南海3種小型硨磲的外殼形態差異分析
2022-11-25 13:16:36鈔琪琪顧志峰王愛民PEZVioleta劉春勝
海洋科學 2022年10期
鈔琪琪, 楊 毅, 顧志峰, 2, 王愛民, 2, LóPEZ Violeta, 劉春勝, 2
南海3種小型硨磲的外殼形態差異分析
鈔琪琪1, 楊 毅1, 顧志峰1, 2, 王愛民1, 2, LóPEZ Violeta3, 劉春勝1, 2
(1. 海南大學 海洋學院, 海南 海口 570228; 2. 海南大學 南海海洋資源利用國家重點實驗室, 海南 海口 570228; 3. 西班牙國家自然科學博物館, 西班牙 馬德里 28006)
本研究對中國南海3種小型硨磲(spp.)[番紅硨磲(, 包括2種外殼形態)、長硨磲()和諾亞硨磲()]的外殼形態性狀進行生物統計學差異分析。研究表明, 4組硨磲外殼存在顯著形態學差異。單因素方差分析發現9個外殼比例性狀能夠較好區分3種硨磲, 且2種不同外殼形態的番紅硨磲在殼寬/殼長、足絲孔寬/殼長、殼質量/殼長和左右殼厚比殼長存在顯著性差異(<0.05); 在基于歐氏距離Ward法且指定聚類數= 4的條件下, 4組硨磲層次聚類效果較好; 主成分分析提取3個主成分建立散點圖, 累計主成分貢獻率73.85%, 各主成分貢獻率依次為50.47%, 13.78%和9.60%; 線性判別分析得出3個判別函數, 方差解釋率依次為70.13%、20.97%和8.90%。3種多元統計方法從不同角度解釋各組硨磲的外殼形態差異, 結果均顯示兩組番紅硨磲的外殼形態更為接近, 同時長硨磲與諾亞硨磲的外殼形態較為接近。
硨磲(spp.); 外殼形態; 多元統計分析; 南海
硨磲(Tridacninae)隸屬于軟體動物門(Mollusca)、雙殼綱(Bivalvia)、簾蛤目(Veneroida)、硨磲科(Tridacnidae), 目前已發現的硨磲有2個屬共12種, 其中硨磲屬()有10種, 硨蠔屬()有2種[1]。硨磲是海洋中最大的雙殼貝類, 生活在印度太平洋地區的熱帶和亞熱帶珊瑚礁海域。硨磲與蟲黃藻建立了互利共生關系, 為獲得足夠光照進行光合作用, 其大部分種類分布于20 m以淺的珊瑚礁或瀉湖區[2-4]。硨磲在珊瑚礁生態系統中發揮著重要作用, 對硨磲種類鑒定及生物多樣性分析是開展硨磲相關研究的前提[5]。然而不同種類的硨磲外部形態極其相近, 國內外亦少有關于硨磲形態學系統分類的報道[6]。
貝類物種鑒定主要有分子學和形態學兩大手段, 當前形態學與分子學相結合的鑒定方法依舊是硨磲分類的重要依據。SU等[6]基于外殼形態之間的顯著性差異, 同時結合分子手段, 確定硨磲屬內一個正式新種—諾亞硨磲()。西印度洋隱存種和的發現也是利用外殼形態結合基因鑒定技術[7]。雙殼貝類的傳統形態鑒別方法主要依據各項外殼形態指標進行多元統計分析, 進而鑒定不同類群或推斷種間或種內類群之間的差異[8]。魏開建等[9-10]運用聚類分析、主成分分析和判別分析對蚌的外殼形態差異進行研究; 張永普[11]同樣運用這3種方法分析泥蚶種內形態變異特點與地理分化規律。吳楊平[12]通過單因素方差分析、聚類分析、主成分分析、判別分析及Mantel檢驗對3種文蛤進行地理種群形態差異性研究。MEI[13]對不同處理組干殼重量和殼強度均值進行方差分析和典型判別分析。
