哺乳動物FSHR基因選擇壓力分析?
2022-11-22 08:12:12王偉萍
生物學(xué)通報 2022年1期
王偉萍 賀 剛 吳 斌,3??
(1 江西省水生生物保護救助中心 江西南昌 330000 2 江西省水產(chǎn)科學(xué)研究所 江西南昌 330000 3 南京師范大學(xué)生命科學(xué)學(xué)院 江蘇南京 210023)
卵泡刺激素(follicle stimulating hormone,F(xiàn)SH)在促進和維持性腺發(fā)育和生殖功能中發(fā)揮重要作用,但其生理作用需通過分布于細胞膜表面的卵泡刺激素受體(follicle-stimulating hormone re‐ceptor,F(xiàn)SHR)介導(dǎo),卵泡刺激素受體是GPCR(Gprotein-coupled receptors)家族的成員[1]。研究發(fā)現(xiàn),F(xiàn)SHR基因與排卵和繁殖力等存在密切關(guān)系[2-3]。而檢測選擇壓力的最大似然法檢驗已被廣泛接受,其中,選擇系數(shù)ω(ω表示非同義/同義置換速率比值)直觀地反映了生物體在密碼子水平上的適應(yīng)性進化趨勢。ω>1、ω=1 和ω<1 分別代表在進化過程中基因承受正選擇、中性選擇及負選擇(凈化選擇)[4]。本研究通過對哺乳動物FSHR基因序列進行選擇壓力分析,分析其適應(yīng)性進化趨勢,為水生哺乳動物,特別是長江江豚(Neophocaena asiaeorientalis)的保護提供參考。
1 材料與方法
1.1FSHR基因序列 基于NCBI(https://www.ncbi.nlm.nih.gov/)上公布的基因序列,選取33 種哺乳動物(11 種鯨豚類、11 種靈長類、11 種其他類),分別代表一般1 胎1 仔的水生類群、一般1 胎1 仔的陸生類群、一般1 胎多仔的典型類群(表1)。
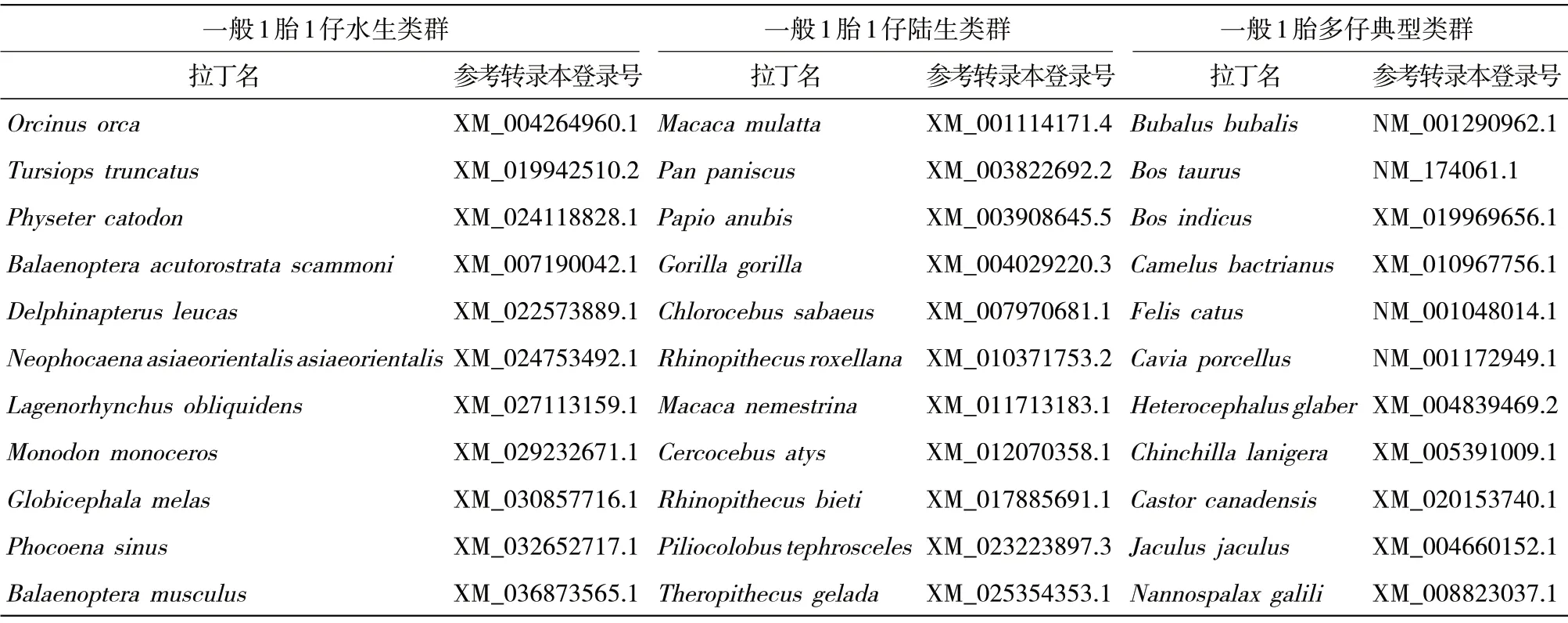
表1 FSHR 基因轉(zhuǎn)錄本信息
1.2 方法 首先,通過FasParser2.10.0 軟件[5],獲得FSHR基因轉(zhuǎn)錄本最長開放閱讀框序列(open reading frame,ORF),隨后通過MEGA X[6]將上述序列對齊。采用DAMBE6.4 軟件[7]對FSHR基因轉(zhuǎn)錄本最長ORF 序列進行飽和度檢測,使用MEGA X中的ML 方法構(gòu)建系統(tǒng)發(fā)育樹,并運用PhyloSuite v1.2.2[8]為Paml 程序運行提供所需文件的格式轉(zhuǎn)換[4]。
