸為溶劑和氧化劑[4]。但化學氧化法生產葡萄糖二酸存在收率低、副產物多、環境不友好等缺點。而采用生物法合成葡萄糖二酸對環境更加友好,并且能夠實現低成本生產葡萄糖二酸。生物法生產葡萄糖二酸有很多種途徑,現階段,葡萄糖二酸的生物生產方法主要以微生物發酵為主。PRATHER團隊通過共同表達釀酒酵母(Saccharomycescerevisiae)的肌醇-1-磷酸合成酶(Ino1)、小家鼠的肌醇加氧酶(MIOX)和丁香假單胞菌(Pseudomonassyringae)的糖醛酸脫氫酶(Udh),構建了利用大腸桿菌從葡萄糖生物法合成葡萄糖二酸的途徑[5]。由于酵母的耐酸性和較強的抗噬菌體能力,GUPTA等[6]將大腸桿菌的葡萄糖二酸合成途徑轉移到釀酒酵母中,在外加肌醇的條件下葡萄糖二酸的最大滴度達到1.6 g/L。此外,釀酒酵母可以通過自身的Ino1和肌醇單磷酸酶(Inm1/2)轉化葡萄糖-6-磷酸生成肌醇,因此,研究人員只需要在釀酒酵母中表達外源的miox基因和udh基因即可實現葡萄糖到葡萄糖二酸的轉化[6-8]。
在研究葡萄糖二酸代謝途徑過程中,發現MIOX的酶活力比Udh低2個數量級[5],并且穩定性較差,容易失活。因此MIOX的催化過程成為葡萄糖二酸合成的限速步驟。研究人員采取了一系列代謝工程策略提高葡萄糖二酸的產量,這些策略包括蛋白質融合標簽[9]、定向進化、構建多肽支架[10]等。本實驗室通過將擬南芥(Arabidopsisthaliana)的miox4基因和丁香假單胞菌的udh基因構建到釀酒酵母的delta位點上提高了葡萄糖二酸的產量[11];之后利用linker將MIOX4和Udh進行融合,在釀酒酵母的delta位點表達[12],使葡萄糖二酸的產量大大提高;這些方法均提高了葡萄糖二酸的產量,但MIOX4的活力及穩定性依然無法滿足應用要求,因此采用定向進化的方法篩選出酶活力較高的MIOX4具有重要意義。
定向進化是利用基因工程技術模擬自然界中的進化,建立蛋白質突變庫,從中篩出具有優良特性的蛋白質突變體[13]。研究人員通常采用易錯PCR技術對酶進行定向進化[14-15]。葡萄糖二酸經跨膜蛋白轉運進入胞內后激活CdaR[16]蛋白使其轉變為活性形式,進而與操縱子區域結合,啟動下游結構基因轉錄。基于上述原理,王毳等[17]在大腸桿菌中構建了葡萄糖二酸生物傳感系統篩,并將其應用到篩選來自小鼠的肌醇加氧酶突變株的研究過程。本研究采取兩輪易錯PCR的方法提高肌醇加氧酶的活性,通過葡萄糖二酸的產量篩選正突變菌株。同時,利用本實驗構建的大腸桿菌葡萄糖二酸傳感器質粒R7M10[18]作為高通量篩選工具篩選MIOX4突變株。
