精氨酸水平對梅花鹿仔鹿瘤胃發酵參數和菌群結構的影響
2022-10-24 07:42:46范玉潔司華哲王曉旭楊乾龍張新宇鐘偉王凱英
草業學報 2022年10期
范玉潔,司華哲,王曉旭,楊乾龍,張新宇,鐘偉,王凱英
(中國農業科學院特產研究所,特種經濟動物分子生物學國家重點實驗室,吉林長春 130112)
幼齡反芻動物斷奶后因固體飼料的攝入其消化方式開始由非反芻向反芻轉變,瘤胃發酵功能和微生物區系不斷完善,瘤胃內環境逐漸趨于穩定[1]。大量研究表明[2-5],通過調整幼齡反芻動物飼糧結構,可調控瘤胃發育、完善瘤胃微生物群落和微生態系統,促進反芻動物健康生長。離乳期是梅花鹿生長發育的關鍵時期,仔鹿身體各器官功能及各系統構建均不完善,可塑性極強。因此,調整仔鹿飼糧結構,會對梅花鹿的生長發育以及后續鹿茸生產產生直接影響。精氨酸(arginine,Arg)作為幼齡哺乳動物的條件性必需氨基酸,是機體合成蛋白質最重要的氮源,精氨酸對各種營養物質的代謝起著重要的協同平衡作用,進而調控動物的生長發育[6-7]。目前精氨酸在營養方面的研究主要集中于單胃動物,在反芻動物尤其是幼齡反芻動物上的研究則相對較少。本研究以離乳期梅花鹿為試驗對象,在其飼糧中添加不同水平的精氨酸,調整基礎飼糧結構,探究精氨酸水平對離乳期梅花鹿瘤胃發酵參數和微生物菌群結構的影響,為精氨酸在仔鹿飼糧中的合理利用提供理論依據。
1 材料與方法
1.1 試驗設計
選取3月齡健康離乳雄性梅花鹿仔鹿12只,隨機分為3組,每組4只,各組之間體重差異不顯著(P>0.05)。仔鹿飼喂蛋白質水平為12.28%的低蛋白質飼糧,精氨酸水平分別為0.80%(A組)、1.08%(B組)、1.26%(C組)。按照各組飼糧精粗比相同,賴氨酸和蛋氨酸含量一致的原則,試驗飼糧以玉米(Zea mays)、豆粕、酒糟蛋白、玉米胚芽、苜蓿(Medicago sativa)草粉、糖蜜、食鹽、預混料等為原料,按不同比例配制成顆粒化全混合日糧(total mixed rations,TMR)。試驗飼糧組成及營養水平見表1,試驗飼糧氨基酸含量見表2。試驗于2019年8月27日至10月10日在中國農業科學院特產研究所茸鹿試驗基地進行。每天06:30和14:30分2次定量飼喂,自由飲水。試驗期為50 d,其中預試期為15 d,正試期為35 d。
