日本海神蛤(Panopea japonica)性腺發育生物學零度及有效積溫研究
2022-11-26 06:00:24車宗豪馬倩穎霍忠明閆喜武
海洋科學 2022年10期
趙 雯, 車宗豪, 劉 括, 馬倩穎, 劉 澤, 霍忠明, 閆喜武
日本海神蛤()性腺發育生物學零度及有效積溫研究
趙 雯1, 2, 車宗豪1, 2, 劉 括1, 2, 馬倩穎1, 2, 劉 澤1, 2, 霍忠明1, 2, 閆喜武1, 2
(1. 大連海洋大學水產與生命學院, 遼寧 大連 116023; 2. 遼寧省貝類良種繁育工程技術研究中心, 遼寧 大連 116023)
日本海神蛤(一種象拔蚌)為名貴貝類, 近年來我國已逐漸發展成為日本海神蛤最大的消費市場, 商業潛力較大。為探究日本海神蛤繁殖特性, 于2019年1月至3月, 以采自日本海海域的日本海神蛤為材料, 在大連莊河市貝類育苗場, 通過室內人工升溫促熟方法, 結合精卵排放觀察和數理統計, 求算日本海神蛤生物學零度及有效積溫。結果顯示, 運用二點法、直線回歸法和有效積溫法求得日本海神蛤生物學零度分別為–0.87 ℃、–1.14 ℃、–0.90 ℃, 有效積溫分別為356.21 ℃·d、366.69 ℃·d、357.31 ℃·d。綜合3種方法, 求得日本海神蛤生物學零度及有效積溫的平均值分別為(–0.97±0.14) ℃和(360.07± 5.76) ℃·d。研究發現, 與其他貝類相比, 日本海神蛤性腺發育的生物學零度較低, 這可能與日本海神蛤冷水生活的適應性及繁殖習性有關。在日本海神蛤室內人工苗種繁育過程中, 結合日本海神蛤性腺發育的生物學零度和產卵的有效積溫特點, 通過調控生境水溫, 可以加快性腺發育速度, 達到提早繁育效果。本研究為日本海神蛤在自然海域的苗情預測預報提供科學依據, 對我國北黃海日本海神蛤養殖產業發展和資源修復具有重要的意義。
日本海神蛤; 性腺發育; 人工促熟; 生物學零度; 有效積溫
日本海神蛤()為名貴貝類, 屬軟體動物門(Mollusca)、雙殼綱(Bivalvia)、海螂目(Myoida)、縫棲蛤科(Hiatellidae)、海神蛤屬(), 分布于我國黃海北部、朝鮮半島和日本北海道等海域[1]。在中國市場上, 日本海神蛤與分布在北美西海岸的高雅海神蛤()的商業名稱均為象拔蚌, 其肉質鮮美、營養豐富[2], 深受消費者喜愛, 市場潛力大。目前我國日本海神蛤的自然資源幾近枯竭, 全部依賴進口。