本研究以釀酒酵母BY4741opi1Δ(敲除OPI1基因的BY4741菌株)[11]為出發菌株,利用酵母自身的Ino1將葡萄糖-6-磷酸轉化為肌醇-1-磷酸,再經過酵母的Inm1/2將肌醇-1-磷酸轉化成肌醇,肌醇經過外源的MIOX4轉化為D-葡萄糖醛酸,之后再經過外源Udh的催化生成葡萄糖二酸。因此本實驗通過將擬南芥來源的肌醇加氧酶miox4基因和丁香假單胞菌來源的醛酸脫氫酶udh基因整合到BY4741opi1Δ的基因組上,以此構建葡萄糖二酸生物合成途徑。本研究對擬南芥來源的MIOX4進行定向進化,通過兩輪易錯PCR構建MIOX4突變體文庫,并利用大腸桿菌葡萄糖二酸傳感器作為高通量篩選工具對MIOX4突變體文庫進行初篩,然后對篩選到的陽性轉化子進行搖瓶發酵,挑選正向突變體并確定發生有益突變的氨基酸位點(圖1)。
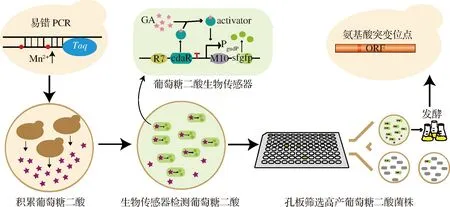
圖1 MIOX4突變體文庫構建和篩選策略示意圖Fig.1 The schematic diagram of the construction and screening strategy for MIOX4 mutant library
1 材料與方法
1.1 材料
1.1.1 菌株和質粒
本實驗所用菌株和質粒詳見表1。
1.1.2 培養基
釀酒酵母用YPD[(g/L)(酵母提取物10、蛋白胨20、葡萄糖20)]、SD-URA、SD-HIS[(g/L)(葡萄糖20,無氨基酵母氮源1.7,硫酸銨5.0,10×必需氨基酸混合液,無對應的氨基酸URA/HIS)]培養基進行培養,發酵培養基在YPD培養基的基礎上外加60 mmol/L肌醇;大腸桿菌用LB[(g/L)(氯化鈉10,酵母提取物5,蛋白胨10)]培養基進行培養,加入相應的抗生素用于篩選轉化子;檢測熒光的大腸桿菌用M9[(g/L)(磷酸氫二鈉6.78,磷酸二氫鉀3,氯化鈉0.5,氯化銨1,硫酸鎂0.24,葡萄糖4,氯化鈣0.1 mmol/L)]培養。以上培養基添加2.0%(質量分數)瓊脂粉配制固體培養基。

表1 本研究所用菌株和質粒Table1 Strains and plasmids used in this study
1.1.3 試劑和儀器
2×TaqPCR Master Mix,南京諾唯贊科技有限公司;PCR引物(表2)及基因序列測序,蘇州金唯智生物有限公司;肌醇、葡萄糖二酸標準品,Sigma公司;其他常規試劑采用國產分析純或進口分裝。
ETC811基因擴增儀,德國Eppendorf公司;Gel Doc XR+凝膠成像儀,美國Bio-Rad公司;SYNERGY H1多功能酶標儀,南京拜爾克智能科技有限公司;UV-1800分光光度計,上海翱藝儀器有限公司;1260Ⅱ高效液相色譜,美國安捷倫科技有限公司。

表2 本研究所用引物序列Table 2 Primer sequence used in this study
1.2 實驗方法
1.2.1 MIOX4蛋白活性位點的預測及三級結構模擬