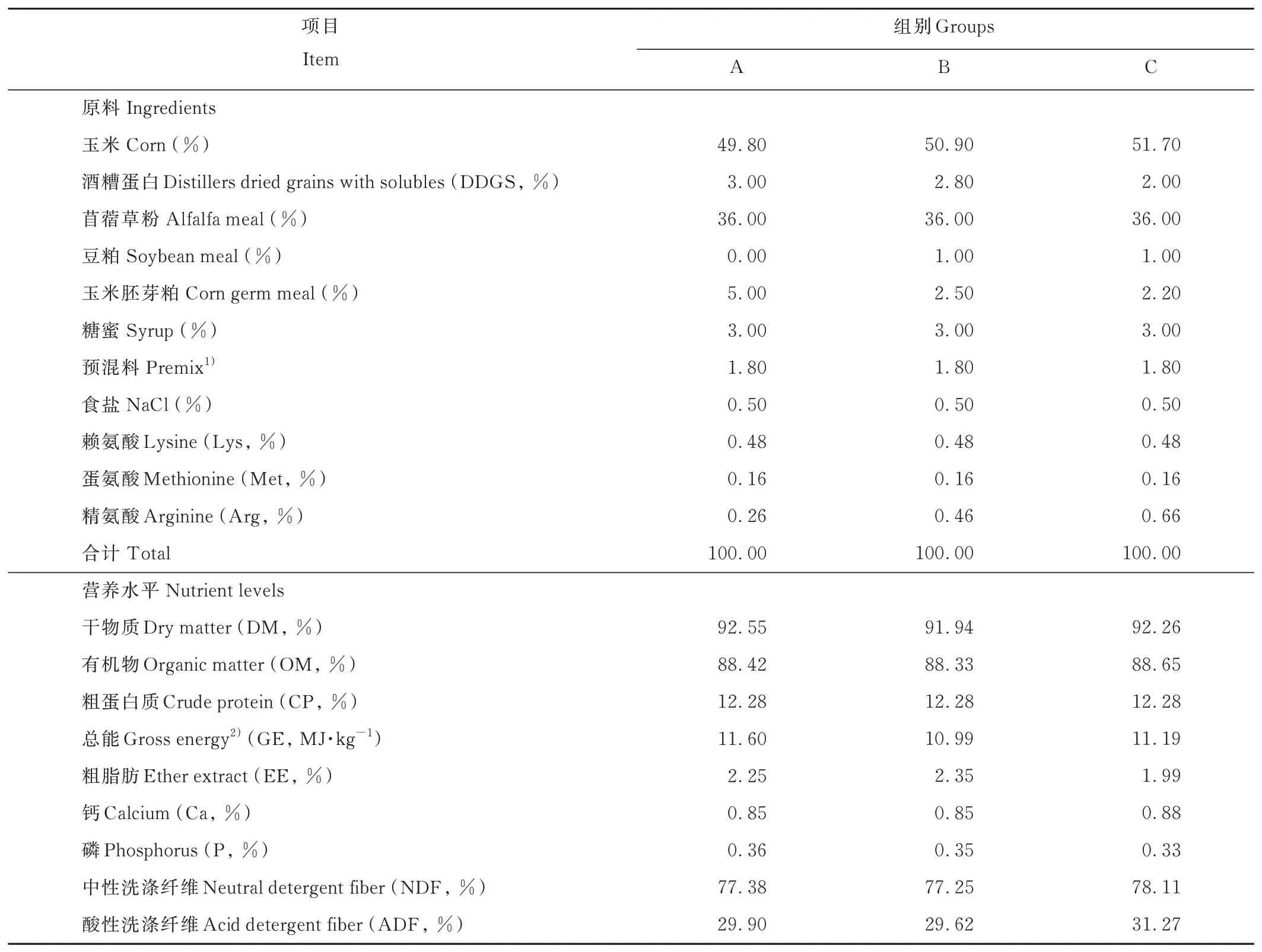
表1 試驗飼糧組成及營養水平(風干基礎)Table 1 Composition and nutrient levels of experiment diets(air-dry basis)
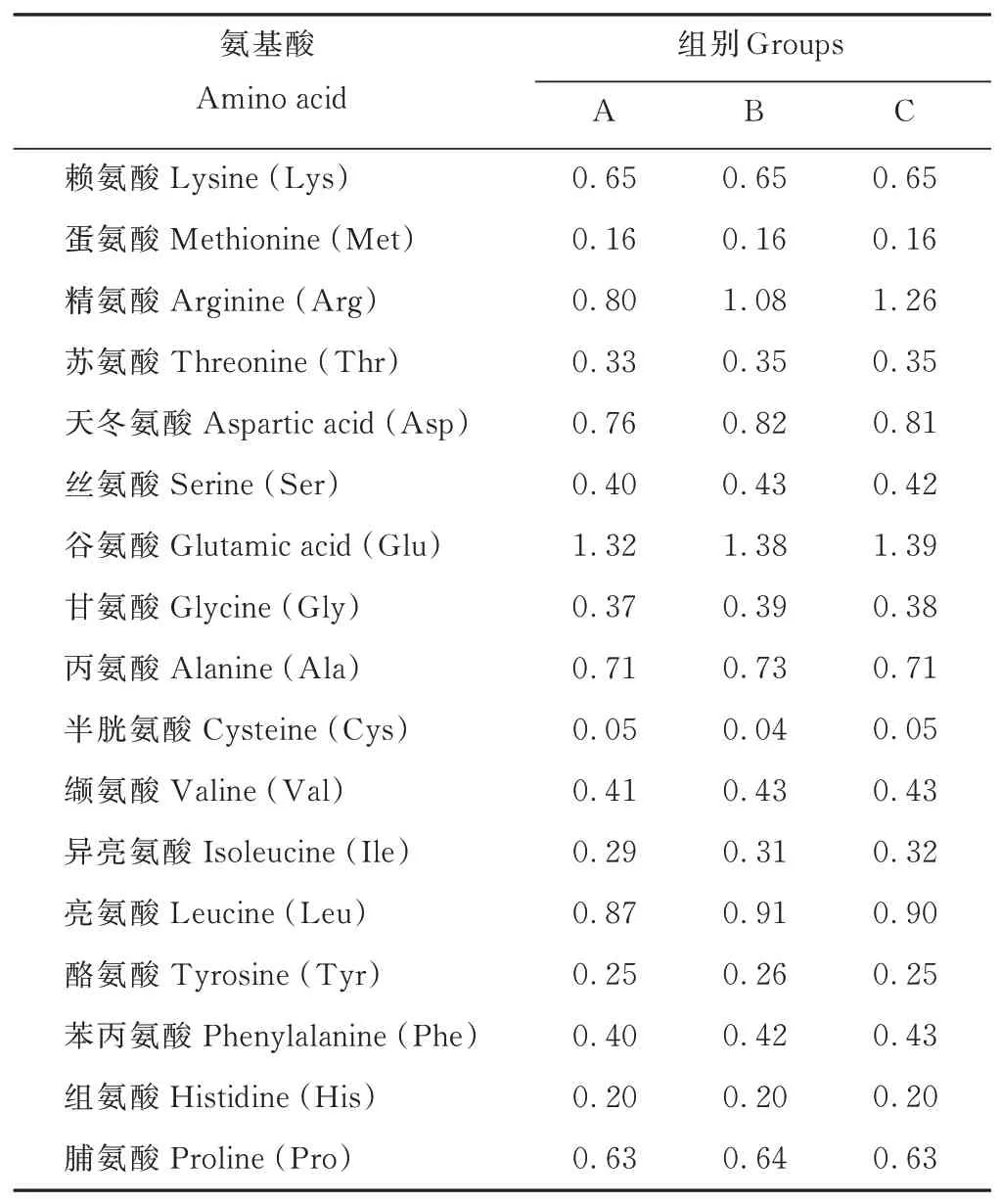
表2 試驗飼糧氨基酸含量(風干基礎)Table 2 Amino acid contents of experiment diets(air-dry basis,%)
1.2 樣品采集
試驗第50天,晨飼前麻醉仔鹿,使用瘤胃管通過口腔抽取瘤胃液20 mL,分裝在凍存管中,立即投入液氮,帶回實驗室將瘤胃液保存于-80℃,一部分用于測定瘤胃發酵參數,另一部分用于提取DNA,測定瘤胃細菌的多樣性。
1.3 指標測定及方法
1.3.1瘤胃發酵參數的測定 瘤胃液pH值采用PHS-3C型pH計(江蘇)進行測定;揮發性脂肪酸(volatile fatty acid,VFA)濃度的測定參考Li等[8]的方法,采用安捷倫6890氣相色譜儀(美國)進行測定,主要測定瘤胃液中乙酸(acetic acid,ACE)、丙酸(propionic acid,PRO)、丁酸(butyric acid,BUTY)、異丁酸(isobutyric acid,ISOB)、異戊酸(isovaleric acid,ISOV)和戊酸(valeric acid,VAL)濃度,并計算乙酸/丙酸(乙丙比)和總揮發性脂肪酸(total volatile fatty acid,TVFA)濃度;氨態氮(NH3-N)濃度的測定參考馮宗慈等[9]改進的比色法。
1.3.2瘤胃細菌多樣性分析 將采集到的瘤胃液送至天津諾禾致源生物信息科技有限公司進行16S rDNA測序。方法如下:采用CTAB或SDS方法對樣本的基因組DNA進行提取,之后利用瓊脂糖凝膠電泳檢測DNA的純度和濃度,取適量的樣本DNA于離心管中,使用無菌水稀釋樣本至1 ng·μL-1。