其次,應(yīng)用Paml 軟件codeml 模塊中的4 種不同的模型對FSHR基因進行選擇壓力分析。本研究采用位點模型中的M0:假設(shè)所有位點具有相同的ω值;M1:假設(shè)存在2 類位點,保守位點0<ω<1,中性進化位點ω=1,并且估算這2 類位點的比率(p0、p1);M2:假設(shè)存在3 類位點,保守位點、中性進化位點、正選擇位點ω>1,并且估算這3 類位點的比率(p0、p1、p2);M3:離散模型,假設(shè)所有位點的ω值呈離散分布;M7:假設(shè)所有位點0<ω<1 且呈現(xiàn)beta 分布;M8:在M7 模型的基礎(chǔ)上,增加一類正選擇位點(ω>1)。通過M1/M2、M7/M8 重建似然比檢驗LRT,使用M2 和M8 鑒別受到正選擇的位點,分析各位點承受正選擇的壓力位點。
最后,用枝模型分析水生哺乳動物的FSHR基因是否存在加速進化。枝模型假設(shè)系統(tǒng)發(fā)育樹上的不同枝之間有不同的選擇壓(即不同的ω值),以檢測不同種分枝的正選擇壓。這里采用枝標(biāo)簽分3 種情況進行標(biāo)記:1)鯨豚類動物為前景枝;2)靈長類動物為前景枝;3)其他陸生動物為前景枝,均分別用M0 和M2 模型進行相關(guān)分析。
2 結(jié)果與分析
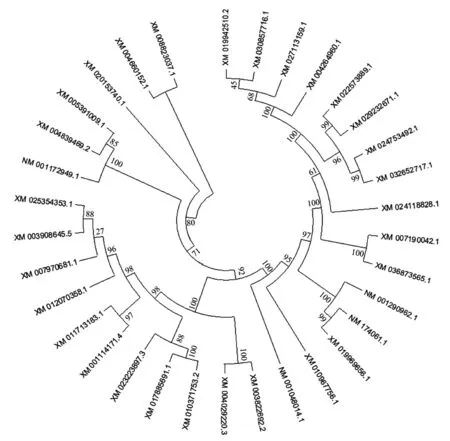
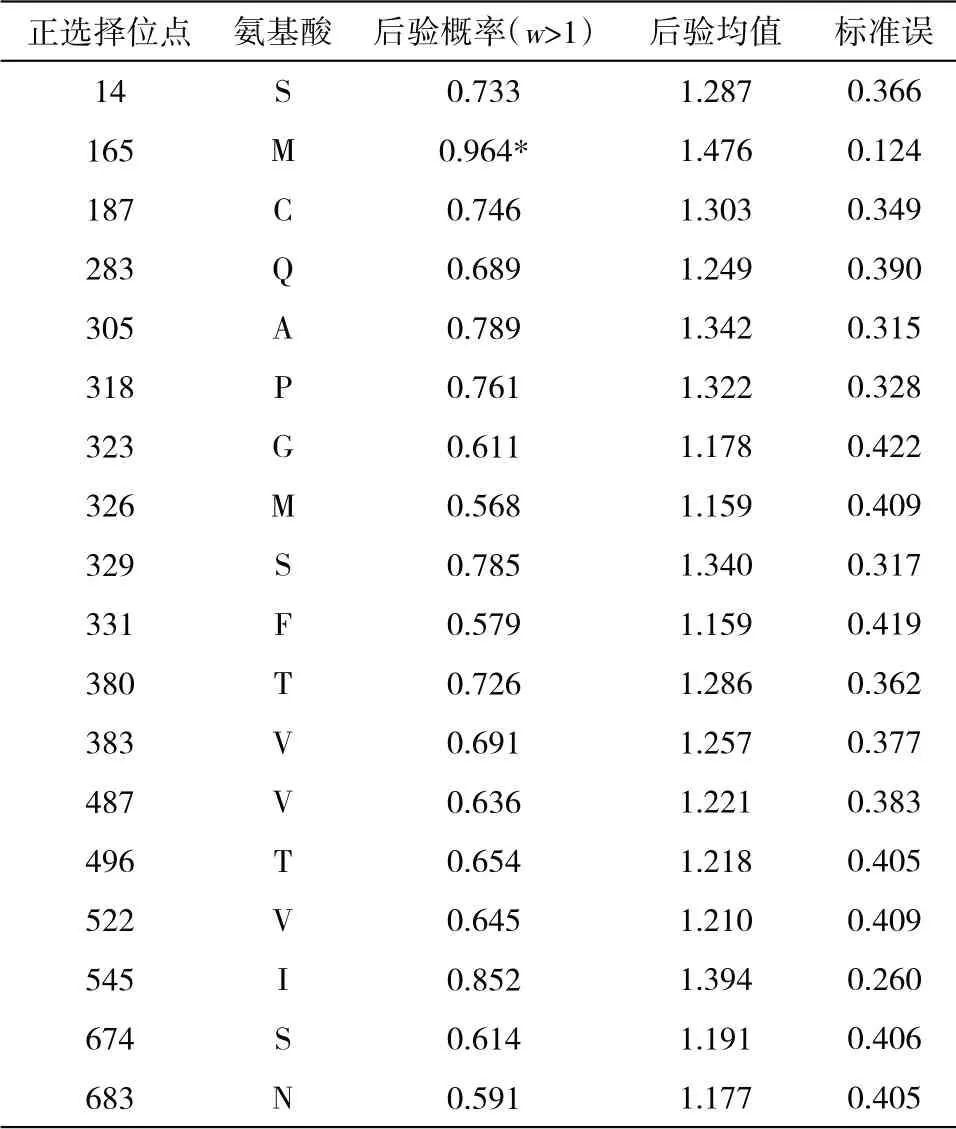
2.1 核苷酸的序列飽和度檢測結(jié)果 采用DAMBE6.4軟件對FSHR基因轉(zhuǎn)錄本最長ORF 序列進行飽和度檢測,均滿足條件P=0.0000,ISS 圖1 FSHR 基因轉(zhuǎn)錄本最長ORF 序列飽和度分析 2.2 系統(tǒng)發(fā)育樹的構(gòu)建 本研究運用MEGAX 中的ML 方法構(gòu)建FSHR基因轉(zhuǎn)錄本最長ORF 序列系統(tǒng)發(fā)育樹(圖2)。構(gòu)建的系統(tǒng)發(fā)育樹拓撲結(jié)構(gòu)與傳統(tǒng)分類基本一致,且bootstrap 值也基本一致,大多數(shù)在90 以上,極少數(shù)低于70,推測FSHR基因進化速率適中,基本形成了比較穩(wěn)定的功能。 圖2 FSHR 基因轉(zhuǎn)錄本最長ORF 序列構(gòu)建的系統(tǒng)發(fā)育樹 2.3 位點模型 比較M0 和M3、M7 和M8,2ΔlnL分別為306.07、20.30,通過χ2檢測,差異極顯著。