在ZhangLab[19]https://zhanggroup.org/COACH/網站上預測來自擬南芥的MIOX4蛋白的配體結合位點和催化活性位點;利用Swiss-Modelhttps://swissmodel.expasy.org/在線軟件對MIOX4進行同源建模;使用可視化工具PyMOL對MIOX4進行結構模擬。
1.2.2 基因的擴增
以菌株Bga-3[11]的基因組為模板,利用引物對HIS-udh-F/R擴增帶有同源臂的HIS-udh,將該片段轉化到釀酒酵母BY4741opi1Δ,涂布到SD-HIS的平板上,挑取轉化子提基因組后利用引物對check-udh- F/R對基因組進行PCR驗證,驗證條帶正確的轉化子表示udh整合到OPI1的終止子上,即為BY4741opi1Δ-udh。
以質粒pY26-miox4-udh[11]為模板,利用引物對URA3-F/R擴增帶有同源臂的URA3,利用引物mi- ox4-1-F和miox4-4-R擴增miox4,將擴增得到的片段URA3和miox4利用引物URA3-F和miox4-4-R進行融合得到URA3-miox4,將該片段轉化到BY4741opi1Δ-udh菌株中,涂布到SD-URA的平板上,對轉化子提基因組驗證,驗證條帶正確的轉化子表示miox4整合到OPI1的啟動子上,得到未突變的對照菌株BY4741opi1Δ-udh-miox4。
1.2.3 易錯PCR構建突變體文庫
將miox4分成4段,分別標記為1、2、3、4,miox4-1片段中包含miox4的啟動子和ORF的上游,miox4-2片段為易錯片段1,miox4-3片段為易錯片段2,miox4-4片段中包含miox4的終止子和ORF的下游。以質粒pY26-miox4-udh為模板,利用引物對miox4-1-F/R擴增帶有同源臂的miox4-1片段,引物miox4-3-F和miox4-4-R擴增miox4-3-4(包含3和4片段)片段,引物miox4-1-F和miox4-2-R擴增miox4-1-2(包含1和2片段),使用引物對miox4-2-F/R和miox4-3-F/R對miox4-2和miox4-3分別進行易錯PCR;將擴增得到的miox4-1、易錯片段miox4-2*、miox4-3-4進行融合,片段miox4-1-2、易錯片段miox4-3、miox4-4進行融合PCR,均得到易錯miox4*;將易錯miox4*和URA3片段利用引物miox4-1-F和URA3-R進行融合PCR得到miox4*-URA3,將該片段轉化到BY4741opi1Δ-udh菌株中,涂布到SD-URA的平板上,所得到的轉化子即為突變體文庫。
1.2.4 利用大腸桿菌葡萄糖二酸傳感器初篩MI-OX4的正向突變體