在中國, 硨磲分布于臺灣澎湖列島和海南島以南熱帶海域。迄今已發現有 8種, 包括硨磲屬的番紅硨磲()、大硨磲()、無鱗硨磲()、鱗硨磲()、長硨磲()、諾亞硨磲()和硨蠔屬的硨蠔()和瓷口硨磲(), 其中小型的番紅硨磲、長硨磲以及諾亞硨磲分布最為廣泛[6, 14-15]。本研究以中國3種常見小型硨磲為對象, 采用單因素方差分析、秩和檢驗、聚類分析、主成分分析以及判別分析對其外殼性狀進行描述分類統計, 研究不同外形和品種的硨磲在各個外殼形態指標的差異, 探討各指標在硨磲的分類鑒定方面的綜合影響, 為硨磲的形態分類提供支撐。
1 材料方法
1.1 實驗樣品
在中國南海采集3種硨磲, 其中番紅硨磲中存在外殼形態扁平和圓鈍兩類, 分別命名為-1組(32只)和-2組(22只)、長硨磲(17只)、諾亞硨磲(3只)(圖1)。

圖1 4組硨磲外殼
1.2 外殼性狀指標測量
根據NORTON等[16]對硨磲各外殼形態學測量方法, 對所有硨磲樣品的殼長(SL)、殼高(SH)、殼寬(SW)、鉸合線長(HLL)、足絲孔長(BHL)、足絲孔寬(BHW)、殼質量(SM)、左殼厚(LST)、右殼厚(RST)和最大放射肋寬(MRW)等可量性狀以及放射肋數目(NRR)和鉸合齒數目(NHT)等可數性狀進行測量, 如圖2。

圖2 硨磲可量性狀示意圖[16]
1.3 數據處理
1.3.1 描述統計
首先將9個硨磲外殼的可量性狀與殼長相除, 進行標準化處理, 獲得校正值。在SPSS 25.0中, 對9個比例性狀分別進行單因素方差分析(one-way ANOVA),若數據偏正態分布可采用秩和檢驗(Kruskal-Wallis Test)。單因素方差分析前方差齊性檢驗結果>0.05時采用ANOVA描述各因素組間多重比較的差異顯著性,<0.05時, 采用韋奇爾(Welch)的值描述各因素組間差異顯著性水平。
1.3.2 聚類分析
確定使WGSS足夠小的聚類數k, 運用Ward法在R Studio 4.1.1中對4組硨磲的9個比例性狀進行層次聚類分析(Hierarchical Clustering, HC)[17], 距離計算依據歐式距離, 公式如下:

表示兩個個體在維空間的距離, 其中每兩個個體的9個比例性狀均可用坐標(1,2, …,9)和(1,2, …,9)表示, 即計算個體兩兩之間歐式距離矩陣。
1.3.3 主成分分析
在R Studio 4.1.1中對數據進行KMO檢驗和Bartlett球形檢驗, 通過檢驗(KMO檢驗各項MSA≥0.49, 總體MSA = 0.78; Bartlett球形檢驗= 2.06e–50)后進行主成分分析(Principal Component Analysis, PCA)[ 18-21]。我們從9個比例性狀中提取貢獻率最高的3個主成分建立3D立體坐標系可視化4組硨磲的分布差異。
1.3.4 判別分析
