野生鳥類腸道菌群多樣性與生態適應研究進展
2022-11-28 08:36:00莫麒穎李明軒周一凡劉玲張雪漣張婉婷吳弘趙大鵬
四川動物 2022年6期
莫麒穎,李明軒,周一凡,劉玲,張雪漣,張婉婷,吳弘,趙大鵬
(天津師范大學生命科學學院,天津300387)
動物腸道中存在一個龐大的微生物群落,其與宿主動物的相互作用參與了個體的營養吸收(B?ckhedet al.,2005)、生長發育(Diaz-Sanchezet al.,2012)、免疫防御(de Kivitet al.,2014)以及行為調節(Davidsonet al.,2020)等過程,在穩態維持和生態適應等方面起著重要作用(Moeller & Sanders,2020)。腸道菌群通常被描述為生物體內一個重要的“微生物器官”(B?ckhedet al.,2005),它們與宿主動物構成的整體被定義為“共生有機體”(Shapira,2016)。
鳥類是進化非常成功且極富物種和基因多樣性的一類生物(Waite&Taylor,2014),相較于哺乳動物,鳥類的腸道菌群可能更具低穩定性和高可塑性(Hird,2017)。首先,鳥類的直腸很短,消化物不經貯存便直接作為糞便排出體外(Duke,1997),這種快速的消化過程導致了鳥類腸道菌群極易被改變和調整(Laviad-Shitritet al.,2019);其次,鳥巢是一個高度可變且微生物組成復雜的結構(Main?waringet al.,2014;Hird,2017),其環境影響著雛鳥腸道菌群的建成(Gonzalez-Braojoset al.,2012;Am?brosiniet al.,2019);更重要的是,野生鳥類具有多樣的飲食類型、飛行習性和發育策略(García-Amadoet al.,2018),使其腸道菌群更為復雜(周立志,2018)。野生鳥類腸道菌群的變化影響著宿主的生理特征、營養狀態和應激反應等方面(Laviad-Shitritet al.,2019)。腸道菌群的動態調整常作為鳥類提高自身生態適應性的一種重要機制(Riselyet al.,2018;Bodawattaet al.,2022a)。
全基因組概念指出宿主基因組及其微生物組形成一個共同經歷自然選擇的進化單元(Rosen?berg & Zilber-Rosenberg,2016;Shapira,2016)。宿主為益生菌群提供合適的生態位以供它們較其他菌群更好地生長繁殖,而腸道菌群可以通過調節宿主食性生態位、放大使表型可塑性的信號以及促進免疫系統發育來促進生物的生態適應和進化歷程(Moeller & Sanders,2020)。樹袋熊Phasco?larctos cinereus幼崽在接種了專門針對一種桉樹的腸道微生物后會攝入更多該種桉樹的樹葉(Blytonet al.,2019);候鳥在遷徙過程中由禁食產生的壓力信號導致腸道中厚壁菌門Firmicutes 占比增加,這可能有利于它們最大效率地攝入能量(Turjemanet al.,2020);黑禿鷲Coragyps atratus和紅頭美洲鷲Cathartes aura腸道中富含2 類對于其他鳥類通常具有致病性的菌屬,它們可能有助于野生禿鷲消化腐肉,并抵御其他適應腸道生態位的細菌侵入(Roggenbucket al.,2014)。