將突變體文庫菌株接種到裝有2 mL YPD培養基的24孔板中,30 ℃培養12 h作為種子液,再轉接到裝有10 mL YPD(外加60 mmol/L肌醇)的試管中進行發酵培養,在24、48 h分別補5 g/L的葡萄糖,發酵培養7 d后,12 000 r/min轉速離心5 min收集發酵液。
將含有質粒R7M10的大腸桿菌BL21種子以2%的接種量轉接到M9培養基中,接種之后的混合物與發酵液按1∶1加入到96深孔板中,培養至OD600值為0.6~0.8,用酶標儀BioTek HT檢測熒光(激發485 nm,發射528 nm)和OD600值,取熒光/OD600的值,篩選熒光比值比對照高的突變菌株。
1.2.5 突變菌株的搖瓶發酵
將突變菌株接種到250 mL的搖瓶中發酵培養,30 ℃、250 r/min的條件下培養,在細胞生長對數期24、48 h分別補加5 g/L的葡萄糖,每隔24 h取樣檢測OD600值及葡萄糖二酸的濃度。
1.2.6 葡萄糖二酸的測定
標準樣品的準備:稱取一定質量的葡萄糖二酸鉀標樣,融于超純水中,用KOH溶液調pH至中性,定容到終質量濃度為1 g/L的葡萄糖二酸溶液。
樣品處理:取發酵溶液12 000 r/min轉速離心5 min,收集上清液,用10 mmol/L稀硫酸稀釋2倍,再12 000 r/min轉速離心5 min混勻,經0.22 μm濾膜過濾,供液相分析。
高效液相色譜(high-performance liquid chromatography,HPLC)分析條件:流動相為5 mmol/L稀硫酸,色譜柱為HPX-87H,進樣量20 μL,流速0.6 mL/min,柱溫35 ℃,檢測器為示差檢測器。
2 結果與分析
2.1 易錯PCR片段的確定
由于MIOX4的編碼區序列較長,直接易錯PCR效率較低,因此為了提高MIOX4的突變概率,本研究在ZhangLab網站上預測MIOX4蛋白的配體結合位點和催化活性位點,利用Swiss-Model在線軟件對MIOX4進行同源建模,使用可視化工具PyMOL對MIOX4進行結構模擬。通過模擬預測MIOX4的三維結構,發現其結合位點和活性位點在2個鐵原子和底物的周圍(圖2-A)。預測結果顯示,有13個配體結合位點,8個催化活性位點,配體結合位點分別是Arg59、Tyr75、Asp116、Ser118、Asp119、Asp155、Lys158、Gly175、Asp176、His227、His253、Ser254、Tyr256(藍色和紫色);催化活性位點分別是His129、His154、Asp155、Lys158、His227、His253、Ser254、Asp286(粉色和紫色);其中Asp155、Lys158、His227、His253、Ser254既是催化活性位點,又是配體結合位點(紫色)(圖2-B)。預測結果顯示這些氨基酸位點大多集中在MIOX4編碼區序列的中間區域,對MIOX4編碼區的中間區域分成兩段進行易錯PCR,以此提高突變概率。