20世紀90年代, 中國學者開展了大量的高雅海神蛤()引種試驗[3-4], 在苗種繁育方面取得了突破, 但由于受棲息水溫限制, 關于高雅海神蛤在我國大規模養殖的研究尚無系統報道, 制約了象拔蚌產業發展。日本海神蛤為我國北黃海土著貝類資源[1], 有望在我國遼闊的海域推廣養殖。近年來, 國外學者和國內大連海洋大學霍忠明等[5-10]相繼開展了日本海神蛤繁殖生物學的研究工作, 建立了日本海神蛤人工苗種繁育技術, 為象拔蚌的人工養殖開辟了新途徑。
性腺發育的生物學零度和有效積溫是海洋貝類繁殖生物學研究的重要指標, 有助于了解性腺發育的特點, 為貝類人工苗種繁育提供理論支撐。目前, 關于海洋貝類的性腺發育的生物學零度和有效積溫已有較多報道, 主要包括海灣扇貝()[11-13]、蝦夷扇貝()[14]、菲律賓蛤仔()[15]、西施舌()[16]、縊蟶()[17]、褐云瑪瑙螺()[18]等。畢庶萬等[13]通過使用二點法、最小二乘法、積溫儀計數法對海灣扇貝性腺發育生物學零度和有效積溫進行了研究。劉德經等[16]運用回歸直線法開展了西施舌生物學零度和有效積溫研究。閆喜武[19]通過有效積溫公式=(–), 擬合線性升溫曲線, 結合組織學觀察求算了菲律賓蛤仔性腺發育生物學零度。田斌等[14]采用有效積溫擬合曲線的方法, 計算了蝦夷扇貝性腺發育的生物學零度和有效積溫。Xue等[17]比較了1齡及2齡縊蟶新品種“申浙1號”性腺發育的生物學零度和有效積溫。
目前, 關于海神蛤生物學零度和有效積溫的研究尚未見報道。Marshall等[20]研究了溫度對高雅海神蛤的性腺發育的影響, 發現高雅海神蛤性腺發育的最佳水溫是7 ℃, 繁育水溫是11 ℃, 當水溫升高到15 ℃以上時高雅海神蛤性腺開始退化, 當水溫達到19 ℃時高雅海神蛤的性腺不發育。Aragón-noriega 等[21]對球形海神蛤()的性腺發育及繁殖周期進行了研究, 結果表明, 球形海神蛤在水溫30 ℃時性腺開始逐漸發育, 當水溫逐漸降低至18 ℃時進入繁殖盛期。本研究運用二點法、回歸直線法和有效積溫法3種方法求算日本海神蛤性腺發育的生物學零度及有效積溫, 以期為我國日本海神蛤資源恢復和人工苗種繁育提供科學依據。
1 材料與方法
1.1 親貝的選擇和暫養
于2019年1月26日, 在日本海海域采集殼型規整、無損傷, 水管粗壯且收縮力強的日本海神蛤158個作為親貝低溫運輸至大連莊河貝類育苗場, 在30 m3的水泥池中進行暫養。室內暫養水溫為4.1~4.3 ℃, 鹽度30~31, 餌料按球等邊金藻()和角毛藻()體積比1∶1混合投喂, 日投餌3~4次, 日投餌量約為(1.2~1.8)×104cells/mL, 每天全量換水一次。