鳥類復雜的生存模式以及宿主-微生物群的相互作用,預示著鳥類腸道菌群具有宿主特異性(Sekirovet al.,2010),其腸道中可能富集著能適應宿主腸道生態位且有益于宿主健康的微生物(Sha?pira,2016)。但鳥類個體小、飛行能力強、生活習性復雜等導致對鳥類腸道微生物的采樣和研究難度都大大增加(Waite & Taylor,2014)。鳥類占據著廣泛的生態位,在各類生態系統中均發揮著重要生態作用。然而,當今世界全球性環境問題對自然生態系統造成的影響和破壞日趨嚴重(李秉成,2006),鳥類腸道菌群也因此受到影響(Wuet al.,2018;Berlowet al.,2021a)。這不僅影響鳥類的生存,還關系到再引種的成敗(Redfordet al.,2012)。因此,關注野生鳥類腸道菌群多樣性具有重要研究價值和生態價值,探究宿主因素和環境因素對野生鳥類腸道菌群的影響,解釋野生鳥類腸道菌群與其生態適應的關系,能夠更深入地了解腸道微生物與宿主協同進化的生理機制,為生物多樣性維持及生態環境保育工作提供基礎理論依據。據此,本文歸納整理了現有關于野生鳥類腸道菌群的研究成果,總結了野生鳥類腸道菌群與宿主和環境的相互關系,為進一步探究野生鳥類腸道菌群的建成機制和生態作用提供參考。
1 研究方法
本研究以“腸道微生物”“腸道菌群”與“野生鳥類”為關鍵詞,在中國知網(www.cnki.net)進行檢索,以“gut microbiota”“intestinal flora”“gut microbi?ome”“intestinal microbiota”“microbial community”與“wild birds”“wild avian”為關鍵詞,在數據庫Web of Science(http://www.webofscience.com/)中進行檢索。通過對摘要和全文的閱讀,保留研究野生鳥類腸道微生物多樣性與結構特征,或探討某種因素對野生鳥類腸道微生物的影響效應的相關論文,并著重分析研究結果體現腸道菌群與鳥類生態適應性相關的研究論文。
本研究記錄所有論文的基本信息,包括文獻基礎信息、研究物種信息、研究方法等。將調研文獻中的每個樣本視為獨立群體,統計所有研究群體的采樣地點、分類地位、瀕危等級、保護級別以及樣本容量。數據分析采用Excel 2019、OriginPro 2021、Adobe Photoshop CC 2019。
2 研究進展
截至2022 年4 月,共搜集到符合要求的文獻123 篇(期刊論文109 篇,學位論文14 篇)。本研究發現1992—2011 年相關論文發表數量無明顯增長趨勢,而2012—2022 年4 月相關論文發表數量呈顯著增長趨勢;國內研究起步晚于國外研究,但其增長速度較快(圖1),其中中文文獻共16篇。

圖1 已檢索到的國內外相關研究論文數量Fig. 1 Number of related domestic and foreign literatures
2.1 物種組成多樣性
在研究物種的分類地位方面,當前研究物種共涉及野生鳥類24 目87 科212 屬303 種。其中,在目水平上,以雀形目Passeriformes(54.1%)、鸻形目Charadriiformes(12.5%)和雁形目Anseriformes(4.6%)為主;在科水平上,以裸鼻雀科Thraupidae(9.6%)、鷸科Scolopacidae(6.9%)和鴨科Anatidae(4.6%)為主。
在研究物種的采樣地區方面,共涉及6洲28國。其中,大洲以亞洲(32.5%)、南美洲(17.1%)和非洲(14.9%)為主;國家以中國(29.4%)、赤道幾內亞(14.7%)和美國(12.9%)為主(圖2)。
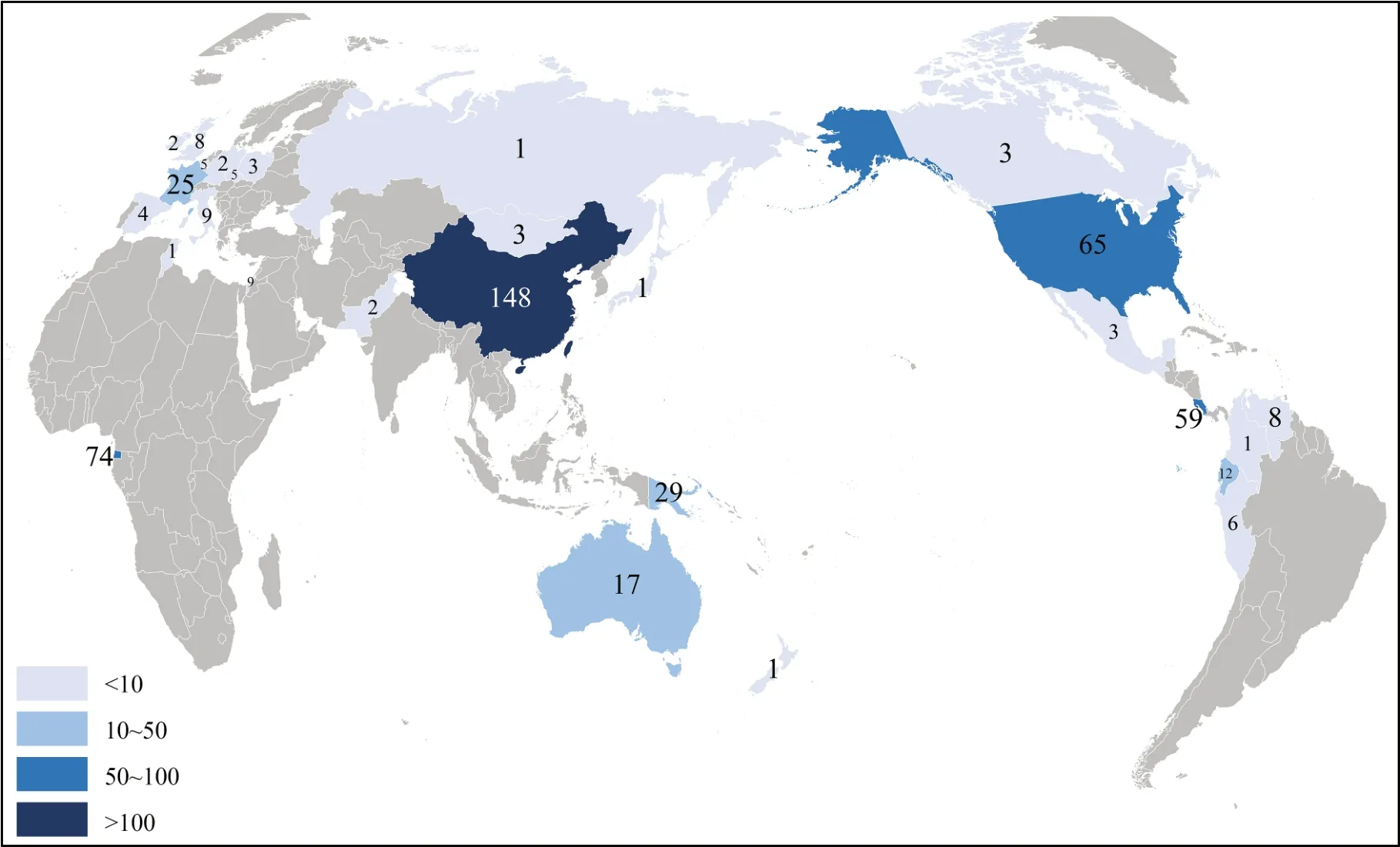
圖2 已研究的野生鳥類群體的國家分布情況Fig. 2 National distributions of known wild bird clusters
在研究物種的瀕危等級方面,世界自然保護聯盟(IUCN)紅色名錄中“無危(LC)”物種占77.1%。在研究物種的保護級別方面,研究地點在我國,且為“國家一級重點保護野生動物”的占41.9%。
2.2 研究方法多樣性
2.2.1 采樣方法選擇現有研究主流的采樣方法為“間接采樣(糞樣)”(74.8%),且隨著時間的推移,該方法使用比重呈上升趨勢,多種采樣方式結合的形式開始出現(表1)。