2.2 葡萄糖二酸合成途徑的構建
為了方便對突變MIOX4進行篩選,本研究在釀酒酵母BY4741opi1Δ菌株上構建葡萄糖二酸的代謝途徑,葡萄糖二酸由肌醇加氧酶MIOX4和糖醛酸脫氫酶Udh催化肌醇產生葡萄糖二酸,因此通過葡萄糖二酸的產量來篩選MIOX4突變株。以BY4741opi1Δ為出發菌株,在該菌株的基因組上構建葡萄糖二酸的代謝途徑。首先將udh構建到BY4741opi1Δ菌株OPI1的終止子上,構建基礎菌株BY4741opi1Δ-udh;然后將經過隨機突變的miox4*構建到該菌株OPI1的啟動子上得到突變體菌株BY4741opi1Δ-udh-miox4*,作為突變體文庫,未發生突變的miox4作為對照菌株BY4741opi1Δ-udh-miox4(圖3)。以菌株Bga-3基因組為模板擴增3 330 bp的HIS-udh(圖4-A),并轉化BY4741opi1Δ,正確的BY4741opi1Δ-udh轉化子經過驗證能擴增915 bp(圖4-B)。
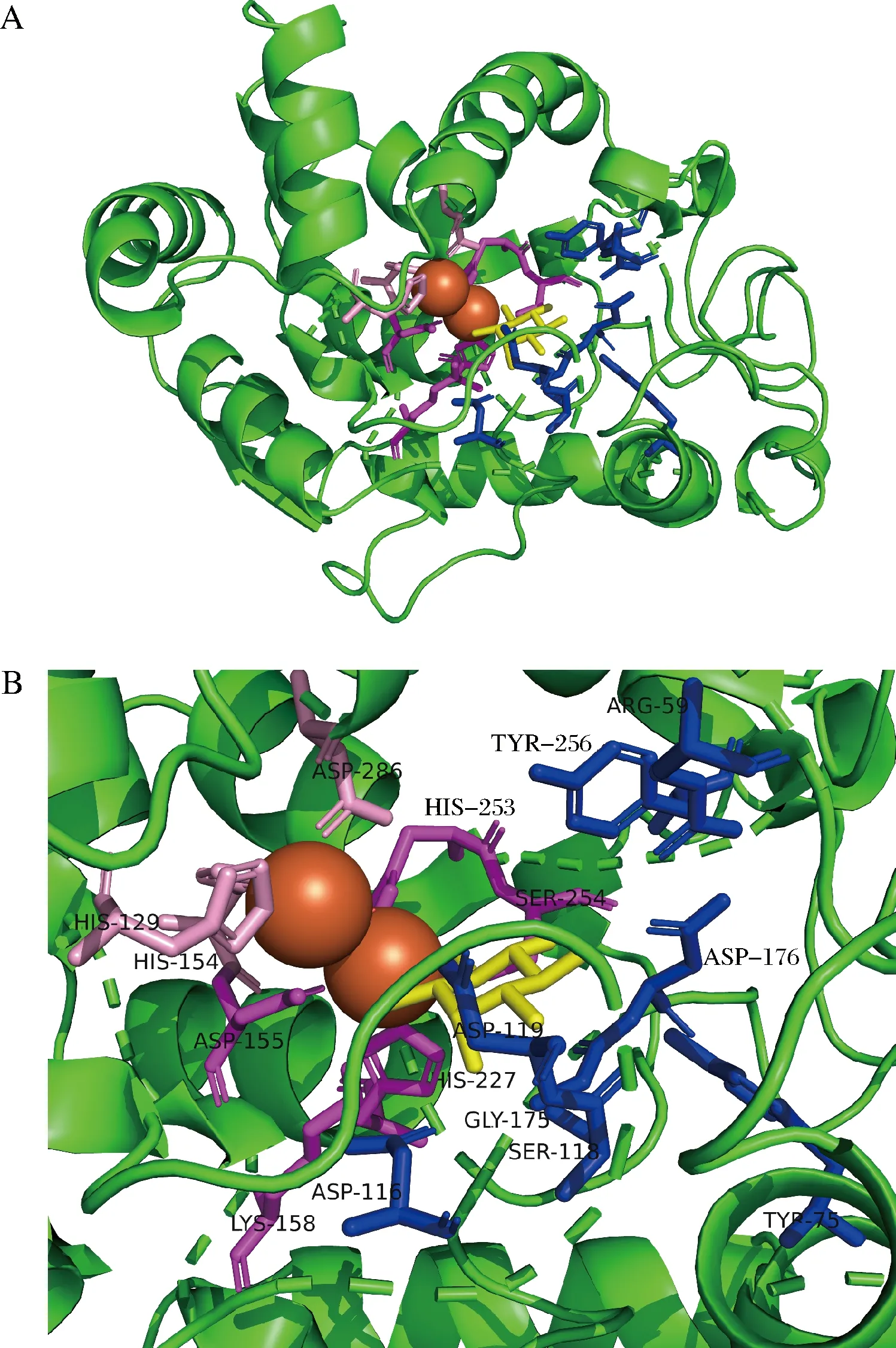
A-模擬MIOX4的三維結構;B-預測MIOX4的催化活性位點和配體結合位點圖2 MIOX4蛋白的三維結構模擬圖Fig.2 A simulation diagram of the three-dimensional structure of the MIOX4 protein注:橙色球體為鐵原子,黃色為底物肌醇,藍色和紫色為配體結合位點,粉色和紫色為催化活性位點,紫色為配體結合位點和催化活性位點
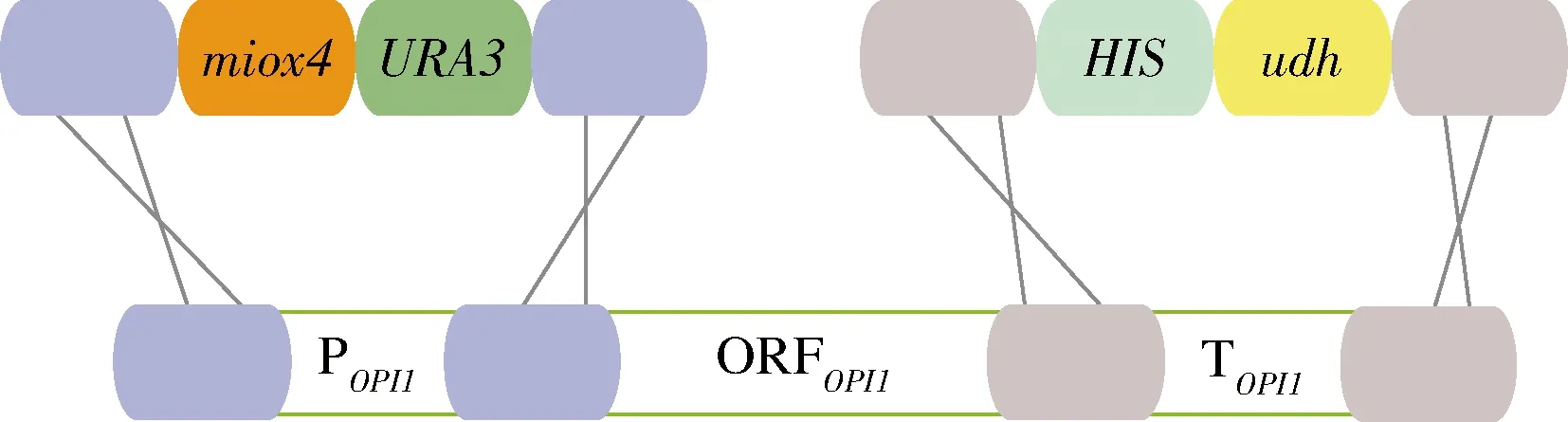
圖3 構建葡萄糖二酸代謝途徑的示意圖Fig.3 Schematic diagram of constructing glucaric acid metabolic pathway
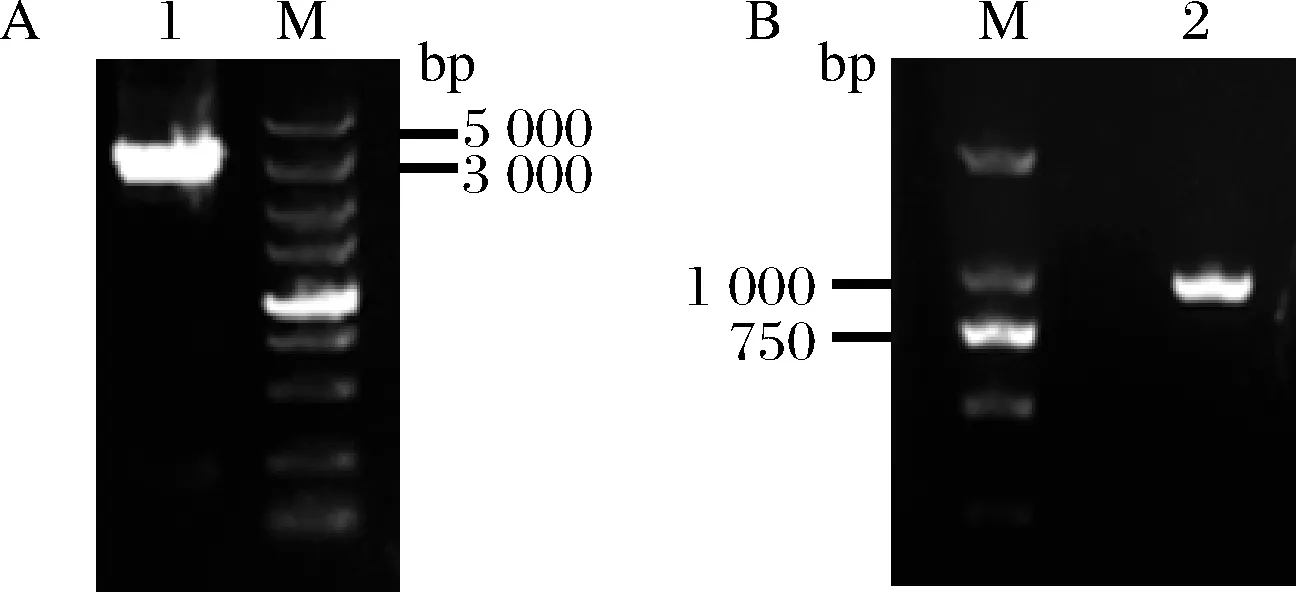
A-擴增HIS-udh;B-轉化子驗證結果圖4 BY4741opi1Δ上構建udh 基因Fig.4 Construction of udh gene in BY4741opi1Δ
2.3 易錯PCR對MIOX4定向進化
通過對MIOX4蛋白結構的預測,MIOX4蛋白的配體結合位點和催化活性位點集中在編碼區的中間區域,為了提高突變率,將miox4的編碼區的中間區域分成2段分別進行易錯PCR。這樣就有2種情況,一種情況是miox4-2發生突變,miox4-2序列包括39位的天冬氨酸到205位的酪氨酸(圖5-A),另一種情況是miox4-3發生突變,miox4-3序列包括196位的谷氨酸到298位的谷氨酸(圖5-B),以此擴大突變文庫。
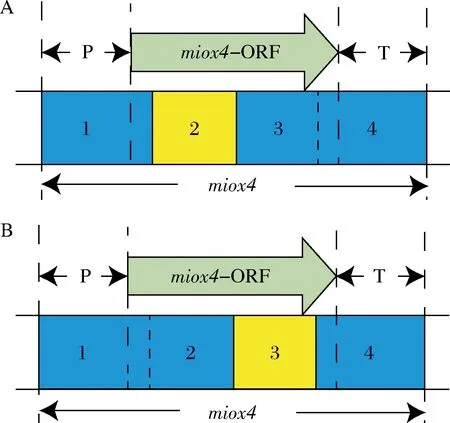