以稀釋后的基因組DNA為模板,根據測序區域的選擇,使用帶Barcode的特異引物,對應區域為16S V4區引物(515F和806R),選擇New England Biolabs公司的Phusion? High-Fidelity PCR Master Mix with GC Buffer和高效高保真酶進行PCR,確保擴增效率和準確性。PCR產物使用2%濃度的瓊脂糖凝膠進行電泳檢測并進行PCR產物等濃度混樣,充分混勻后使用1×TAE的瓊脂糖凝膠電泳進行純化,選擇并回收主帶大小在400~450 bp的序列作為目標條帶,使用膠回收試劑盒(qiagen公司)進行回 收。使 用TruSeq? DNA PCR-Free Sample Preparation Kit建庫試劑盒進行文庫構建,構建好的文庫經過Qubit和Q-PCR定量,文庫合格后,使用NovaSeq 6000進行上機測序。
測序得到的原始數據(raw data)進行拼接、過濾,得到有效數據(clean data)。利用Uparse軟件(Uparse v7.0.1001,http://www.drive5.com/uparse/)對所有樣本的全部有效數據進行聚類,默認以97%的一致性(identity)將 序 列 聚 類 成 為OTUs(operational taxonomic units分類單元),篩選出OTUs的代表序列,進行物種注釋分析(設定閾值為0.8~1.0)。對OTUs進行Alpha和Beta多樣性指數組間差異分析,使 用Qiime軟 件(Version 1.9.1)計 算ACE,Chao1,Shannon,Simpson,Goods-coverage指 數;計 算Weighted Unifrac距離、構建非加權組平均聚類分析(unweighted pair-group method with arithmetic means,UPGMA)樣本聚類樹。使用R軟件(Version 2.15.3)繪制主坐標分析圖(principal co-ordinates analysis,PCoA),通過Amova分析來檢驗不同組間差異顯著性。
1.4 數據統計與分析
運用Excel 2019進行試驗數據初步整理后,采用SAS 9.4軟件的ANOVA進行單因素方差分析,采用Duncan氏法進行多重比較分析組間差異顯著性,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結果與分析