但比較M1 和M2,2ΔlnL為0,通過χ2檢測,差異不顯著。M3 模型結(jié)果表明,p0、p1 和p2 分別為0.74904、0.25096 和0.00000,對應(yīng)的dN/dS(w)為0.04768、0.66855 和25.36134。M1 模型結(jié)果表明,p0 和p1 分別為0.83578 和0.16422,對應(yīng)的dN/dS(w)為0.07879 和1.00000。M8 模型結(jié)果表明,以XM_008823037.1 為參考序列,F(xiàn)SHR基因轉(zhuǎn)錄本最長ORF 序列正選擇位點有19 個,但只有aa 165 M 一個位點的P>95%,表明FSHR基因所承受的正選擇壓力相對較少,選擇壓力分析結(jié)果見表2。 表2 FSHR 基因正選擇位點結(jié)果 2.4 枝模型 枝模型檢驗結(jié)果顯示,靈長類和其他陸生動物為前景枝,比較M0 和M2,2ΔlnL分別為1.22 和1.45,通過χ2檢測,差異均不顯著。而鯨豚類動物為前景枝,比較M0 和M2,2ΔlnL為5.60,通過χ2檢測,差異顯著。即M2 模型優(yōu)于M0 模型,不同的枝有不同的選擇壓力,且二者均為0<ω<1,表明鯨豚類動物FSHR基因存在加速進化,結(jié)果見表3。 表3 枝模型的似然比檢驗和種系選擇壓結(jié)果 動物對環(huán)境的適應(yīng)及適應(yīng)性進化一直是研究熱點問題,高度保守的繁殖相關(guān)基因的進化系統(tǒng)研究能簡便有效地揭示出其潛在的生物學(xué)功能。例如,鼬科、浣熊科、小熊貓科和臭鼬科均出現(xiàn)胰液核糖核酸酶基因復(fù)制,且檢測到正選擇作用和適應(yīng)性位點,同時結(jié)合RNA 組織表達等實驗結(jié)果,表明該基因不僅有食性相關(guān)的功能,可能還有其他功能[9]。動物采取不同的繁殖策略以適應(yīng)所處環(huán)境,通常由行為及內(nèi)分泌共同調(diào)節(jié)。有研究表明,同一單元內(nèi)雌性川金絲猴(Rhinopithecusroxellana)可能通過季節(jié)層面地錯開生殖狀態(tài)、懷孕機會及邀配行為的方式以減緩生殖競爭并適應(yīng)環(huán)境的季節(jié)性變化[10]。有研究表明,F(xiàn)SHR基因含量的高低可能與哺乳動物的繁殖力之間存在一定相關(guān)性[3],F(xiàn)SHR基因正常表達是卵巢獲得促性腺激素反應(yīng)能力的關(guān)鍵,受體的缺失會引起卵泡不能正常發(fā)育,嚴(yán)重的則會引起繁殖力降低甚至不孕[11]。