1.2 親貝的升溫促熟
分別在兩個水泥池(1號培育池與2號培育池)中按兩種升溫方式進行日本海神蛤室內人工升溫促熟。在親貝人工升溫促熟過程中, 兩個實驗組餌料投喂等日常管理操作保持一致。1號培育池與2號培育池均于1月27日開始升溫, 1號培育池每2日升溫0.5 ℃, 2號培育池每日升溫0.5 ℃, 每日早、中、晚進行3次水溫測量, 根據監測溫度計算日均水溫, 記錄從升溫促熟開始到產卵的總天數。
1.3 統計分析
數據以平均值±標準差表示, 使用SPSS 17.0軟件進行數據分析, 采用 Excel 軟件進行作圖。
2 結果
經過室內人工升溫促熟, 1號培育池在促熟后43 d開始出現產卵排精現象, 平均水溫為(7.41±2.37) ℃, 2號培育池在促熟后38 d出現產卵排精現象, 平均水溫是(8.50±2.45) ℃。兩池人工升溫促熟的日均水溫見表1。根據有效積溫公式:=(–0), 其中為日本海神蛤開始促熟到產卵所需的時間(d);為升溫促熟日均水溫(℃);0為生物學零度;為有效積溫。運用二點法[13]、回歸直線法[16, 22-23]和有效積溫法[14, 19]3種方法求算日本海神蛤性腺發育的生物學零度0及有效積溫。
2.1 二點法
利用兩點法計算, 根據有效積溫相等, 可得(1–0)1==(2–0)2, 其中12分別為1號培育池、2號培育池升溫促熟日均水溫(℃);0為生物學零度;12分別為1號培育池、2號培育池促熟時間(d),為有效積溫。
整理得, 生物學零度:
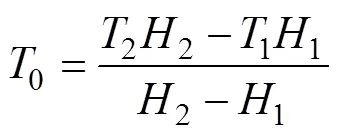
式中1和2分別為7.41 ℃和8.50 ℃;1和2分別為43 d和38 d。結果表明, 日本海神蛤性腺發育生物學零度為–0.874 ℃, 有效積溫為356.212 ℃·d。
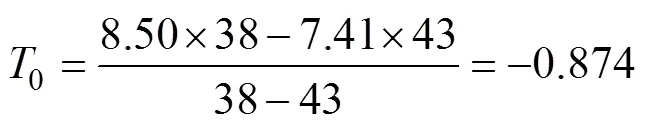
=(7.41+0.874)×43=(8.50+0.874)×38=356.212。
2.2 回歸直線法
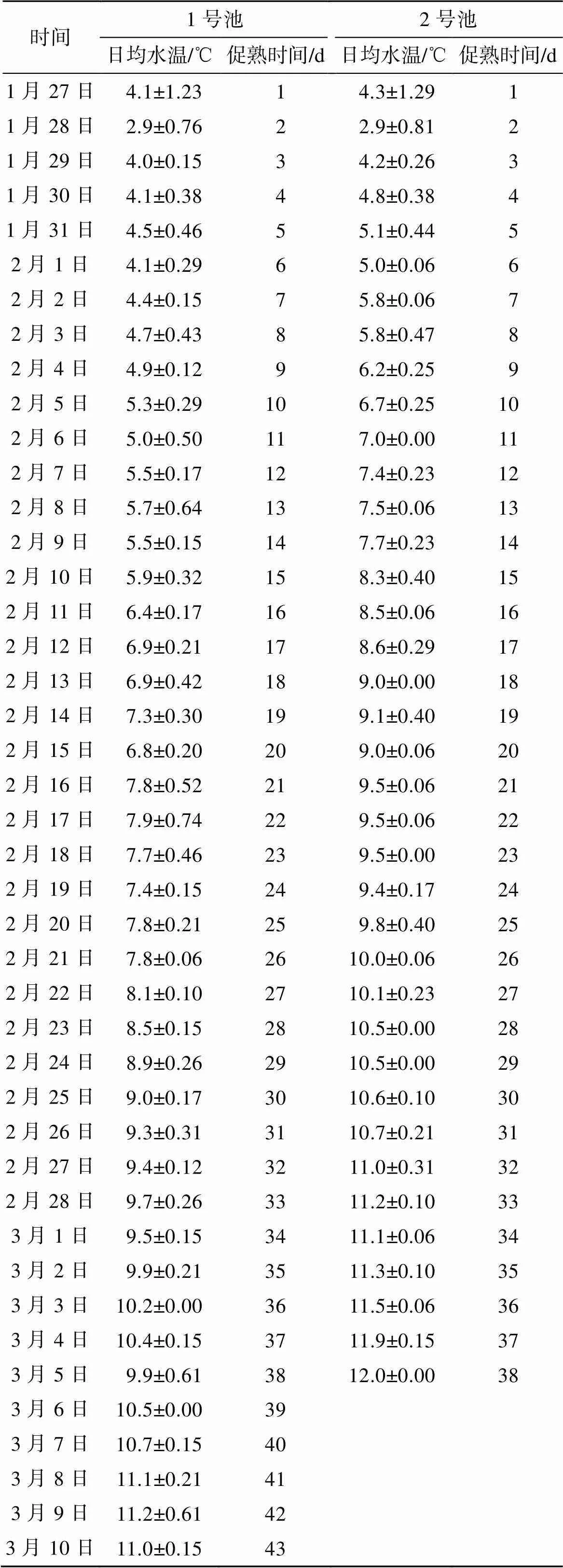
表1 日本海神蛤室內人工升溫促熟日均水溫