表1 不同時間段各采樣方法的數量Table 1 Number of sampling methods in different time periods
2.2.2 研究對象選擇在研究物種的樣品數量方面,樣本容量為1~546,以1~20(77.9%)為主;整體趨勢表現為瀕危等級越高,對應的樣本容量越小(圖3)。在研究物種、地區和時期的數量選擇方面,以對單物種(64.2%)、單地區(65.9%)和單時期(82.1%)的研究為主。
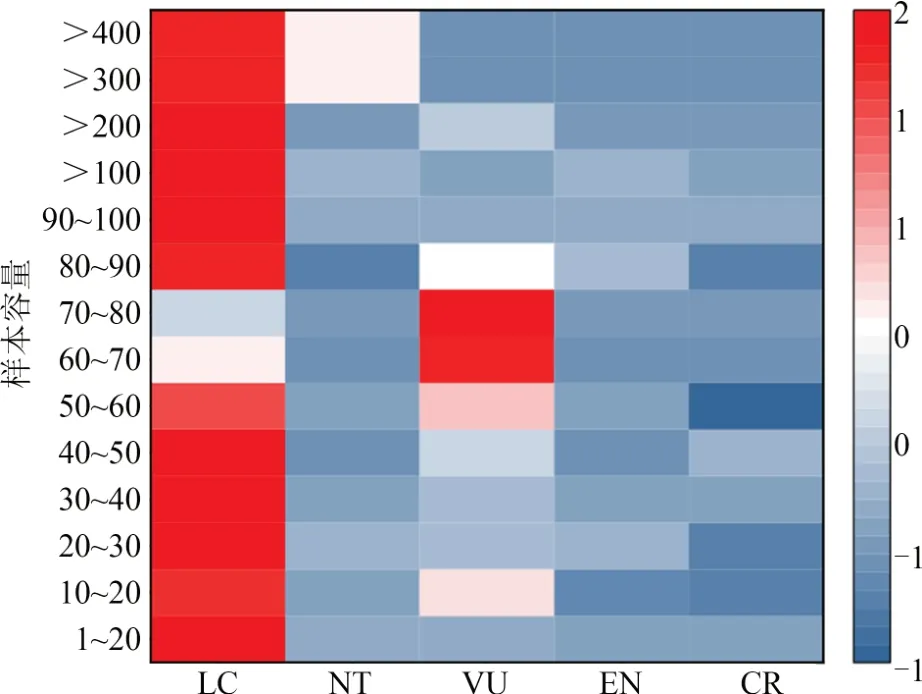
圖3 樣本容量與IUCN瀕危等級的聯系Fig. 3 Association between sample sizes and IUCN classification
2.2.3 實驗技術選擇早期研究受技術的限制,對腸道微生物的鑒定使用以純培養、染色和顯微鏡觀察為主的方法(Aguirreet al.,1992)。這類方法操作簡易,但周期較長、結果受環境影響較大(劉玉婷等,2016),且鳥類腸道微生物中可培養的菌種百分比未知(張政,2021)。
基于16S rRNA 和18S rRNA 基因包含著不同物種特有保守基因序列的原理(Zaneveld,2010),現有研究主要通過分子生物學技術對腸道菌群進行檢測。早期測序常用的自動化核糖體基因間隔分析(automated ribosomal intergenic spacer analy?sis,ARISA)(van Dongenet al.,2013)可直接用來推測生物體腸道菌群的變化。也可通過克隆技術對優勢菌群進行進一步分析(劉玉婷等,2016)。隨著高通量測序技術的發展,Illumina 等測序平臺的出現,給研究者提供了更加全面認識腸道微生物組成特征和物種信息的條件,16S rRNA 高通量測序技術已經成為檢測野生鳥類腸道細菌群落的主要途徑(Waite & Taylor,2015)。基于16S rRNA 高通量測序結果,研究者通常使用微生物生態學定量分析(quantitative insights into microbial ecology,QIIME)對微生物原始數據進行歸一化處理(Caporasoet al.,2010)后,對所得數據進行操作分類單元(operational taxonomic units,OTU)劃分,繼而對其進行測序質量評估、物種組成分析、Alpha多樣性分析以及Beta多樣性分析(趙晗旭,2015;Xieet al.,2016),并使用 PICRUSt 進行功能預測(Langilleet al.,2013)。
宏基因組測序是一種新興的對腸道微生物進行檢測和表征的技術(張政,2021),可以全面反映環境中所有微生物的基因信息(Mendozaet al.,2018),其在反映和測量腸道菌群組成特征以及基因組編碼代謝潛力方面有強大的功能(Tuohy,2009),但更大的測序深度也增加了分析的成本和計算難度(董元秋,2019),當前關于野生鳥類腸道菌群宏基因組的研究較少(Wanget al.,2019;Moh?sin Bukhariet al.,2022)。
2.3 腸道菌群多樣性
在門水平上,野生鳥類的腸道菌群以變形菌門Proteobacteria、厚壁菌門、擬桿菌門Bacteroide?tes、放線菌門Actinobacteria 為優勢菌門,其次還包括軟壁菌門Tenericutes、藍細菌門Cyanobacteria等。縱、橫向比較各研究中鳥類群體腸道菌群的結構發現,各門的豐度和結構在種間和種內均存在較大差異,這可能是由于野生鳥類腸道菌群的構建受到多種因素的共同影響。生活在比利時北部一林地中的大山雀Parus major以厚壁菌門、放線菌門和變形菌門腸道為優勢菌門,其中厚壁菌門和放線菌門占絕對優勢(Teyssieret al.,2018);而生活在愛爾蘭科克市的大山雀以厚壁菌門、變形菌門和軟壁菌門為優勢菌門(Davidsonet al.,2021)。以昆蟲為食物的大山雀個體腸道菌群以變形菌門、厚壁菌門和軟壁菌門為主,而以種子為食物的個體腸道菌群以變形菌門、軟壁菌門和藍細菌門為主(Davidsonet al.,2020)(表2)。