2.1 精氨酸水平對梅花鹿仔鹿瘤胃發酵參數的影響
由表3可知,瘤胃pH隨著精氨酸水平的升高而逐漸降低,B組瘤胃液pH與A、C組之間無顯著差異(P>0.05),A組顯著高于C組(P<0.05);C組氨態氮含量極顯著低于B組(P<0.01),顯著低于A組(P<0.05),A、B組之間無顯著差異(P>0.05);精氨酸水平對總揮發性脂肪酸、乙酸、異丁酸、丁酸、戊酸含量和乙丙比產生了極顯著或顯著影響(P<0.01或P<0.05),對丙酸、異戊酸無顯著影響(P>0.05)。其中,A組乙酸、丁酸含量極顯著高于B、C組(P<0.01),總揮發性脂肪酸含量顯著高于B、C組(P<0.05),B、C組之間無顯著差異(P>0.05);B組異丁酸含量極顯著低于A組(P<0.01),顯著低于C組(P<0.05),A、C組之間無顯著差異(P>0.05);A、B組戊酸含量顯著高于C組(P<0.05);B組的乙丙比極顯著低于A、C組(P<0.01),而A組顯著高于C組(P<0.05)。
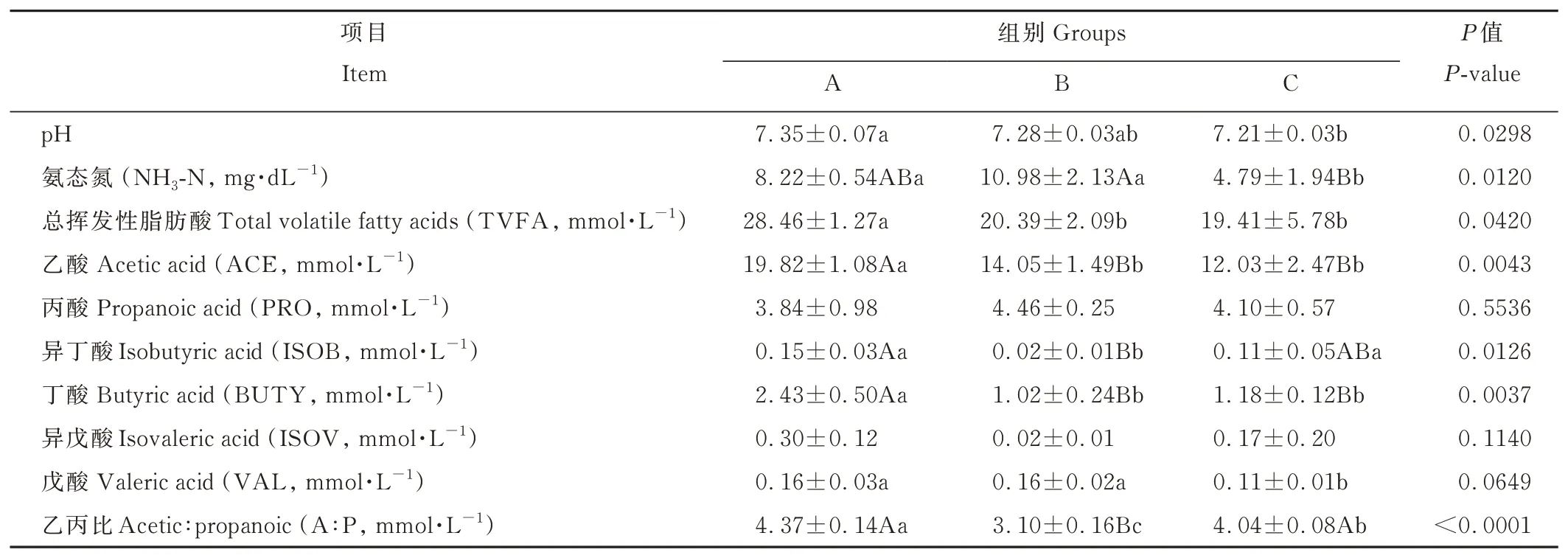
表3 不同精氨酸水平對離乳期梅花鹿瘤胃發酵參數的影響Table 3 Effects of arginine levels on rumen fermentation parameters of weaning sika deer
2.2 精氨酸水平對細菌豐富度和多樣性的影響
基于Illumina NovaSeq測序平臺進行16S rDNA測序,共獲得418397個有效序列,默認以97%的一致性將有效序列進行聚類,如圖1顯示,共產生2098個OTU,其中1021個是3組共有,占總OTU數量的48.67%,A組特有的OTU數量為245個,B組特有的OTU數量為207個,C組特有 的OTU數量為202個。