通過對33 種哺乳動物的FSHR基因的選擇壓力檢測,發(fā)現(xiàn)M3 模型中p2 為0,表明FSHR基因主要承受的選擇壓力是凈化選擇。Kamath和Getz[12]研究指出,在長期的進化過程中選擇壓力不可能一直效應(yīng)于整個基因中,而是作用于某些特殊效用位點上,例如,某個基因編碼區(qū)域的特定位點。因此,F(xiàn)SHR基因的凈化選擇可能主要是FSHR基因?qū)Σ溉閯游镏陵P(guān)重要,較保守。但在FSHR基因也檢測出多個正選擇壓力位點,選擇信號是選擇作用遺留在基因上的印跡。任何一種生物想要生存下去,都必須不斷地進行繁殖,保證自己所屬物種的數(shù)量,繁殖是動物實現(xiàn)自身適合度的方法[13]。這可能是為了應(yīng)對不同的環(huán)境及不同的婚配制度帶來的多重挑戰(zhàn),保證物種的可持續(xù)性發(fā)展,動物通常采用不同的繁殖策略與行為策略以提高自身的交配機會、成功率及后代的存活率[14]而發(fā)生正向選擇突變。對鼠類研究指出,無論是依賴光周期的天算式繁殖,還是綜合實際水熱和食物條件的鼠算式繁殖,都顯示出鼠繁殖策略的靈活性。鼠對環(huán)境調(diào)節(jié)的有效把握和相對較快的傳代速度使其更有可能演化出新的適應(yīng)策略[15]。水生哺乳動物,例如,長江江豚無論在野外還是在人工環(huán)境下均無產(chǎn)雙胞胎的記錄,長江江豚的交配可能發(fā)生在一年中的大多數(shù)月份,并可能有一個以上的交配高峰,撫幼行為比較常見[16],因此,推測鯨豚類FSHR基因的加速進化可能有助于其實現(xiàn)繁殖適應(yīng)。鯨豚類作為物種演化歷史上一類非常重要的次生性水生哺乳動物,成功地適應(yīng)了水生環(huán)境并且快速地輻射進入到全球的各水域。為了適應(yīng)水生生境,鯨類外部形態(tài)和生理、生態(tài)產(chǎn)生了一系列的適應(yīng)性改變,在表型性狀上發(fā)生了明顯變化,特別是在繁殖、生長等性狀上差異較大,但由進化所導(dǎo)致繁殖適應(yīng)的遺傳機制尚不清楚,缺乏系統(tǒng)研究。一個種群繁殖力的強弱直接影響該種群以后的發(fā)展前途及最終命運[17],因此,對以長江江豚為代表的鯨豚類動物繁殖適應(yīng)策略的深入研究有利于科學(xué)開展相關(guān)保護工作。

3 討論
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