表2 部分野生鳥類腸道菌群多樣性Table 2 Diversity of gut microbiota of some wild birds
在屬水平上,野生鳥類腸道菌群也存在著豐富的變化。斑頭雁Anser indicus在青海湖附近一濕地生境下以乳球菌屬Lactococcus、假單胞菌屬Pseudomonas和節細菌屬Arthrobacter為優勢菌屬(Wanget al.,2016a),而同樣在該地區,另一研究結果顯示其以鏈球菌屬Streptococcus、乳球菌屬和芽孢桿菌屬Bacillus為優勢菌屬(Wanget al.,2017);在西藏各越冬區內,斑頭雁以乳球菌屬、芽孢桿菌屬和假單胞菌屬為優勢菌屬(Wanget al.,2016b);在青海湖生境下斑頭雁以梭桿菌屬Fusobacterium、嗜冷桿菌Psychrobacter和Bulleidia為優勢菌屬(鄭思思等,2018)。
2.4 影響因素多樣性
野生鳥類腸道微生物可通過遺傳獲得,也可來源于環境(Theiset al.,2016;Trevellineet al.,2018)。Shapira(2016)提出將腸道菌群分為 2 類:宿主核心菌群和取決于環境條件而瞬時變化的菌群。野生鳥類腸道菌群的建成是宿主內在因素和外界環境因素共同驅動的結果(Hirdet al.,2015)。
2.4.1 宿主因素對腸道菌群的影響食性因素:食性被認為是影響腸道菌群結構最主要的內驅力因素(Carmodyet al.,2015)。植食性和雜食性鳥類腸道菌群多樣性通常很豐富(章娜中,2021),前者以變形菌門、厚壁菌門為主,而后者腸道菌群組成更復雜(Wanget al.,2018;王娟等,2021),有利于消化更多種類的食物(Wanget al.,2019)。目前對肉食性鳥類腸道菌群的研究較少,現有結果顯示其腸道菌群多樣性較低(章娜中,2021),梭桿菌門豐度較高(高澤中,2020)。植食性和雜食性鳥類腸道菌群受季節影響顯著(章娜中,2021),而肉食性鳥類腸道菌群的季節性差異不顯著(Michelet al.,2018)。腸道菌群中一些菌群的存在往往與宿主特 殊 的 飲 食 相 適 應(de Carvalhoet al.,2003;Roggenbucket al.,2014;Ushidaet al.,2016;Michelet al.,2018)。與圈養個體相比較,野生日本巖雷鳥Lagopus muta japonica擁有更豐富多樣的腸道菌群,且呈現部分特有菌群,這可能與其食用含植物毒素的食物有關(Ushidaet al.,2016)。
遺傳因素:雌鳥的腸道微生物可以垂直遺傳給子代,但這種影響是有限的(Kreisingeret al.,2017)。在實驗控制和野生環境下,鳥類物種遺傳距離與腸道菌群結構相似程度之間呈負相關(Ca?punitanet al.,2020;Trevellineet al.,2020)。生活于同一區域的不同物種,雖然環境條件一致且存在腸道微生物的跨種傳播,但其腸道菌群結構仍存在顯著差異(Grondet al.,2014;楊預展,2016;Yang&Zhou,2021)。
年齡因素:野生鳥類腸道中特定微生物的相對豐度隨年齡和時間的變化而變化(Spergseret al.,2018;Zhouet al.,2020)。即使在相同的環境下,不同年齡段的個體由于體內特異性環境或生理因素的差異,其核心腸道菌群差異顯著(Gillinghamet al.,2019;Wanget al.,2020)。