式中促熟天數、日均水溫等觀測數據見表2, 將表中數據代入回歸直線法計算公式, 求算生物學零度0和有效積溫值, 結果表明, 日本海神蛤性腺發育生物學零度為–1.14 ℃, 有效積溫為366.69 ℃·d。
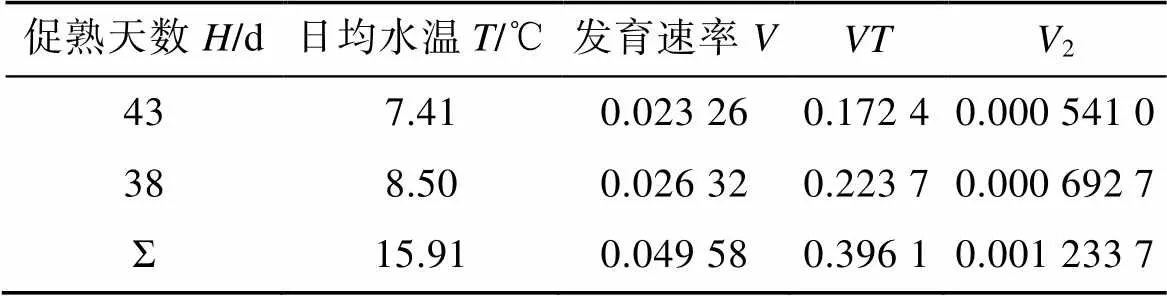
表2 日本海神蛤性腺發育生物學零度與有效積溫計算
2.3 有效積溫法
記錄兩批親貝從開始促熟到自然產卵的日均水溫, 根據積溫公式=(–0)擬合線性升溫曲線, 利用積溫值相同, 即1=2、統計得出生物學零度0及有效積溫。
日本海神蛤人工升溫促熟的兩個培育池擬合水溫變化見圖1、圖2。
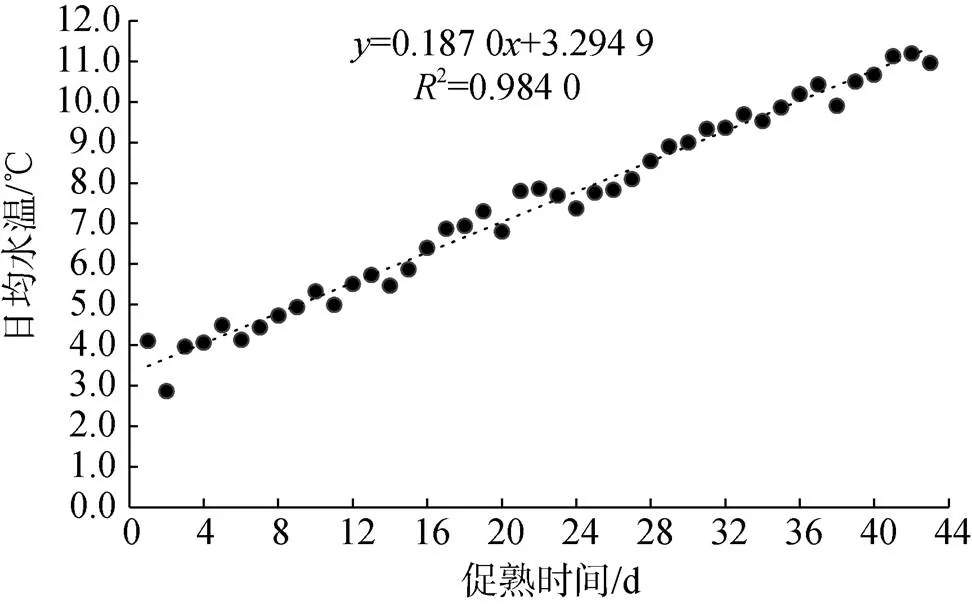
圖1 1號培育池日本海神蛤人工升溫促熟實驗水溫變化
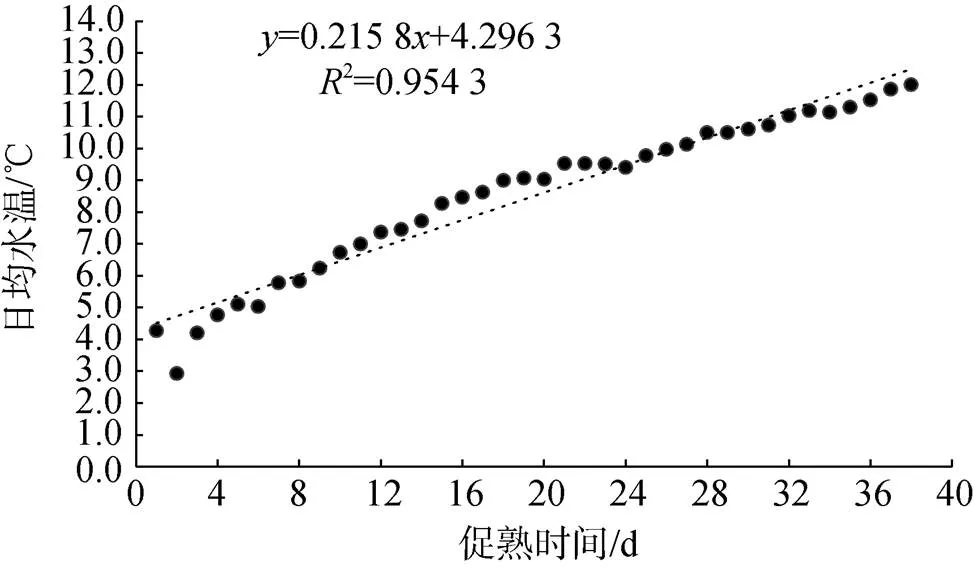
圖2 2號培育池日本海神蛤人工升溫促熟實驗水溫變化
根據日均水溫擬合兩個實驗的升溫曲線分別為L1(圖3a)和L2(圖3b)。
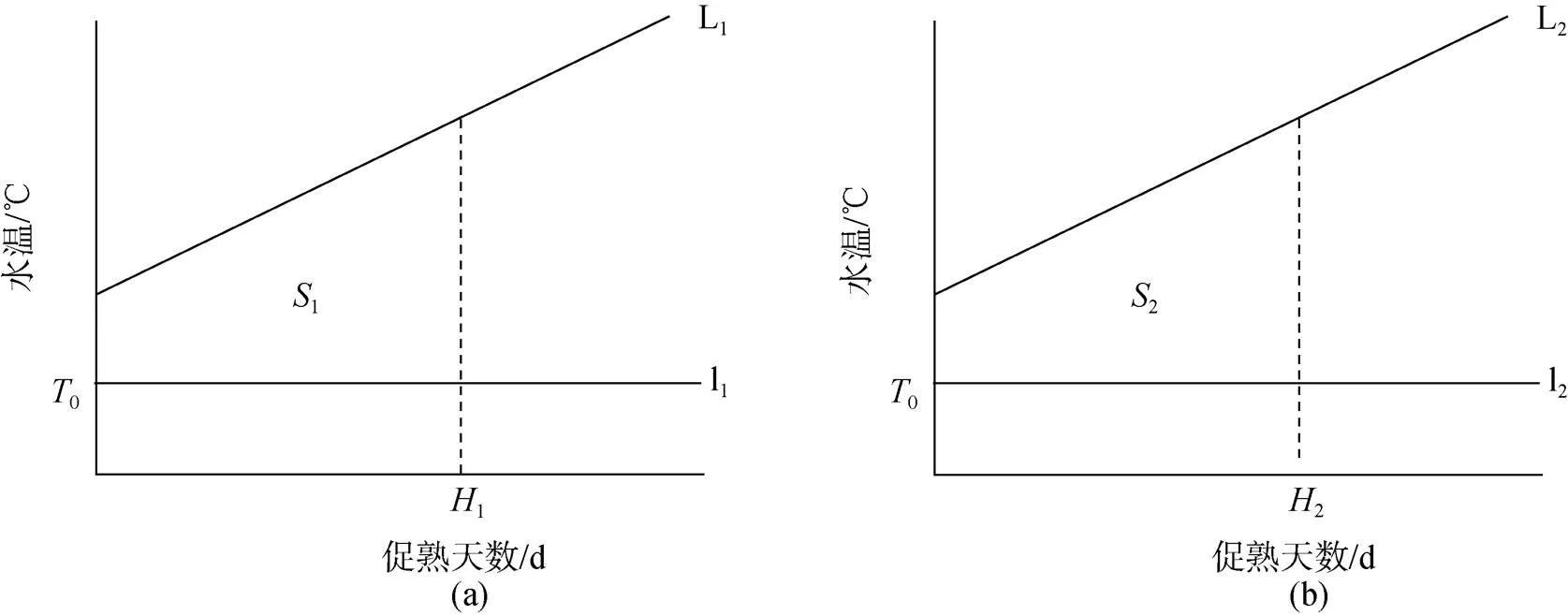
圖3 實驗的積溫圖(實際)
L1:= 0.187 0+ 3.294 92= 0.984 0 (1)
L2:= 0.215 8+ 4.296 32= 0.954 3 (2)
由前兩種方法得知, 日本海神蛤的生物學零度0較低, 親貝入池水溫已高于0, 故實際實驗積溫圖如圖3:
由1=2得,
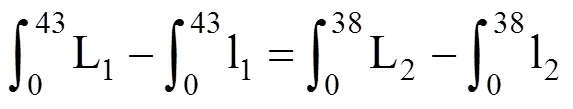
由(3)解得:0=–0.900 96
根據生物學零度–0.900 96 ℃, 按有效積溫公式=(–0)推算, 得出在人工促熟條件下, 日本海神蛤的有效積溫為(357.305±0.09) ℃·d(見表3)。

表3 日本海神蛤有效積溫
根據二點法、直線回歸法和有效積溫法計算獲得的日本海神蛤生物學零度和有效積溫(見表4), 求得日本海神蛤生物學零度及有效積溫平均值分別為(–0.97±0.14) ℃和(360.07±5.76) ℃·d。
3 討論
溫度是影響貝類的性腺發育及繁殖周期的重要因素[24-26], 掌握其性腺發育的生物學零度和產卵的有效積溫, 可通過調節生境溫度的方法, 可以加快性腺發育速度, 達到提早繁育效果, 為貝類室內人工升溫促熟苗種繁育提供科學的理論依據[27]。目前關于貝類的生物學零度的計算方法較多, 如二點法、回歸直線法和有效積溫法。利用二點法計算, 需要樣本數少, 計算方法簡單, 可快速得到數據, 但由于代入數值為日均溫度, 存在精度不足的問題。