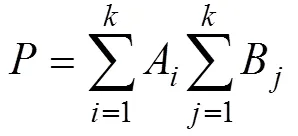
2 結果分析
2.1 4組硨磲外殼性狀描述統計
2.1.1 4組硨磲外殼形態描述
番紅硨磲-1組殼型扁長, 整體呈等腰扇形, 正對殼邊緣觀呈梭形, 放射肋4~5個, 略微突出; 番紅硨磲-2組殼型圓鈍, 整體輪廓為等邊三角形, 正對殼邊緣形狀似元寶, 有4~5個不突出放射肋。長硨磲組個體殼型比番紅硨磲長, 放射肋寬但數目少, 一般為4個, 第五個不明顯, 放射肋上鱗片十分立體, 排列最為緊密。諾亞硨磲組殼型細長, 放射肋數目多且形狀突出, 放射肋上鱗片立體, 排列疏松。
2.1.2 單因素方差分析
單因素方差分析的多重比較見表1, 結果表明番紅硨磲-2組的足絲孔寬以及左右殼厚最大, 與其他3組硨磲存在顯著差異(<0.05); 殼寬、殼質量與足絲孔長方面, 番紅硨磲-1組和番紅硨磲-2組之間存在顯著差異(0.05)。3種硨磲的殼高存在顯著差異(<0.05), 均值大小依次為番紅硨磲>長硨磲>諾亞硨磲; 長硨磲的最大放射肋寬最大, 與另外2種硨磲差異顯著(<0.05); 除鉸合線長、殼質量和放射肋數目外, 諾亞硨磲各項外殼形態指標均最小。

表1 4組硨磲外殼形態指標校正值均值
注: 上標不同英文字母表示組間存在顯著性差異, 相同字母組表示無顯著性差異(<0.05), 2個可數性狀“放射肋數目”與“鉸合齒數目”不進行方差分析
2.2 4組硨磲外殼性狀多元統計分析
2.2.1 層次聚類
如圖3所示, 聚類數為4時(灰色虛線框表示), 番紅硨磲-1組和番紅硨磲-2組聚為一支, 同時長硨磲組和諾亞硨磲組聚為一支, 兩大支最后匯聚為一支。分支長度解釋4組硨磲之間外殼形態聚類距離的遠近。與番紅硨磲-1組支系相比, 番紅硨磲-2組與“長硨磲組+諾亞硨磲組”支的聚類距離更近; 與長硨磲組支系相比, 諾亞硨磲組與“番紅硨磲-1組+番紅硨磲-2組”大支的聚類距離接近。聚類結果表明番紅硨磲-2組與番紅硨磲-1組外殼形態的確存在差異。
2.2.2 主成分分析
以4組硨磲的9個外殼比例性狀進行主成分分析, 主成分與各比例性狀的相關系數和主成分貢獻率見表2。結果顯示: 第一主成分PC1貢獻率為50.47%, 第二主成分PC2貢獻率為13.78%, 第三主成分PC3貢獻率為9.60%, 累積貢獻率為73.85%。主成分特征向量絕對值表明PC1主要反映左右殼厚(左殼厚/殼長和右殼厚/殼長)兩個性狀并僅與最大放射肋寬(最大放射肋寬/殼長)的大小成反比, PC2主要反映殼質量(殼質量/殼長)并與之成反比, PC3主要反映殼高和殼質量(與殼高/殼長呈正相關, 與殼質量/殼長呈負相關)兩個性狀。
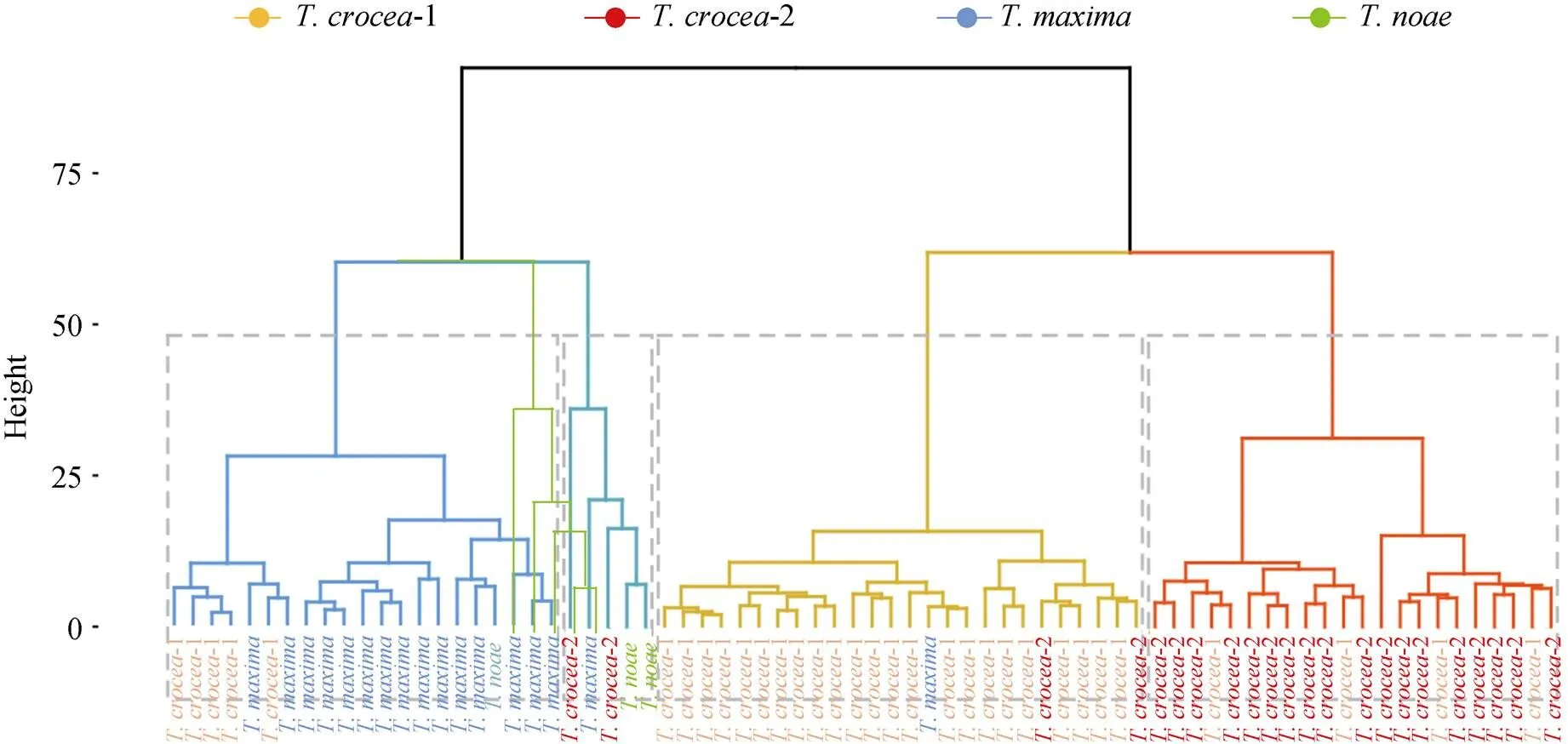
圖3 4組硨磲外殼比例性狀層次聚類系譜圖

表2 4組硨磲9個比例性狀3個主成分特征向量與主成分貢獻率
注: 上標“*”表示主成分反映的主要性狀, “—”表示此性狀與該主成分無相關性