A-對miox4-2片段易錯;B-對miox4-3片段易錯圖5 兩種易錯PCR的情況示意圖Fig.5 Schematic diagram of two error-prone PCR scenarios
Mn2+可以通過降低聚合酶對模板的特異性而增加堿基錯配概率,研究人員一般采用外加Mn2+的方式進行易錯PCR[20-21]。我們亦采用外加不同濃度的Mn2+進行易錯PCR,為了提高突變率,本研究采用兩輪易錯PCR的方式來進行。以質粒pY26-miox4-udh為模板,選取濃度為1、1.5、2、2.5、3、3.5、4、4.5 mmol/L的Mn2+分別對miox4-2和miox4-3進行第一輪易錯PCR,由圖6-A可知,隨著Mn2+濃度升高,條帶逐漸變淡,選取Mn2+濃度為3.5、4、4.5 mmol/L的PCR產物作為模板對其進行第二輪易錯PCR,Mn2+濃度為1、1.5、2、2.5、3、3.5、4、4.5 mmol/L,隨著Mn2+濃度升高,條帶逐漸變淡并且消失(圖6-B)。
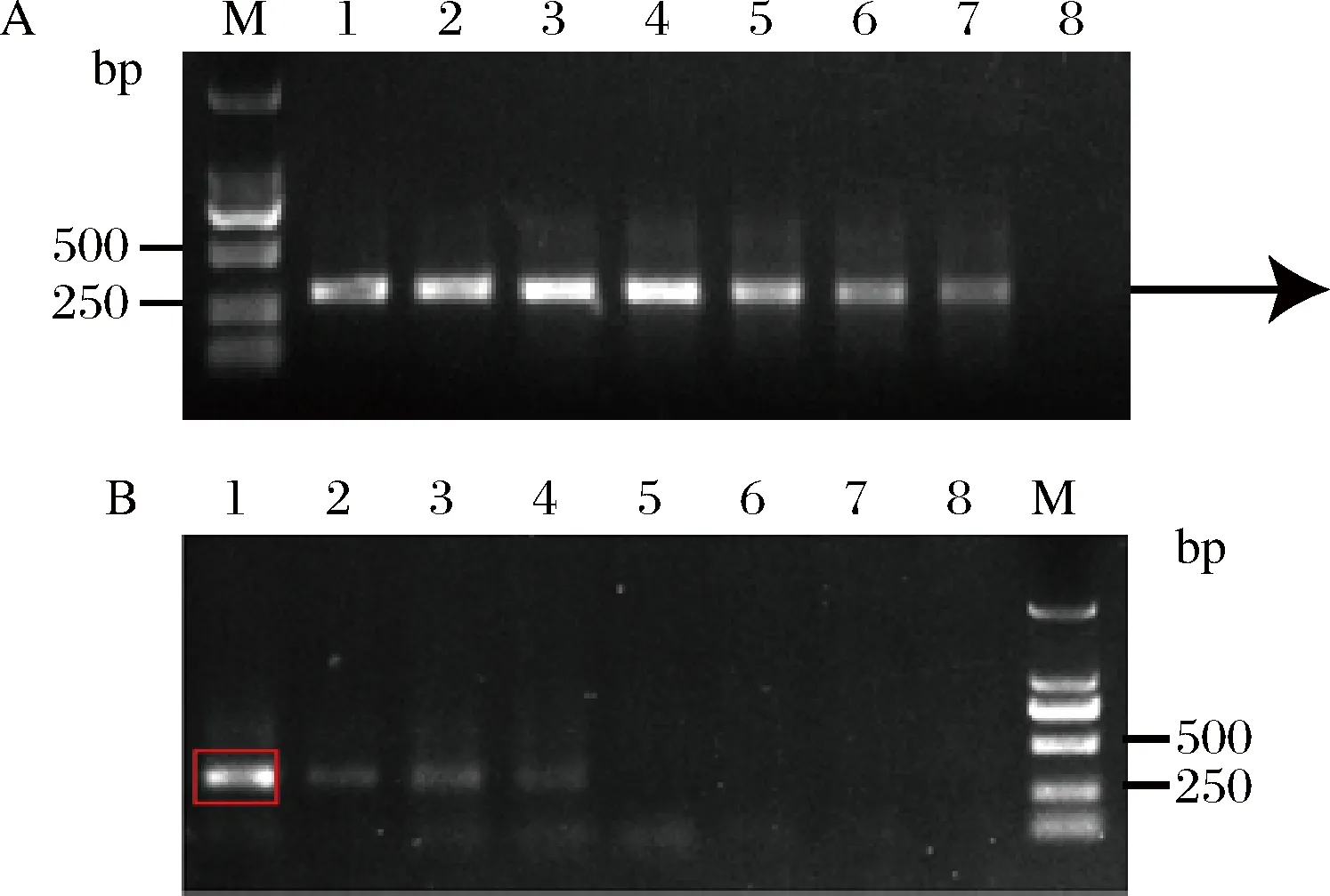
1~8-Mn2+濃度1、1.5、2、2.5、3、3.5、4、4.5 mmol/LA-第一輪易錯PCR;B-第二輪易錯PCR圖6 外加不同Mn2+濃度進行易錯PCRFig.6 Different Mn2+ concentrations for error-prone PCR
將易錯片段miox4-2*和miox4-3*分別與其他片段融合得到突變的miox4*擴增產物,以質粒pY26-miox4-udh為模板擴增URA3,將突變的miox4*和URA3進行融合得到URA3-miox4*片段3 089 bp(圖7),融合片段轉化到菌株BY4741opi1Δ-udh中,構建突變體文庫。

泳道1-第一種易錯PCR情況與URA3融合的結果(miox4-2*與miox4-1、miox4-3-4、URA3融合);泳道2-第二種易錯PCR情況與URA3融合的結果(miox4-3*與miox4-1-2、miox4-4、URA3融合)圖7 兩種易錯PCR產物融合的結果Fig.7 The results of the fusion of two error-prone PCR products
2.4 利用大腸桿菌葡萄糖二酸傳感器對突變體文庫進行初篩
在大腸桿菌中,葡萄糖二酸經膜蛋白轉運到胞內后與cdaR結合變成激活形式,進而與操縱子區域結合,啟動下游報告基因gfp的轉錄[17, 22]。R7M10質粒同時表達轉錄因子cdaR和綠色熒光蛋白GFP,能將葡萄糖二酸濃度以熒光的強弱顯示出來[18]。為了確定熒光檢測的時間,以對照菌株BY4741opi1Δ-udh-miox4(未突變)的發酵液為對照,檢測不同比例的菌液和發酵液在不同時間的熒光/OD600值,實驗結果顯示,菌液和發酵液體積比為1∶1時在6~7 h檢測熒光效果較好(圖8)。