回歸直線法將發育起點溫度轉化為求算直線回歸系數, 簡化了計算過程, 但是這種方法在計算過程中顛倒了自變量和因變量的關系, 將原來的自變量變成了因變量。二點法及回歸直線法法均為早期方法, 計算較為簡便, 首先在昆蟲等變溫性動物中應用[22, 28], 后被引入貝類等水產動物。閆喜武[19]在計算菲律賓蛤仔時應用了有效積溫法, 根據=(–)擬合線性升溫曲線, 利用有效積溫值進行數理統計得出性腺發育生物學零度, 再通過組織學方法進行驗證。此方法適用于實際生產。擬合線性升溫曲線, 一定程度上減少了實驗溫度不穩定原因造成的影響, 再結合組織學方法進行驗證, 具有較強的說服力。本文采用3種方法計算得出日本海神蛤生物學零度分別為–0.874 ℃、–1.1352 ℃和–0.900 96 ℃, 有效積溫分別為356.212 ℃·d、366.689 ℃·d及357.305 ℃·d, 結果較為相近。綜合3種方法, 求得日本海神蛤的生物學零度平均值為(–0.97±0.14) ℃, 有效積溫平均值為(360.07±5.76) ℃·d。

表4 日本海神蛤生物學零度和有效積溫
本研究表明日本海神蛤性腺發育生物學零度較低, 這可能與日本海神蛤冷水生活環境適應性和繁殖習性有關。在目前已有的報道中, 性腺發育生物學零度低于0 ℃的水產動物較少, 王妍妍等[29]研究表明條斑星鰈()胚胎發育的生物學零度為–0.1 ℃, 分析指出這可能與條斑星鰈為冷溫性魚類, 與其性腺早期在較低水溫開始發育有關。日本海神蛤為冷水性貝類, 成年個體生活適溫為8~ 22 ℃。李瑩[30]通過日本海神蛤周年性腺發育組織切片觀察發現, 幾乎全年都存在性腺發育處于增殖期的雌性個體, 3—9月出現性腺發育處于成熟期雌性個體; 而雄性日本海神蛤全年都存在性腺發育處于成熟期個體; 雌雄日本海神蛤性腺發育具有不同步的特點。本實驗計算求得日本海神蛤性腺發育生物學零度為–0.97 ℃, 與日本海神蛤自然水域的性腺發育實際情況相一致。
4 結論
本研究計算日本海神蛤生物學零度及有效積溫的平均值分別為(–0.97±0.14) ℃和(360.07±5.76) ℃·d。在日本海神蛤室內人工苗種繁育過程中, 結合日本海神蛤性腺發育的生物學零度和產卵的有效積溫特點, 通過調控生境水溫, 可以加快性腺發育速度, 達到提早繁育效果。
[1] 齊鐘彥, 馬繡同. 黃渤海的軟體動物[M]. 北京: 農業出版社, 1989: 228-229.