性別和繁殖因素:不同性別和繁殖階段的個體之間腸道菌群的差異與兩性在整個繁殖季節行為和飲食的變化相一致(Gongoraet al.,2021)。鳥類在繁殖季節可能周期性地獲得新的細菌,然后在非繁殖季節失去它們(Escallonet al.,2019)。這可能與繁殖活動需要大量的能量有關(尚偉平,2021)。
居留方式因素:遷徙鳥類在遷徙過程中會接觸到各種外界環境,面臨能量需求、環境壓力、疾病免疫等方面的挑戰,其腸道菌群的多樣性可能高于留鳥(Riselyet al.,2017,2018;董元秋,2019)。
個體體型、健康狀態等因素也影響著野生鳥類腸道菌群的建成。體型較大的雀形目鳥類個體間腸道菌群的差異更小,且與環境食物共享更少的微生物(Berlowet al.,2021a;Bodawattaet al.,2022b);腹瀉個體與健康個體的腸道菌群結構差異顯著(Xiet al.,2021)。
2.4.2 環境因素對腸道菌群的影響環境對野生鳥類腸道菌群建成的影響因素包括生物和非生物因素的差異(Hirdet al.,2014),即食物因素(趙廣紅,2016)、人為因素(Fuirstet al.,2018)、季節(Donget al.,2019)、地理因素(Looet al.,2019)和環境微生物因素(Xianget al.,2019)等。
食物因素:當鳥類飲食組成發生變化時,腸道微生物群落的Alpha 多樣性不會發生顯著變化,而Beta 多樣性會發生顯著變化(白頭鶴Grus mona?cha:Zhaoet al.,2017;章 娜 中 ,2021;大 鴇Otis tarda:Luet al.,2021;青頭潛鴨Aythya baeri:Xiet al.,2021;橙腹果鳩Ptilinopus iozonus:Bodawattaet al.,2022b)。這可能是由于雖然食物組成存在差異,但個體食性始終一致(章娜中,2021),食物的差異對鳥類腸道菌群的影響主要是針對一些特定的菌屬,而不是對菌群整體的改變(Bodawattaet al.,2022b)。棲息生境的植被組成很大程度上決定了環境食物的復雜性,從而影響鳥類腸道菌群的多樣性和組成(Drobniaket al.,2021;Goossenset al.,2022)。在食源豐富的生境中,鳥類腸道微生物的豐度和多樣性通常更高(東方白鸛Ciconia boyci?ana:Wuet al.,2021;赤麻鴨Tadorna ferruginea:杜菲等,2022)。相似生境中的食物資源相似,因此鳥類腸道菌群的結構也更相似(楊竹青,2021;Zhanget al.,2021)。而在食物類型有限的生境下,鳥類腸道菌群結構簡單(杜菲等,2022)、種間差異小(Wuet al.,2018)。
人為因素:頻繁的人類活動、噪音污染等人為干擾加大了野生鳥類的生存壓力,可能引起其免疫-體液調節系統的調整,從而導致腸道菌群的改變(Ber?lowet al.,2021b)。人為干擾程度越高,對鳥類腸道菌群的影響程度越大(Knutieet al.,2019),可能導致腸道菌群多樣性降低(Maulet al.,2005),而產生更多的潛在致病菌(Ushidaet al.,2016;顧晶晶,2021)。