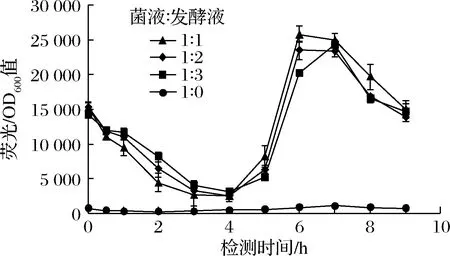
圖8 菌液和發酵液比例以及熒光檢測時間的確定Fig.8 Determination of the ratio of bacterial liquid and fermentation liquid, fluorescence detection time
利用大腸桿菌葡萄糖二酸傳感器作為一個高通量工具篩選突變體文庫,檢測突變株發酵液中的葡萄糖二酸。將帶有質粒R7M10的大腸桿菌BL21菌液以2%的接種量轉接到M9培養基中,然后與發酵液以1∶1的體積比加入到96深孔板中,培養6~7 h后用酶標儀檢測,篩選熒光較強的突變株,從熒光結果發現,1 200株菌中有60株熒光較高且高于對照的菌株(圖9)。
為了進一步確定這60株菌的葡萄糖二酸的產量,對60株菌進行發酵復篩。液相結果顯示,有24株菌的葡萄糖二酸產量高于對照菌株(未突變菌株)(圖10),其中9、15、41產量最高,分別是1.08、1.01、1.18 g/L,分別較對照菌株提高了34%、26%、48%。
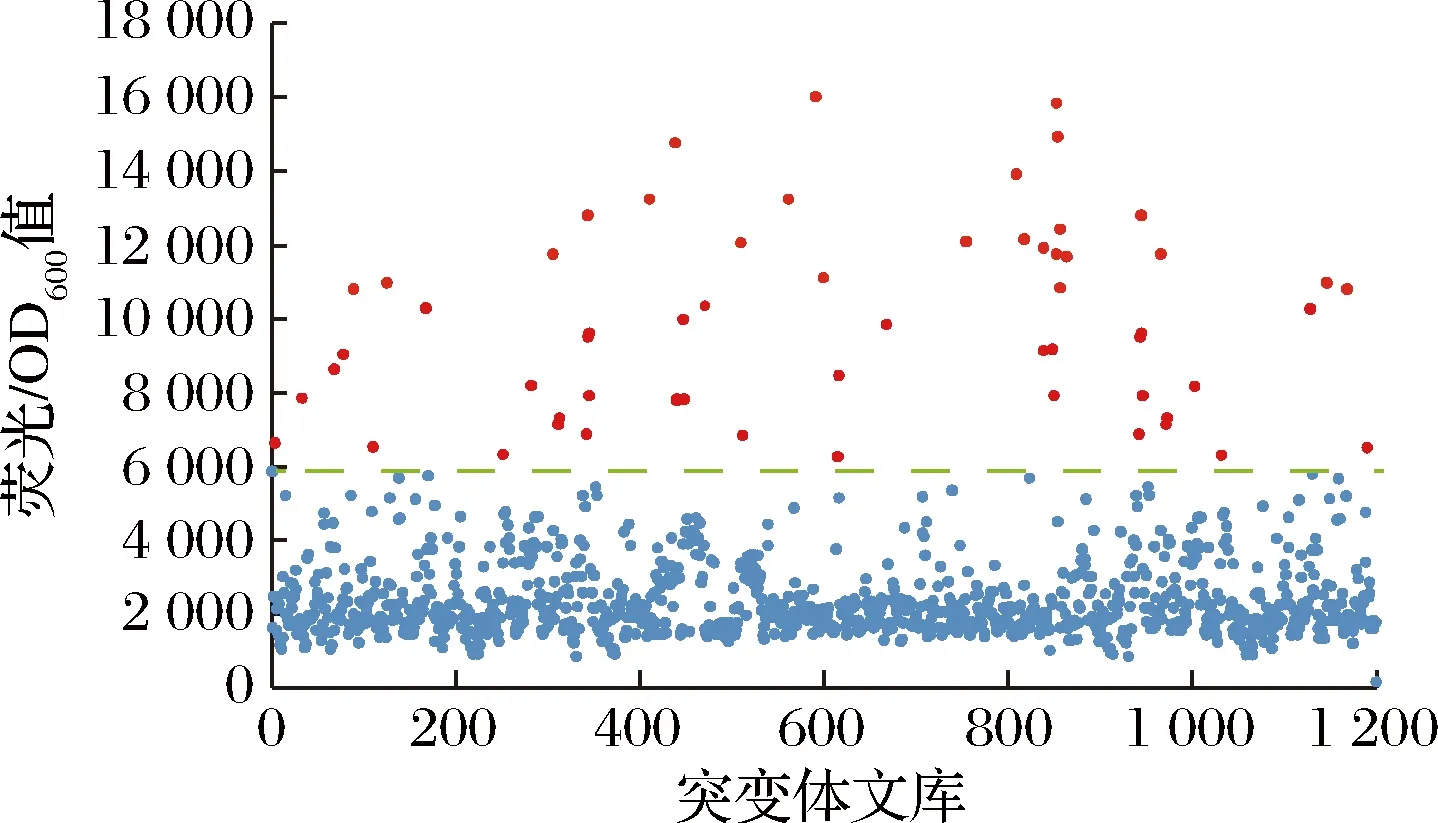
圖9 大腸桿菌葡萄糖二酸傳感器質粒對突變體文庫初篩的熒光結果Fig.9 Fluorescence results of screening of mutant library by E.coli glucaric acid biosensor
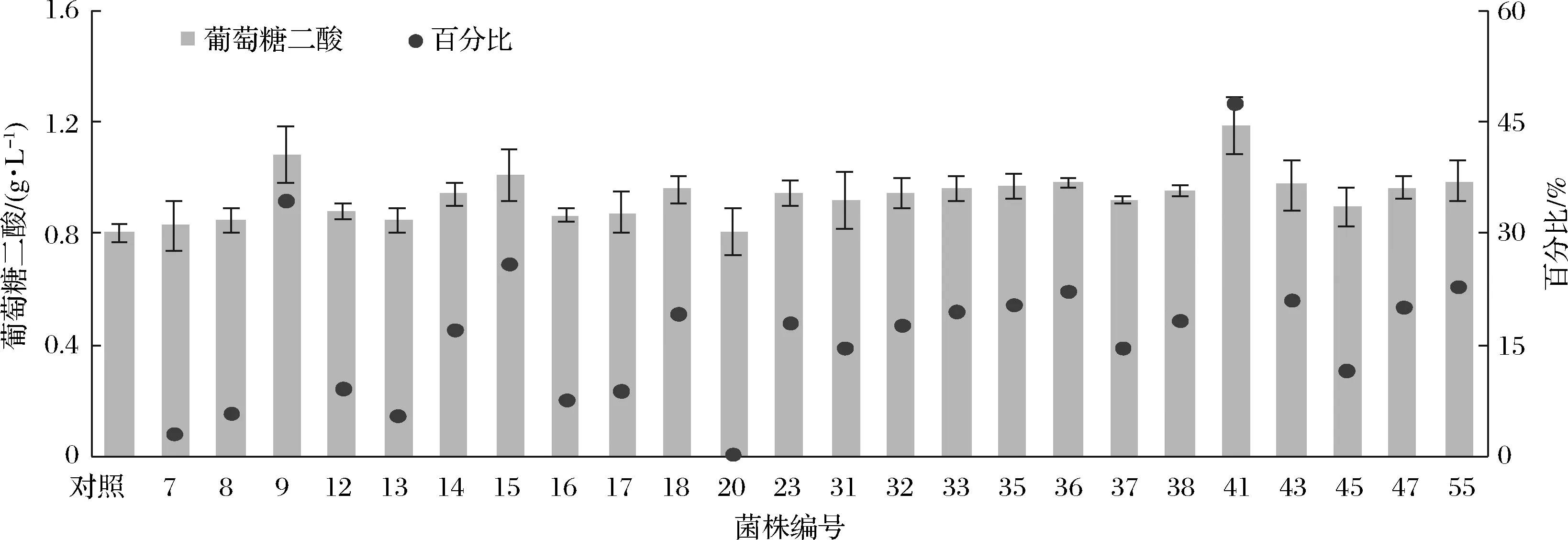
圖10 60株熒光較高的菌株進行發酵復篩Fig.10 Fermentation rescreening of 60 strains with higher fluorescence
2.5 突變菌株突變位點的確定及搖瓶發酵
為了鑒定上述24株菌的MIOX4的突變位點,對這24株菌的MIOX4序列進行了DNA測序。測序結果顯示,第8、9、12、15、41、45號菌株的MIOX4的氨基酸序列發生了突變,其余沒有發生突變,MIOX4的突變位點如表3所示。為了進一步確定這些突變位點是否為有益突變,對這6株突變株在外加60 mmol/L肌醇的培養基中進行搖瓶發酵,以未突變的菌株作為對照菌株。生長進入對數期后,在24、48 h分別補5 g/L葡萄糖。實驗結果顯示,突變株S133R、K88R、E72V、T40P在發酵168 h時葡萄糖二酸產量較對照菌株產量分別提高了53%、30%、21%、17%(圖11),說明這4個位點為正向突變位點。
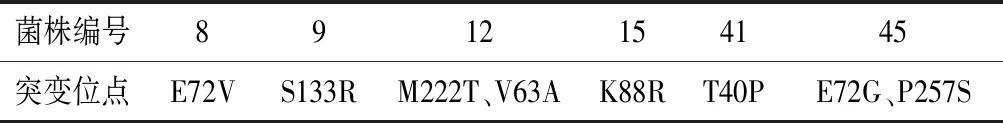