QI Zhongyan, MA Xiutong. Mollusca of Huanghai and Bohai[M]. Beijing: Agriculture Press, 1989: 228-229.
[2] 李春艷, 閻磊, 王品虹, 等. 日本海神蛤營養成分分析與評價[J]. 營養學報, 2008, 30(1): 113-114, 116.
LI Chunyan, YAN Lei, WANG Pinhong, et al. Analysis of Nutritional Compositions in Geoduck[J]. Acta Nutrimenta Sinica, 2008, 30(1): 113-114, 116.
[3] 魏振華, 魏利平. 象拔蚌引種及人工育苗技術[J]. 齊魯漁業, 2004, 21(8): 4-7.
WEI Zhenhua, WEI Liping. Studies on artificial intro-du-ction and reproduction of the Pacific geoduck[J]. Shandong Fisheries, 2004, 21(8): 4-7.
[4] 劉明坤, 王昌勃, 孔令鋒, 等. 象拔蚌人工育苗技術研究[J]. 海洋科學, 2013, 37(8): 103-106.
LIU Mingkun, WANG Cangbo, KONG Lingfeng, et al. Studies on artificial reproduction of the Pacific geo-duck[J]. Marine Sciences, 2013, 37(8): 103-106.
[5] Nam M M, Chu L, Kim M K, et al. Development and growth in fertilized eggs and larvae of the Japanese geoduck,reared in the laboratory[J]. The Korean Journal of Malacology, 2014, 30(4): 303- 309.
[6] LEE C S, BAIK K K, HONG K E. Ecological Studies on the habitat of geoduck clam,[J]. Journal of Aquaculture, 1998, 11(1): 105-111.
[7] 霍忠明, 趙雯, 肖友翔, 等. 日本海神蛤人工繁殖及早期生長發育[J]. 水產學報, 2021, 45(2): 235-245.
HUO Zhongming, ZHAO Wen, XIAO Youxiang, et al. Artificial breeding, early growth and development of[J]. Journal of Fisheries of China, 2021, 45(2): 235-245.
[8] Huo Z M, Rbbani M G, Cui H, et al. Larval deve-lopment, juvenile survival, and burrowing rate of geo-duck clams () under different pH conditions[J]. Aquaculture International, 2019, 27(5): 1331-1342.
[9] Huo Z M, RBBANI M G, Tian Y, et al. Nitrite tole-ran-ce of larvae and juveniles of geoduck clam[J]. Aquaculture Research, 2018, 49(8): 2913- 2917.
[10] Huo Z M, Guan H W, RBBANI M G, et al. Effects of environmental factors on growth, survival, and metamorphosis of geoduck clam(A. Adams, 1850) larvae[J]. Aquaculture Reports, 2017, 8: 31-38.
[11] 周瑋. 海灣扇貝性腺發育的生物學零度[J]. 水產學報, 1991, 15(1): 82-84.
ZHOU Wei. The biological zero of gonad development of Bay scallop(Lamarck)[J]. Journal of Fisheries of China, 1991, 15(1): 82-84.
[12] 田傳遠, 梁英, 王如才. 海灣扇貝性腺發育的生物學零度[J]. 青島海洋大學學報, 1995(1): 56-58.
TIAN Chuanyuan, LIANG Ying, WANG Rucai. Study on the biological zero of gonad development of Bay scallop Agropecten irradians[J]. Journal of Ocean University Of Qingdao, 1995(1): 56-58.
[13] 畢庶萬, 徐宗發, 于光溥, 等. 海灣扇貝控溫育苗采卵時間的預報方法[J]. 海洋與湖沼, 1996, 27(1): 93-97.