季節、地理因素:季節和地理條件常通過影響食物間接影響鳥類腸道菌群(Davidsonet al.,2020)。季節更替使棲息地食物發生變化,可能導致鳥類腸道菌群出現季節性波動,冬季野生鳥類腸道微生物的Alpha 多樣性更高(Zhanget al.,2020;Mahtabet al.,2021;Wanget al.,2021a);不同海拔、水熱條件等地理因素改變了野生植被的類型,這可能促進了鳥類腸道菌群的分化(Looet al.,2019;Wanget al.,2020);農村生境中食物會發生季節性變化,這不僅會引起鳥類腸道菌群的季節性變化,還會增加鳥類定期獲取新菌群的概率(Fuirstet al.,2018;Teyssieret al.,2018,2020),而城市對鳥類腸道菌群的效應也受環境中食物組成的影響,當城市環境可以提供多種營養時可以提高腸道微生物多樣性,反之則會降低(Teyssieret al.,2018;Murrayet al.,2020)。
環境微生物因素:一方面,鳥類腸道菌群會受到食源性微生物(Davidet al.,2014;Bodawattaet al.,2022b)以及來自土壤(Yanget al.,2016;章娜中,2021)和巢穴(Ambrosiniet al.,2019;Gillinghamet al.,2019)中微生物的影響;另一方面,由于不同個體直接或間接的接觸也會導致腸道菌群的種內傳播(Kreisingeret al.,2017)和跨種傳播(Gu & Zhou,2021;Wanget al.,2021b)。環境微生物對候鳥的影響較顯著,即使只是短暫停歇也會導致鳥類腸道菌群與環境微生物的趨同(Turjemanet al.,2020)。此外,受流感病毒等微生物感染的鳥類腸道菌群也會出現顯著變化(綠頭鴨Anas platyrhynchos:Ganzet al.,2017;大天鵝Cygnus cygnus:Zhaoet al.,2018)。
2.4.3 內外因素共同影響腸道菌群宿主因素決定了種間差異,而不同生境也導致了腸道微生物結構的明顯分化(Yanget al.,2016)。在長途遷徙過程中,雖然候鳥生存環境和飲食發生了明顯變化,但其核心菌群基本保持穩定(Wuet al.,2018;王偉,2021;Zhaoet al.,2021),這可能是由于宿主核心菌群對腸道中微量的環境微生物具有特異性拮抗作用(Riselyet al.,2018;王偉,2021),因此鳥類腸道環境可以快速區分并選擇定植的細菌群落(Gillinghamet al.,2019)。但部分研究結果與之相反,Hird 等(2014)研究發現,寄生于不同物種巢穴中發育成熟的成年棕頭燕八哥Molothrus ater個體之間不存在一個共同的腸道核心菌群,這表明宿主內驅力作用可能并不顯著;Goossens 等(2021)關于普通Sitta europaea的研究結果與之類似。這在一定程度上意味著多種生態因素對腸道菌群的綜合影響可能超過了宿主因素的單一效應(Barbosaet al.,2016;Lewiset al.,2016;Grondet al.,2019)。可見,內外因素對野生鳥類腸道菌群建成的貢獻程度目前尚無明確定論。
3 野生鳥類腸道菌群的生態適應
動物腸道菌群在生態適應方面表現出高度的可塑性(Weinhold,2022)。腸道菌群的動態變化可以通過提高消化效率、增強免疫能力、維持體內穩態等方式來提高宿主生存適應性。
3.1 腸道菌群適應宿主飲食的各種變化
飲食對腸道微生物群變化的影響是由飲食成分和宏觀營養素共同驅動的(Bodawattaet al.,2021)。腸道菌群對宿主飲食差異高度敏感(Davidet al.,2014;呂小艷,2021),可以對宿主飲食的變化迅速作出反應(趙廣紅,2016),以促進宿主對特定生態位的適應,滿足其對能量和營養的需求(Gonget al.,2021)。當宿主的飲食發生改變,腸道菌群的結構和功能也會隨之改變(楊預展,2016;Liet al.,2021)。