基于OTU聚類分析結果,進行Alpha多樣性分析。由表4可知,3組ACE和Chao1指數變化趨勢一致,Simpson指數和Shannon指數變化趨勢相反。其中,B組ACE指數顯著高于C組(P<0.05),而A組的ACE指數與B、C組之間均無顯著差異(P>0.05);B組的Chao1指數顯著高于A、C組(P<0.05),A、C組之間無顯著差異(P>0.05)。B組Simpson指數極顯著高于A、C組(P<0.01),Shannon指數極顯著低于A、C組(P<0.01),而A、C組之間無顯著差異(P>0.05)。3組 的 覆 蓋 率(Goods_coverage)均 為99%(P>0.05)。
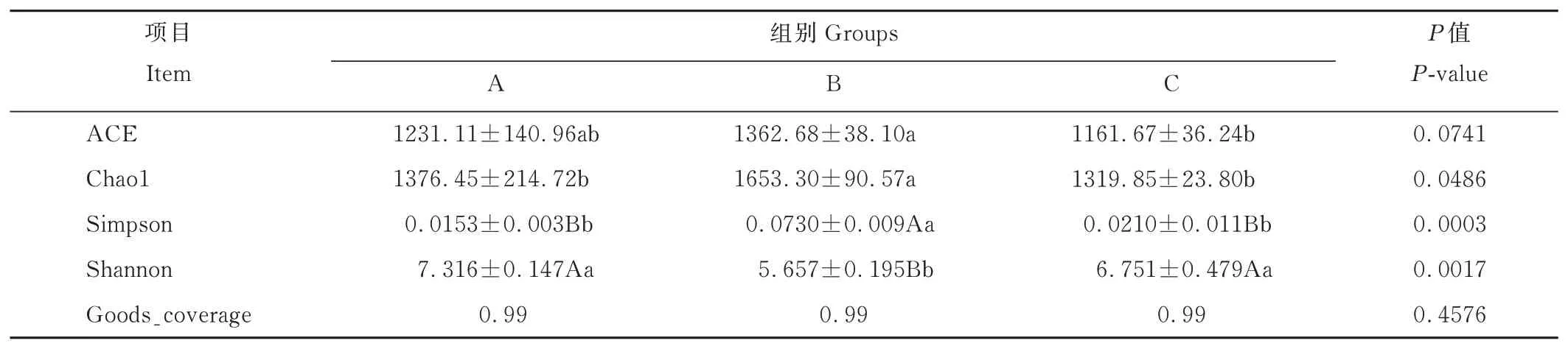
表4 精氨酸水平對離乳期梅花鹿瘤胃微生物α多樣性的影響Table 4 Effects of different arginine levels on rumen microbial alpha diversity of weaning sika deer
Beta多樣性分析是對不同樣本的微生物群落構成進行比較分析,基于Weighted Unifrac距離進行非加權組平均聚類分析(UPGMA)(圖2)和主坐標分析(PCoA)(圖3),并通過Amova分析(表5)檢驗各組間細菌群落結構差異顯著性,結果顯示,B、C組仔鹿瘤胃內細菌群落結構較為相似(P>0.05),而二者與A組群落結構差異顯著(P<0.05)。

表5 Amova組間差異分析Table 5 Analysis by Amova group differences analysis
2.3 精氨酸水平對瘤胃細菌組成的影響
如圖4所示,根據物種注釋結果,本研究選取了各組仔鹿瘤胃液細菌在門(phylum)水平上豐度排名前10的物種,生成物種相對豐度柱形累加圖,以便直觀的了解各組仔鹿瘤胃細菌在門分類水平上相對豐度較高的細菌種類及其比例;由表6可知,3組的優勢菌門均為擬桿菌門(Bacteroidetes)、厚壁菌門(Firmicutes)、變形菌門(Proteobacteria)。其中,B組擬桿菌門(Bacteroidetes)相對豐度極顯著低于A組(P<0.01),且略低于C組(P>0.05);而B組厚壁菌門(Firmicutes)相對豐度顯著高于A組(P<0.05);B組變形菌門(Proteobacteria)相對豐度極顯著高于A組(P<0.01),顯著高于C組(P<0.05);B組廣古菌門(Euryarchaeota)相對豐度極顯著低于A組(P<0.01);B組放線菌門(Actinobacteria)相對豐度極 