BI Shuwan, XU Zongfa, YU Guangpu, et al. Forecast of the egg-collection time in seed rearing ofin temperature-controlled water[J]. Oceano-lo-gia et Limnologia Sinica, 1996, 27(1): 93-97.
[14] 田斌, 王璐. 蝦夷扇貝性腺發育的生物學零度與有效積溫研究[J]. 中國水產, 2018(5): 93-96.
TIAN Bin, WANG Lu. Study on the biological zero of gonad development of Japanese scallop[J]. China Fisheries, 2018(5): 93-96.
[15] 閆喜武, 張國范, 楊鳳, 等. 菲律賓蛤仔莆田群體與大連群體生物學比較[J]. 生態學報, 2005, 25(12): 3329-3334.
YAN Xiwu, ZHANG Guofan, YANG Feng, et al. Biological comparisons between Putian population and Da-lian population of manila clams[J]. ACTA Ecologica sinica, 2005, 25(12): 3329-3334.
[16] 劉德經, 黃天華, 肖思祺, 等. 西施舌()生殖腺發育生物學零度和有效積溫的初步研究[J]. 特產研究, 2002, 24(1): 33-34.
LIU Dejing, HUANG Tianhua, XIAO Siqi, et al. Studies on the Thermal Sum Beneficial and Biological Zero of Gonad Development of(Spengler)[J]. Special Wild Economic Animal and Plant Research, 2002, 24(1): 33-34.
[17] XUE B, MENG D, LI H, et al. Determining the biological zero for gonadal development razor clams(lamarck 1818) in Zhejiang, China[J]. Aquaculture and Fisheries, 2021, 6(1): 78-88.
[18] 周衛川, 吳宇芬, 蔡金發, 等. 褐云瑪瑙螺發育零點和有效積溫的研究[J]. 福建農業學報, 2001, 16(3): 25-27.
ZHOU Weichuan, WU Yufen, CAI Jinfa, et al. Developmental zero and effective accumulated temperature of giant African snail[J]. Fujian Journal of Agricultural Sciences, 2001, 16(3): 25-27.
[19] 閆喜武. 菲律賓蛤仔養殖生物學、養殖技術與品種選育[D]. 青島: 中國科學院研究生院(海洋研究所), 2005.
YAN Xiwu. The culture biology and technology and selective breeding in Manila clam,[D]. Qingdao: Institute of Oceanology, Chinese Academy of Sciences Qingdao, 2005.
[20] Marshall R, McKinley R S, Pearce C M. Effect of temperature on gonad development of the Pacific geoduck clam(Gould, 1850)[J]. Aquaculture, 2012, 338/341: 264-273.
[21] Aragon-Noriega E A, Chavez-Villalba J, Gribben P E, et al. Morphometric relationships, ga-metogenic development and spawning of the geoduck clam Panopea globosa (Bivalvia: Hiatellidae) in the central of california[J]. Journal of Shellfish Research, 2007, 26(2): 423-431.
[22] 周楠, 王莉萍, 盧南, 等. 高山小毛蟲發育起點溫度和有效積溫的研究[J]. 云南林業科技, 1993(1): 52-56.
ZHOU Nan, WANG Liping, LU Nan, et al. Study on the biological zero point and effective accumulated temperature for development of[J]. Journal of West China Forestry Science, 1993(1): 52-56.
[23] 李爽, 李耕, 潘玉洲, 等. 刺參生殖腺發育的生物學零度和有效積溫研究[J]. 安徽農業科學, 2016, 44(1): 81-82.
LI Shuang, LI Geng, PAN Yuzhou, et al. Study on the biological zero point and effective accumulated temperature for the gonad development of[J]. Journal of Anhui Agriculture, 2016, 44(1): 81-82.
[24] 樓允東, 吳萍. 溫度在水產動物性別控制中的作用[J].上海水產大學學報, 2008, 17(4): 481-485.
LOU Yundong, WU Ping. The role of temperature in sex control of aquatic animals[J]. Journal of Shanghai Fisheries University, 2008, 17(4): 481-485.
[25] 朱星海, 孫紅振, 楊祖晶, 等. 風信標扇貝的性腺發育與繁殖周期規律研究[J]. 中國海洋大學學報(自然科學版), 2019, 49(2): 52-58.
ZHU Xinghai, SUN Hongzhen, YANG Zujing, et al. Studies on gonadal development and the reproductive cycle of[J]. Periodical of Ocean University of China, 2019, 49(2): 52-58.
[26] Thomas B M, Gabriela N, Lorraine M B, et al. Effect of temperature, diet, light, and cultivation density on growth and survival of larval and juvenile white(Bartsch, 1940)[J]. Journal of Shellfish Research, 2016, 35(4): 981-992.
[27] 林志華, 單樂州, 柴雪良, 等. 文蛤的性腺發育和生殖周期[J]. 水產學報, 2004(5): 510-514.