提取的3個主成分雖然不能高度反映所有性狀, 但三者累積貢獻率最高, 最能綜合展示4組硨磲的9個外殼性狀特征。因此, 以PC1、PC2和PC3建立三維立體坐標系如圖4。結果表明: 番紅硨磲-1組和番紅硨磲-2組主成分分布綜合接近, 且左右殼厚(左殼厚/殼長和右殼厚/殼長)最大, 說明番紅硨磲的殼厚度最大; 長硨磲組與諾亞硨磲組的最大放射肋寬(最大放射肋寬/殼長)較兩組番紅硨磲更大, 即長硨磲和諾亞硨磲擁有更寬的放射肋; 諾亞硨磲組殼高/殼長比另外3組的小, 證明諾亞硨磲殼高小, 殼型狹長。
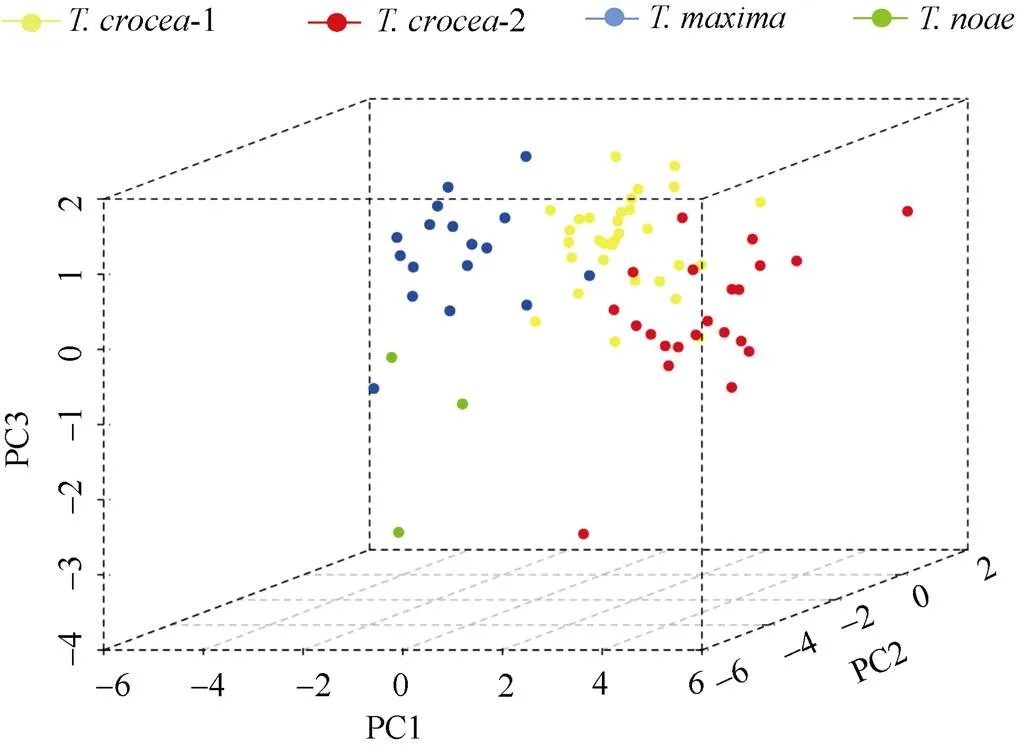
圖4 4組硨磲主成分分析分布散點圖
2.2.3 線性判別分析
Fisher線性判別分析法進行預測分類得到3個線性組合LD1、LD2和LD3, 方差解釋率依次為70.13%, 20.97%和8.90%。3個線性判別函數的公式依次為:
LD1= –0.59201+0.12572–0.36893–0.72624–
0.02185–0.35556–0.10517–0.30378+
0.97969;
LD2=1.09961–0.37712+0.07273+0.70894–
0.21035–0.82536+0.27447–0.89048+
0.64099;
LD3=0.45751–1.02112+0.48733–0.43954–
0.01085+0.25096–0.16547–0.3210–
0.64969;
其中,1~9依次為殼高/殼長、殼寬/殼長等9個外殼比例性狀。以方差解釋率最高的LD1和LD2為軸建立分布坐標系如下圖5。

圖5 4組硨磲的線性判別分析
線性判別結果表明, 在殼寬(殼寬/殼長)和最大放射肋寬(最大放射肋寬/殼長)方面, 番紅硨磲最小, 并且兩組番紅硨磲也存在差異(番紅硨磲-2組較小), 長硨磲與諾亞硨磲兩者表現相似。番紅硨磲的殼高(殼高/殼長)、足絲孔長(足絲孔長/殼長)、左右殼厚(左右殼厚/殼長)和殼質量(殼質量/殼長)在3種硨磲中最大; 值得一提的是番紅硨磲-2組的殼質量(殼質量/殼長)大于番紅硨磲-1組。
3 討論
3.1 4組硨磲形態差異比較分析
2組番紅硨磲單因素方差分析、秩和檢驗和判別分析均表明某些外殼形態指標存在差異, 如殼寬/殼長、足絲孔寬/殼長、殼質量/殼長、左殼厚/殼長和右殼厚/殼長。4組硨磲主成分分析和判別分析結果均支持彼此外殼形態存在差異, 即2組番紅硨磲分布相近, 4組硨磲能夠明顯區分。諾亞硨磲主成分分析PC3方面分布與其他類群差異大, 結合描述性統計分析表明體現在殼質量/殼長(相關系數–0.116)與鉸合線長/殼長(相關系數–0.142)方面, 鉸合線長/殼長可能與閉殼肌形狀有關[23], 殼質量/殼長可能是其殼長與殼高的綜合表現有別于番紅硨磲和長硨磲導致[24]。