顯 著 高 于A、C組(P<0.01);A組 螺 旋 菌 門(Spirochaetes)、纖維桿菌門(Fibrobacteres)相對豐度極顯著高于B、C組,其中纖維桿菌門(Fibrobacteres)相對豐度在B、C組之間也有顯著差異(P<0.05);C組軟壁菌門(Tenericutes)相對豐度極顯著高于A組(P<0.01),顯 著 高 于B組(P<0.05);互 養 菌 門(Synergistetes)相對豐度在3組之間無顯著差異(P>0.05)。

表6 各組仔鹿瘤胃液細菌門水平群落組成Table 6 Rumen bacteria composition on phylum level in three groups of weaning sika deer(%)
如圖5所示,根據物種注釋結果,選取了在屬(genus)水平上相對豐度排名前20的物種,生成物種相對豐度柱形累加圖,也能直觀的了解各組仔鹿瘤胃細菌在屬分類水平上相對豐度較高的細菌種類及其比例;由表7可知,未分類普雷沃氏菌科(unidentified_Prevotellaceae)為3組共有的優勢菌,且相對豐度在3組之間無顯著差異(P>0.05)。另外,A組的優勢菌還包括甲烷短桿菌屬(Methanobrevibacter)和未分類疣微菌科(unidentified_Ruminococcaceae),B組的優勢菌屬還包括琥珀酸弧菌屬(Succinivibrio)和戴阿利斯特桿菌屬(Dialister),C組的優勢菌屬還包括琥珀酸弧菌屬(Succinivibrio)和解琥珀酸菌屬(Succiniclasticum)。A組甲烷短桿菌屬(Methanobrevibacter)、奎因氏菌屬(Quinella)、纖維桿菌屬(Fibrobacter)相對豐度極顯著高于B、C組(P<0.01);A組未分類螺旋體科(unidentified_Spirochaetaceae)相對豐度極顯著高于B組(P<0.01),顯著高于C組(P<0.05);A組Agathobacter、互營球菌屬(Syntrophococcus)相對豐度極顯著低于B組(P<0.01),顯著低于C組(P<0.05);B組琥珀酸弧菌屬(Succinivibrio)相對豐度極顯著高于A組(P<0.01),顯著高于C組(P<0.05);B組戴阿利斯特桿菌屬(Dialister)相對豐度顯著高于A組(P<0.05);C組解琥珀酸菌屬(Succiniclasticum)、未分類毛螺 菌 科(unidentified_Lachnospiraceae)、聚乙酸菌屬(Acetitomaculum)相對豐度極顯著高于A、B組(P<0.01);C組未分類理研菌科(unidentified_Rikenellaceae)相對豐度顯著高于A、B組(P<0.05);未分類韋榮氏球菌科(unidentified_Veillonellaceae)、歐陸森氏菌屬(Olsenella)、未分類疣微菌科(unidentified_Ruminococcaceae)、雙歧桿菌屬(Bifidobacterium)、Kandleria、未分類擬桿菌目(unidentified_Bacteroidales)、厭氧弧菌屬(Anaerovibrio)相對豐度在3組之間無顯著差異(P>0.05)。
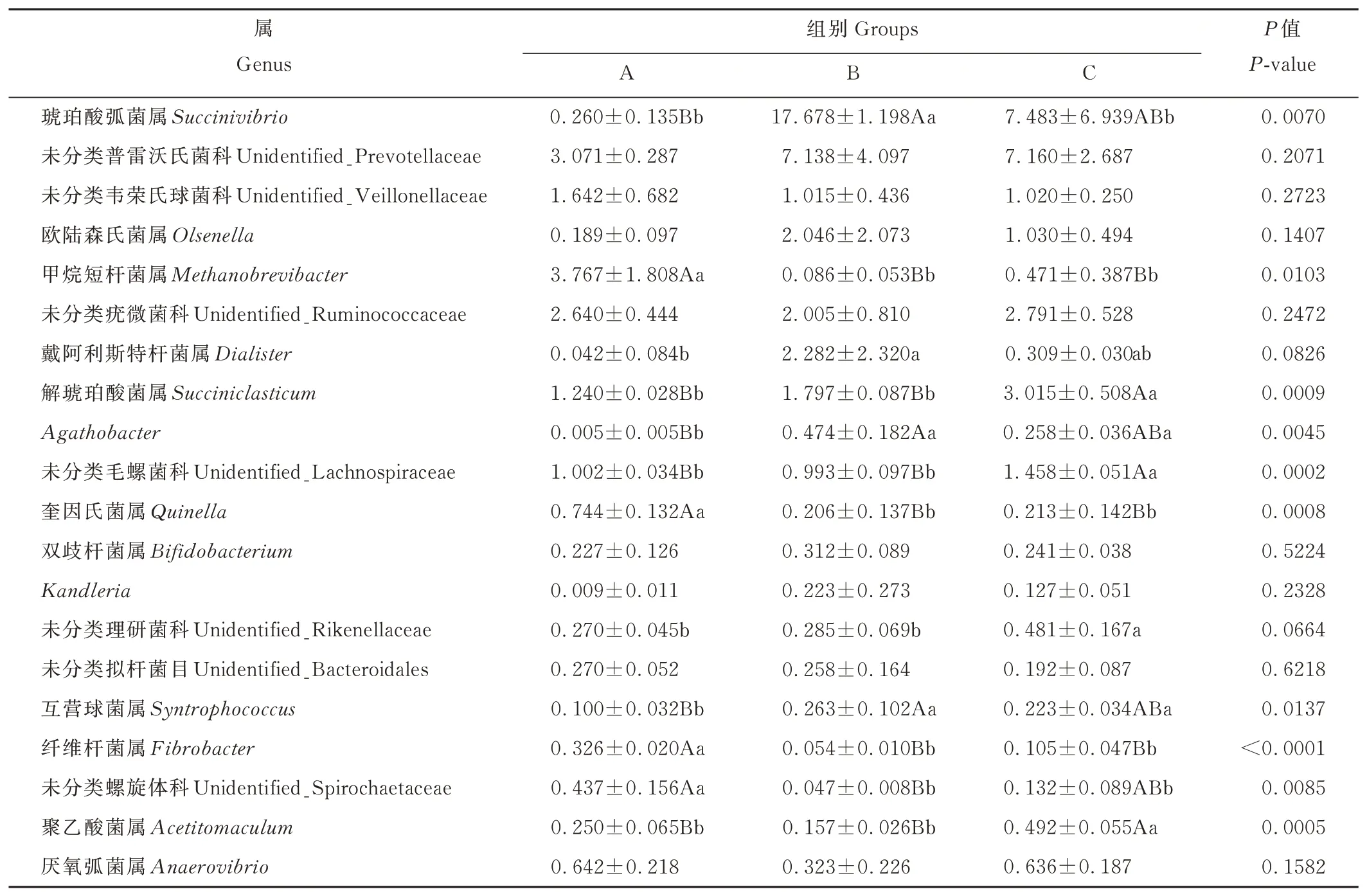
表7 各組仔鹿瘤胃液細菌屬水平群落組成Table 7 Rumen bacteria composition on genus level in three groups of weaning sika deer(%)
3 討論
3.1 精氨酸水平對梅花鹿仔鹿瘤胃發酵參數的影響
瘤胃pH、氨態氮(NH3-N)濃度和揮發性脂肪酸(volatile fatty acid,VFA)比例等參數是研究瘤胃發酵狀況的重要指標,能夠反映飼料在瘤胃內的發酵程度和模式。瘤胃pH會受到飼糧結構、采食速度、唾液分泌量以及瘤胃內VFA含量的影響,VFA含量以及其在瘤胃中的吸收狀況也會因瘤胃pH的波動而受到影響[10-11]。一般情況下,VFA含量增加,瘤胃pH會降低[12-13]。本研究中,隨著精氨酸水平的提高,瘤胃pH、乙酸、丁酸以及TVFA含量的變化趨勢為逐漸降低。