LIN Zhihua, SHAN Lezhou, CHAI Xueliang, et al. Study on the gonad development and reproductive cycle of[J]. Journal of Fisheries of China, 2004(5): 510-514.
[28] 馬夢然, 曹鶴, 申家軒, 等. 云南派姬小蜂的發育起點溫度和有效積溫[J]. 植物保護, 2020, 46(3): 194-197.
MA Mengran, CAO He, SHEN Jiaxuan, et al. Threshold temperature and effective accumulated temperature ofLiao[J]. Plant Protection, 2020, 46(3): 194-197.
[29] 王妍妍, 柳學周, 劉新富, 等. 溫度、鹽度對條斑星鰈胚胎發育的影響[J]. 海洋水產研究, 2008, 29(6): 27-32.
WANG Yanyan, LIU Xuezhou, LIU Xinfu, et al. Effects of temperature and salinity on embryonic development of barfin flounder[J]. Marine Fisheries Research, 2008, 29(6): 27-32.
[30] 李瑩. 日本海神蛤性腺發育、營養成分周年變化及遺傳多樣性研究[D]. 大連: 大連海洋大學, 2019.
LI Ying. Gonad development, annual variation of nutrients, transcriptome analysis and genetic diversity of[D]. Dalian: Dalian Ocean University, 2019.
Biological zero degree and effective accumulated temperature of the geoduck clam
ZHAO Wen1, 2, CHE Zong-hao1, 2, LIU Kuo1, 2, MA Qian-ying1, 2, LIU Ze1, 2, HUO Zhong-ming1, 2, YAN Xi-wu1, 2
(1. College of Fisheries and Life Science, Dalian Ocean University, Dalian 116023, China; 2. Engineering Research Center of Shellfish Culture and Breeding in Liaoning Province, Dalian 116023, China)
The geoduck clam () is a kind of precious shellfish that holds high commercial potential. In recent years, China has rapidly developed to be the largest market for geoduck clams. To explore the reproductive character of, the biological zero degree and effective accumulated temperature of(collected from the Sea of Japan) were studied by the methods of gonad maturity promotion with indoor artificial elevated temperature, spawning observation, and mathematical statistics in Zhuanghe, Dalian, China, from January to March 2019. The results demonstrated that the biological zero degree of gonadal reproduction ofwas –0.87 ℃, –1.14 ℃, –0.90 ℃, and the effective accumulated temperature was 356.21 ℃·d, 366.69 ℃·d, 357.31 ℃·d, respectively, when calculated using the two-point method, linear regression method, and effective accumulated temperature method. To summarize, the mean values of biological zero degree and effective accumulated temperature were (–0.97±0.14) ℃ and (360.07±5.76) ℃·d. The results illustrated that the biological zero degree of gonadal development was lower in geoduck clams than any other shellfish due to the living ability and reproduction characteristics of geoduck clams in cold water. The study found that spawning periods can be accelerated in the process of artificial breeding by adjusting the seawater temperature according to the character of the biological zero degree and effective accumulated temperature of gonadal development; meanwhile, the study also provided the scientific basis for the seeds forecast of geoduck clams in natural seawater environments. The findings make critical contributions to the aquaculture industry and resource restoration in the North Yellow Sea of China.
; gonadal development; artificial promoting maturity; biological zero degree; effective accu-mu-la-ted temperature
Jul. 29, 2021
S961.2
A
1000-3096(2022)10-0078-07
10.11759/hykx20210729001
2021-07-29;
2021-08-26
國家重點研發計劃藍色糧倉科技創新項目(2019YFD0900704, 2018YFD090140); 現代農業產業技術體系專項資金資助(CARS-49); 遼寧省“興遼英才計劃項目”(XLYC1807271); 大連市高層次人才創新支持計劃項目(杰出青年科技人才; 2021RJ09)
[Ministry of Science and Technology of China’s National Key Research and Development Program, No. 2019YFD0900704, 2018YFD0901404; Supported by the earmarked fund for CARS-49; The Liaoning Talent Revitalization Plan, No. XLYC1807271; Outstanding Young Scientific and Technolohical Talents Foundation of Dalian, No. 2021RJ09]
趙雯(1996—), 女, 漢族, 山西人, 碩士研究生, 主要從事貝類遺傳育種與繁殖研究, E-mail: zhaowenme@sina.com; 霍忠明(1983—),通信作者, 男, 漢族, 遼寧人, 教授, 主要從事貝類遺傳育種與繁殖研究, E-mail: huozm@dlou.edu.cn
(本文編輯: 楊 悅)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
現代畜牧科技(2021年4期)2021-07-21 06:13:00
科技傳播(2019年22期)2020-01-14 03:06:54
福建基礎教育研究(2019年1期)2019-09-10 07:22:44
福建基礎教育研究(2019年1期)2019-05-28 08:39:49
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
數學物理學報(2017年2期)2017-06-05 09:12:30