表3 突變位點的確定Table 3 Determination of the mutation sites
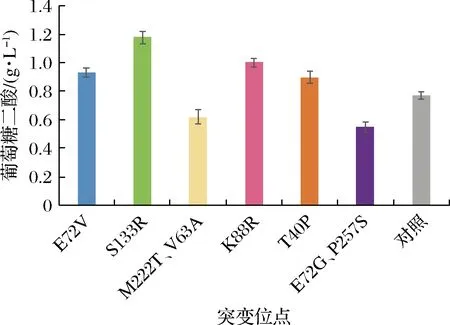
圖11 突變株搖瓶發酵結果Fig.11 Mutant shake flask fermentation results
2.6 MIOX4(S133R)蛋白三級結構分析
本研究中,我們對葡萄糖二酸產量提高最高的突變體MIOX4(S133R)進行蛋白結構分析,將突變后的MIOX4(S133R)蛋白序列輸入到蛋白三級結構在線預測平臺Swiss Model后,對突變蛋白進行同源建模,進一步分析突變后蛋白MIOX4(S133R)的氨基酸結構變化。分析結果顯示,MIOX4蛋白S133突變成R133后,Arg133和底物肌醇之間的距離變遠(圖12),這種距離的增加導致了空間位阻的減少。KARKHANE等[23]通過定點誘變的方法減少空間位阻,使突變后的Phe181與催化Ser114之間的距離增大,脂肪酶活性增加了2.6倍。薄秀梅等[24]將MEKK3蛋白K391突變為M391后,使靠近反應中心的原子側鏈占有的空間位置變大,增大了空間位阻而失去活性。空間位阻會限制底物進入酶活性的位點,因此空間位阻的減小可能會提高MIOX4的催化活性。
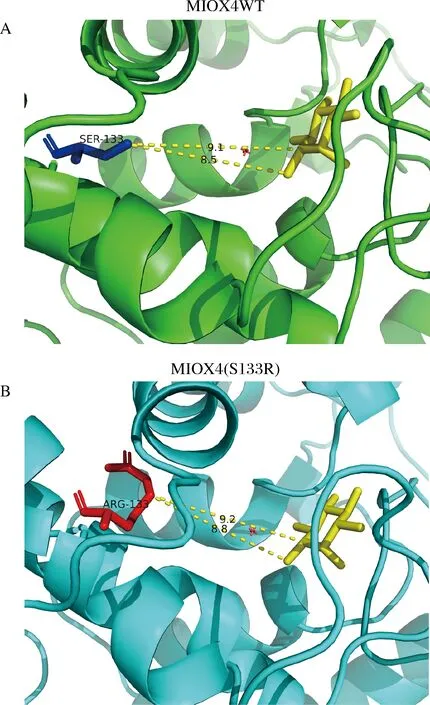
A-野生型MIOX4第133位的絲氨酸Ser與底物肌醇的空間距離;B-突變后133位的精氨酸Arg與底物肌醇的空間距離圖12 野生型MIOX4和突變型MIOX4(S133R)蛋白結構的比較Fig.12 Comparison of the protein structure of wild-type MIOX4 and mutant MIOX4(S133R)
3 結論
目前,葡萄糖二酸的產量經代謝工程及合成生物學的方法已大大提高[11-12],但肌醇轉化成葡萄糖醛酸的過程依舊是葡萄糖二酸代謝的限速步驟,因此提高肌醇加氧酶MIOX4的活性顯得尤為重要。定向進化是一種非常強大的蛋白質工程工具,可以在不深入了解蛋白質結構和酶-底物相互作用的情況下改善酶的性能。本研究以釀酒酵母為宿主,對來自擬南芥的肌醇加氧酶MIOX4進行定向提高其活性。采用易錯PCR的方法來構建MIOX4突變體文庫,利用大腸桿菌葡萄糖二酸傳感器將葡萄糖二酸產量轉變為熒光強度對突變體文庫進行初篩,得到了60株熒光較高的菌株。通過進一步對這60株菌株進行搖瓶發酵,獲得了24株葡萄糖二酸產量高于對照的菌株。最后發現,4個MIOX4的突變體(S133R、K88R、E72V、T40P)的葡萄糖二酸產量較野生型分別提高了53%、30%、21%、17%,其中S133R的產量提高最顯著。本研究結果同時進一步證明了MIOX4在葡萄糖二酸生物合成過程中起著重要作用,為后續提高葡萄糖二酸的產量奠定了基礎。