本研究對4組小型硨磲的外殼形態學分析發現長硨磲與諾亞硨磲更為相近, 并且兩組番紅硨磲外殼存在顯著差異。然而利用COI和16S基因建立系統發育進化樹結果顯示3種硨磲各自形成分支且番紅和長硨磲親緣關系較近, 且兩組番紅硨磲COI和16S基因序列無差異[25]。一方面, 養殖密度、海水流速、底質和捕食者等諸多環境差異因素均能導致貝類外殼形態變化。如鱗硨磲()在有無捕食者處理下, 其形態發生顯著改變[13]; LUTTIKHUIZEN[26]證明捕食者會選擇性捕食不同外殼形態的荷蘭白櫻蛤(); 在高密度環境下, 降低食物濃度可致貝殼變得更狹窄[27]; 此外, EMERSON[28]對實驗室海水養殖沙蜆()的研究發現水流速度越大, 貝殼外形越圓鈍, 然而美國ORTMANN等[29]在俄亥俄和田納西河對淡水蛤(Naiades)的調查顯示, 存在水流湍急上游的個體殼型不及水流緩慢下游個體圓鈍的現象; HINCH等[30]將沙灘和泥灘珠蚌群體進行交換環境養殖發現二者形態發生改變, 說明轉移底質環境養殖對殼型產生影響。另一方面, 盡管COI與16S基因已被證實能夠較好地區分不同硨磲種類, 但其對同種硨磲的不同群體的區分性卻未見報道[25]。截至目前, 微衛星技術已在水產動物不同種群鑒定中被廣泛應用, 如田鎮[31]利用微衛星標記了7個不同種群文蛤()間的基因差異; ESTOUP等[32]發現地中海和大西洋群體的虹鮭魚()之間存在等位基因差異。對于本研究中發現的兩種形態差異番紅硨磲的具體原因有待進一步驗證。
3.2 統計分析方法在硨磲形態鑒別上的應用
3種多元統計分析方法將中國南海3種小型硨磲進行了有效區分。聚類分析將個體進行初步歸類, 量化類群之間的差異[33], 結果側重表現4組硨磲之間的聚類距離, 但存在個別樣本的錯聚現象, 可能由于層次聚類作為無監督算法無法將個體進行二次聚類。主成分分析3個主成分累計貢獻率僅為73.85%, 表明由本研究9個比例性狀提取出的3個主成分與硨磲品種的相關性不足(<85%), 后續可嘗試利用諸如橢圓傅里葉分析法等對其他更精細的形態指標進行分析[34]。線性判別分析LD1和LD2的方差解釋率高達91.1%, 4組硨磲在9個外殼比例性狀方面的綜合分布差異幾乎可以由這兩個判別函數全面展示, 是3種多元統計分析法中的最優解。層次聚類對4組硨磲的分類結果最為直觀, 主成分分析和判別分析側重展示4組硨磲外殼綜合比例性狀的差異。主成分分析與線性判別分析均使用降維思想(即以9個外殼比例性狀為依據將樣本點投影在具有良好分類特征的低維空間), 但線性判別分析考慮硨磲分組信息, 因此本研究中判別分析效果優于主成分分析。然而即使在已知品種的情況下, 判別分析也不總是最準確的多元統計分析方法, 對于樣本量較少的情況, 主成分分析性能優于線性判別分析[35], 從這個方面可以解釋作者的硨磲樣本量足夠大。
[1] NEO M L, COLETTE C C W, RICHARD D B, et al.Giant clams (Bivalvia: Cardiidae: Tridacninae): A comprehensive update of species and their distribution, current threats and conservation status: an annual review[J]. Oceanography and Marine Biology, 2017, 55: 2-303.