腸道菌群對鳥類飲食季節性變化的適應。雜色山雀Sittiparus varius是雜食性鳥類,在夏季主要為肉食性,在春、冬季為植食性,其春、冬季腸道中的厚壁菌門豐度高于夏季(高澤中,2020),由于厚壁菌門在降解纖維方面起著關鍵作用(Flintet al.,2008),因此這種轉變有利于改善鳥類對營養的吸收。白頭鶴(Fuet al.,2020)、大鴇(Luet al.,2021)、灰鶴Grus grus(Pekarskyet al.,2021)的研究結果與之類似。
腸道菌群對鳥類越冬時期飲食變化的適應。白頭鶴在越冬前期以稻谷為主要食物,腸道中主要以參與植物蛋白質分解代謝的普氏菌屬Prevotella為優勢菌,而在以高纖維植物為主要食物的越冬中后期,具有降解腸道中纖維素作用的梭菌屬Clos?tridium為優勢菌(趙廣紅,2016)。白頭鶴(Zhanget al.,2021)、黑 頸鶴Grus nigricollis(Zhaoet al.,2021)的研究結果與之相似。
腸道菌群對鳥類在復雜生境中飲食多樣化的適應。相較于野生類群和圈養類群,半圈養斑頭雁群體中有大量擬桿菌門出現,而擬桿菌門具有高度可塑性,可以很快地適應各種生態位(董元秋,2019)。因此腸道菌群的這種轉變可能有助于斑頭雁適應野生和人工不同的食物資源,使宿主腸道能夠在不同能量消化模式之間輕松切換(Wanget al.,2016a)。在越冬早期,候鳥腸道菌群多樣性通常豐富且多變,這可能由于在長途飛行后腸道建立了對新環境的耐受性和適應性,以最大效率消化食物、補充能量(Zhaoet al.,2021)。
3.2 腸道菌群適應長途飛行的生理挑戰
候鳥在遷徙飛行期間會面臨能量、免疫等方面的生理挑戰(張政,2021),它們已經進化出一系列與遷徙習性相適應的生理特征,而腸道菌群的改變是其適應長途飛行的潛在機制之一(Riselyet al.,2018)。
候鳥長途飛行期間處于長期禁食狀態,研究結果顯示,長途飛行時鳥腸道中厚壁菌門的豐度增加(加拿大黑雁Branta canadensis:Obrochtaet al.,2022),或厚壁菌門與擬桿菌門豐度比(F/B)升高(白頭鶴:付蓉,2020;大天鵝:Wanget al.,2021a)。這可能是由禁食產生的壓力信號所引起,當動物面對環境壓力產生應激反應時通常會分泌糖皮質激素(Martin&Reale,2008),而糖皮質激素的分泌會導致鳥類腸道菌群的改變(Nogueraet al.,2018)。腸道菌群對宿主壓力信號的響應可能是其幫助鳥類適應長途飛行的基礎(Turjemanet al.,2020)。
在適應能量需求方面,研究表明厚壁菌門的基因編碼了許多與能量代謝相關的酶,這些消化酶可以代謝各種物質(Kaakoush,2015),而F/B 的增加會提高機體代謝脂肪的能力,增加腸道吸收營養和轉化能量的能力(B?ckhedet al.,2005;Ya?davet al.,2021)。因此,腸道菌群的這種轉變有利于宿主消化吸收營養物質,提高宿主能量代謝的效率(Kaakoush,2015;Wanget al.,2018)。
在適應疾病免疫方面,糖皮質激素的增加可能降低了宿主對機會性病菌的易感性(Dhabhar,2009),F/B 的增加可以顯著地抑制腸道致病菌的定植(Heet al.,2019)。Davies 等(2022)的研究表明,宿主免疫系統可以通過調節腸道微生物組來調節宿主的健康,微生物組也可能影響免疫基因的選擇性表達。因此,腸道菌群的這種轉變有利于提高宿主免疫能力,宿主與腸道菌群的相互作用可能在維持鳥類功能性免疫遺傳變異中發揮重要作用。
除此之外,值得關注的是候鳥腸道中棒狀桿菌屬Corynebacterium豐度顯著高于留鳥(如紅頸濱鷸Calidris ruficollis:Riselyet al.,2017;彎嘴濱鷸Calidris ferruginea:Riselyet al.,2018;家燕Hirundo rustica:Turjemanet al.,2020)。在同一圈養環境下生長的15 種鶴科Gruidae 鳥類中,短途遷徙物種藍蓑羽鶴Anthropoides paradiseus腸道中棒狀桿菌屬豐度較其他長途遷徙物種的更低(Trevellineet al.,2020)。這可能意味著棒狀桿菌參與了候鳥與微生物的功能性相互作用,使候鳥在遷徙過程中最大限度地增加脂肪沉積和能量攝入(Riselyet al.,2018)。