[2] ZHANG Y H, ZHOU Z H, QIN Y P, et al. Phenotypic traits of two boring giant clam () popu-lations and their reciprocal hybrids in the South China Sea[J]. Aquaculture, 2020, 519: 734890.
[3] HERNAWAN U E. Taxonomy of Indonesian giant clams (Cardiidae, Tridacninae)[J]. Biodiversitas, 2012, 13(3): 118.
[4] LIU C S, YANG X Y, SUN Y, et al. Effects of the daily light/dark cycle on photosynthetic performance, oxidative stress and illumination-related genes in boring giant clam[J]. Marine Biology, 2021, 168(5): 71.
[5] LIU C S, LI X B, WU C L, et al. Effects of three light intensities on the survival, growth performance and biochemical composition of two size giant clamsin the Southern China Sea[J]. Aquaculture, 2020, 528: 735548.
[6] SU Y, HUNG J H, LIU L L, et al.(R?-ding, 1798) – a valid giant clam species separated from(R?ding, 1798) by morphological and genetic data[J]. Raffles Bulletin of Zoology, 2014, 62: 124-135.
[7] FAUVELOT C, ZUCCON D, BORSA P, et al. Phylogeographical patterns and a cryptic species provide new insights into Western Indian Ocean giant clams phylogenetic relationships and colonization history[J]. Journal of Biogeography, 2020, 47(5): 1086-1105.
[8] 劉括, 李曉彤, 車宗豪, 等. 貝類種群殼形態性狀多態性研究進展[J]. 海洋科學, 2021, 45(4): 213-221.
LIU Kuo, LI Xiaotong, CHE Zonghao, et al. Current trends in population research on shell morphological polymorphism of mollusks[J]. Marine Sciences, 2021, 45(4): 213-221.
[9] 魏開建, 熊邦喜, 趙小紅, 等. 五種蚌的形態變異與判別分析[J]. 水產學報, 2003, 1: 13-18.
WEI Kaijian, XIONG Bangxi, ZHAO Xiaohong, et al. Morphological variations and discriminant analysis of five freshwater unionid species (Bivalvia: Unionidae)[J]. Journal of Fisheries of China, 2003, 1: 13-18.
[10] 錢榮華, 李家樂, 董志國, 等. 中國五大湖三角帆蚌形態差異分析[J]. 海洋與湖沼, 2003, 34(4): 436-443.
QIAN Ronghua, LI Jiale, DONG Zhiguo, et al. Morphological variations analysis among populations ofin five large lakes of China[J]. Ocea-no-logia et Limnologia Sinica, 2003, 34(4): 436-443.
[11] 張永普, 林志華, 應雪萍. 不同地理種群泥蚶的形態差異與判別分析[J]. 水產學報, 2004, 28(3): 339-342.
ZHANG Yongpu, LIN Zhihua, YING Xueping, et al. Morphological variations and discriminant analysis of different geographical populations of[J]. Journal of Fisheries of China, 2004, 28(3): 339- 342.
[12] 吳楊平, 姚國興, 陳愛華, 等. 文蛤屬2種貝類多變量形態分析及日本文蛤的物種有效性[J]. 水產學報, 2011, 35(9): 1410-1418.
WU Yangping, YAO Guoxing, CHEN Aihua, et al. Multivariate morphometric analysis of two species of, with notes on Japanese’s validity[J]. Journal of Fisheries of China, 2011, 35(9): 1410-1418.