3.3 腸道菌群適應生境擾動的生存挑戰
由于人類活動范圍的擴張和強度的增加,棲息生境破碎、氣候變化、噪聲污染、光污染等一系列環境問題都導致了野生鳥類巨大的生存壓力(Bestionet al.,2017;Zhaoet al.,2017;姜 俊霞 ,2019;Berlowet al.,2021a,2021b)。為了緩解生存壓力,許多鳥類通過調整自身行為或免疫系統,間接影響了腸道菌群的組成,從而提高了自身生存能力。
面對棲息生境破壞,鳥類會不斷調整覓食策略(Weiet al.,2020),而其多變的腸道菌群可能是水鳥面對濕地資源流失的一種適應機制(Donget al.,2019;章娜中,2021)。此外,一些鳥類腸道內還發現了抗有機污染物的細菌(栗斑腹鹀Emberiza jankowskii:尚偉平,2021)。
面對城市化,鳥類對城市棲息地的使用和飲食的變化顯著改變了其腸道微生物的組成和多樣性,如銀鷗Larus argentatus(Fuirstet al.,2018)、麻雀Passer domesticus(Teyssieret al.,2018;Gadauet al.,2019),這可能導致鳥類更容易感染病原菌(Mur?rayet al.,2020)。腸道菌群多樣性與腸道中病原菌之間存在負相關(Xianget al.,2019),腸道微生物可能通過競爭作用以及與宿主免疫系統的相互作用,抵御病原菌的定植,以保護宿主的健康(Lawleyet al.,2008;Murrayet al.,2020;Craftet al.,2022)。
雙色樹燕Tachycineta bicolor通過與免疫系統更好地耦聯作用,調節自身對熱的敏感性,以更好地適應熱環境(Ingalaet al.,2021),這可能是一些鳥類應對氣候變化的一種潛在機制。高海拔安氏蜂鳥Calypte anna腸道菌群中的擬桿菌門豐度為低海拔的7.0 倍,通過降低腸道中好氧菌豐度可能是一些鳥類應對人類活動造成其活動海拔升高的一種潛在機制(Herderet al.,2021)。噪音似乎能直接作用于腸道菌群使其多樣性升高(Berlowet al.,2021b),而腸道菌群多樣性越高,通常意味著機體對環境擾動的恢復能力越強(Teyssieret al.,2018),這可能是一些鳥類應對噪音污染的一種潛在機制。
4 總結展望
當前研究對野生鳥類腸道菌群的認識不斷深入,但與哺乳動物腸道菌群的研究相比,有關野生鳥類腸道菌群的研究仍然有一定距離(Kohl,2012)。基于現有研究,提出以下建議:(1)建立統一、系統的樣品采集方法,樣本容量、采樣方式、取樣部位、儲存方法的差異均會對腸道菌群檢測結果造成影響(Bodawattaet al.,2018;Drovetskiet al.,2018;Videvallet al.,2018;Vargas-Pelliceret al.,2019);(2)加強腸道菌群宏基因組學和代謝組學的研究,明確腸道微生物的生態功能,揭示宿主與腸道菌群相互作用的內在機制;(3)拓展研究物種范圍,不斷豐富完善鳥類腸道菌群的背景信息,并與人類和其他脊椎動物腸道菌群開展比較研究,深化對宿主-腸道微生物協同進化的認識;(4)增加多地區、多時期、多物種的對比研究,綜合探討內外因素對野生鳥類腸道菌群的影響機制,明確腸道微生物、宿主和環境三者之間的相互關系。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