[13] MEI L N, TODD P A. Predator-induced changes in fluted giant clam () shell morpholo-gy[J]. Journal of Experimental Marine Biology & Eco-logy, 2011, 397(1): 21-26.
[14] OTHMAN A S B, GOH G H S, TODD P A. The distribution and status of giant clams (family Tridacnidae)–a short review[J]. Raffles B. Zool., 2010, 58(1): 103-111.
[15] 劉春勝, 吳川良, 李秀保, 等. 三亞近海諾亞硨磲分布特征及底質對其外套膜顏色的影響[J]. 水生生物學報, 2021, 45(3): 645-651.
LIU Chunsheng, WU Chuanliang, LI Xiubao, et al. Distribution and the influence of substrate on its mantle color of giant clamin coastal waters of Sanya[J]. Acta Hydrobiologica Sinica, 2021, 45(3): 645-651.
[16] NORTON J H, JONES G W. The giant clam: an anatomical and histological atlas[R]. Canberra: Australian Centre for Agriculture Research, 1992, 142.
[17] 徐金明, 龔克峰, 魯辰達. 黏性土層位的聚類與判別分析[J]. 地下空間與工程學報, 2009, 5(6): 1138-1144, 1150.
XU Jinming, GONG Kefeng, LU Chenda. Clustering and discriminant analysis of cohesive soils[J]. Chinese Journal of Underground Space and Engineering, 2009, 5(6): 1138-1144, 1150.
[18] 吳姍姍, 張鳳成, 曹可. 基于集對分析和主成分分析的中國沿海省海洋產業競爭力評價[J]. 資源科學, 2014, 36(11): 2386-2391.
WU Shanshan, ZHANG Fengcheng, CAO Ke. Marine industry competitiveness of coastal provinces in China based on set pair analysis and principal component ana-lysis[J]. Resources Science, 2014, 36(11): 2386-2391.
[19] 楊運萍, 郭榮發. 施磷施鈣處理下花生農藝性狀的相關分析及主成分分析[J]. 廣東農業科學, 2014, 41(5): 119-122, 133.
YANG Yunping, GUO Rongfa. Analysis on correlation and principal component of major agronomic traits of peanut in calcium and phosphorus treatment[J]. Guang-dong Agricultural Sciences, 2014, 41(5): 119-122, 133.
[20] ROBBINS K A, BURKHARDT C. Intra-and inter- plat-form renormalization and analysis of microarray data from the NCBI GEO database[M]. Texas: Department of Computer Science, University of Texas at San Antonio, 2007.
[21] HAIR J F, BLACK B, BABIN B J, et al. Multivariate Data Analysis[M]. Beijing: China Machine Press, 2011.
[22] 馬愛軍, 王新安, 孫志賓, 等. 紅鰭東方鲀()三個不同群體的形態差異分析[J]. 海洋與湖沼, 2016, 47(1): 166-172.
MA Aijun, WANG Xinan, SUN Zhibin, et al. Morphological variations among three different populations of[J]. Oceanologia et Limnologia Sinica, 2016, 47(1): 166-172.
[23] BALLA S A, WALKER K F. Shape variation in the Australian freshwater musselIredale (Bivalvia, Hyriidae)[J]. Hydrobiologia, 1991, 220(2): 89-98.
[24] 陳亭君, 何旭盟, 申玉春, 等. 20月齡尖紫蛤殼形態性狀對體質量的影響[J]. 廣東海洋大學學報, 2019, 6: 23-29.
CHEN Tingjun, HE Xumeng, SHEN Yuchun, et al. Effects of morphological traits on body weight of 20- month-old[J]. Journal of Ocean University of Guangdong, 2019, 6: 23-29.
[25] CHAO Q Q, GU Z F, WANG A M, et al. Diversity of three small type giant clams and their associated endosymbiotic Symbiodiniaceae at Hainan and Xisha Islands, South China Sea[J]. Frontiers in Marine Science, 8: 774925.
[26] LUTTIKHUIZEN P C, DRENT J, VAN D W, et al. Spatially structured genetic variation in a broadcast spawning bivalve: quantitative vs. molecular traits[J]. Journal of Evolutionary Biology, 2003, 16(2): 260- 272.
[27] ALUNNO B M, BOURGET E, FRéCHETTE M. Shell allometry and length-mass-density relationship forin an experimental food-regulated situation[J]. Marine Ecology Progress Series, 2001, 219(1): 177-188.
[28] EMERSON C W. Influence of sediment disturbance and water flow on the growth of the soft-shell clam,L.[J]. Canadian Journal of Fisheries and Aquatic Sciences, 1990, 47(9): 1655-1663.
[29] ORTMANN A E. Correlation of shape and station in fresh-water mussels (Naiades)[J]. Proceedings of the American Philosophical Society, 1920, 59(4): 269-312.
[30] HINCH S G, BAILEY R C, GREEN R H. Growth of(Bivalvia: Unionidae) in sand and mud: A reciprocal transplant experiment[J]. Canndian Journal of Fisheries and Aquatic Sciences, 1986, 43(3): 548-552.
[31] 田鎮. 文蛤()7個群體遺傳多樣性分析[D]. 上海: 上海海洋大學, 2021.
TIAN Zhen. Genetic diversity analysis of 7 populations of[D]. Shanghai: Shanghai Ocean University, 2021.
[32] ESTOUP A, PRESA P, KRIEG F, et al. (CT)n and (GT)n microsatellites: a new class of genetic markers forL.(brown trout)[J]. Heredity, 1993, 71(5): 488-496.
[33] 李勤生, 蔡慶華. 東湖異養細菌群落的分類結構和聚類分析[J]. 水生生物學報, 1991, 15(3): 242-254.
LI Qinsheng, CAI Qinghua. The taxonomic structure and cluster analysis of heterotrophic bacterial communities from the Donghu lake[J]. Acta Hydrobiologica Sinica, 1991, 15(3): 242-254.
[34] COSTA C, MENESATTI P, AGUZZI J, et al. External shape differences between sympatric populations of commercial clamsand[J]. Food and Bioprocess Technology, 2010, 3(1): 43-48.
[35] MARTINEZ A M, KAK A C. PCA versus LDA[J]. IEEE Transactions on Pattern Analysis and Machine Intelligence, 2001, 23(2): 228-233.
Morphological variation in shells of three small giant clam species in the South China Sea
CHAO Qi-qi1, YANG Yi1, GU Zhi-feng1, 2, WANG Ai-min1, 2, LóPEZ Violeta3, LIU Chun-sheng1, 2
(1. Ocean College, Hainan University, Haikou 570228, China; 2. Key Laboratory of Tropical Biological Resources of Ministry of Education, Hainan University, Haikou 570228, China; 3. Spanish National Museum of Natural Sciences, Madrid 28006, Spain)
In this study, biostatistical analysis was conducted on the shell morphological traits of three small giant clam species:,and two shell-shape groups ofnative to the South China Sea. Noteworthy variations in shell morphological traits were observed among the four giant clam groups. Analysis of variance showed that there were significant differences in the ratio of SW/SL, BHW/SL, SM/SL, LBT/SL, and RBT/SL between the two groups of(< 0.05). Hierarchical clustering of these four giant clam groups was found to be more effective under the conditions of the Euclidean distance Ward method and the specified cluster number was= 4. Total three principal components were extracted from PCA to establish a scatterplot. The cumulative contribution rate of principal components was 73.85%, and the contribution rates of each principal component were 50.47%, 13.78%, and 9.60%. Three discriminant functions were obtained by LDA, and the variance explanation rates were 70.13%, 20.97%, and 8.90%, respectively. These three multivariate statistical methods could explain the differences in the shell morphology of four giant clam groups from various perspectives. Moreover, results showed that the shell morphology of the twogroups and ofandwere more similar compared with other species.
giant clams; shell morphology; multivariate statistical analysis; South China Sea
Sep. 25, 2021
Q179
A
1000-3096(2022)10-0024-08
10.11759/hykx20210925005
2021-09-25;
2021-12-03
國家自然科學基金項目(42266003); 海南省重點研發項目(ZDYF2019153)
[National Natural Science Foundaton of China, No. 42266003; Key Research and Development Project of Hainan Province, No. ZDYF2019153]
鈔琪琪(1997—), 女, 河南安陽人, 碩士研究生, 主要從事硨磲遺傳多樣性研究, 電話: 18089769063, E-mail: 1962162966@ qq.com; 劉春勝(1984—),通信作者, 電話: 13086002911, E-mail: lcs5113@163.com
(本文編輯: 譚雪靜)
猜你喜歡
裝備制造技術(2019年12期)2019-12-25 03:07:36
模具制造(2019年7期)2019-09-25 07:30:00
暢談(2018年14期)2018-08-27 09:11:16
山東工業技術(2016年15期)2016-12-01 05:30:54
水產養殖(2016年7期)2016-03-28 22:29:47
水產養殖(2016年5期)2016-03-28 16:53:44
應用海洋學學報(2015年4期)2015-11-24 02:49:28
中國收藏(2015年10期)2015-11-05 15:19:22
意林原創版(2015年12期)2015-06-11 01:14:54
電力工程技術(2012年4期)2012-03-25 10:40:35