3組試驗飼糧結構保持一致,精粗比相同,推測可能與瘤胃微生物種類及數量有關,此外,國內外已有文獻報道[14-16],瘤胃中VFA含量增加會解離出更多的H+,致使pH降低,瘤胃內低pH更有利于VFA在瘤胃上皮被吸收(瘤胃pH在正常范圍時)。因此,瘤胃pH低,可促進VFA的吸收進而導致其在瘤胃中含量降低,本研究結果與上述研究結論一致。由此可間接推測,飼糧精粗比一致的情況下,精氨酸水平為1.08%和1.26%的飼糧增強了仔鹿瘤胃上皮對VFA的吸收能力,進而提高仔鹿對飼料的消化率[17-18]。此外,本研究發現,3組之間丙酸含量無顯著差異,但精氨酸水平為1.08%時仔鹿瘤胃中丙酸含量略高于其他組,而乙丙比極顯著或顯著低于其他組;表明精氨酸水平為1.08%時,仔鹿瘤胃發酵模式以丙酸型發酵為主,精氨酸水平為0.80%組的仔鹿則以乙酸型發酵為主,精氨酸水平為1.26%時介于兩者之間。Ryle等[19]研究表明,瘤胃中發酵產生的乙酸、丙酸和丁酸分別為機體提供的能量為總能量(2805 kJ·mol-1)的62%,109%和78%,因此瘤胃進行丙酸型發酵時能為機體提供更多的能量。Shabat等[20]、李宗軍[21]和董春曉等[22]的研究也證實,當瘤胃發酵模式由乙酸型發酵向產生更多丙酸的模式轉變時,可以有效提高機體能量代謝和飼料轉化效率,促使幼齡反芻動物在早期獲得更大的體重。由此可知,仔鹿飼糧中精氨酸水平為1.08%時,可以改變仔鹿瘤胃發酵模式,提高瘤胃供能效率,促進仔鹿生長發育。瘤胃NH3-N主要由飼糧中的氨基酸、蛋白質和非蛋白氮化合物降解產生,是瘤胃微生物合成菌體蛋白的主要氮源[23-24]。精氨酸是機體合成蛋白質和DNA的重要氮源[25-26]。因此,精氨酸水平可能會影響瘤胃NH3-N濃度,進而影響瘤胃中蛋白降解與菌體蛋白合成間的動態平衡。瘤胃NH3-N濃度正常范圍是5.0~30.0 mg·dL-1[27]。呂小康[28]的研究結果表明,因山羊羔羊的瘤胃NH3-N濃度降低,導致其微生物蛋白的合成受到抑制。本研究中,隨著飼糧精氨酸水平的提高,瘤胃NH3-N濃度呈先上升后下降的趨勢,精氨酸水平為1.26%時仔鹿瘤胃氨態氮濃度顯著或極顯著低于其他兩組,且濃度略低于正常范圍,說明在特定飼糧組成下,精氨酸水平過高,會使仔鹿瘤胃NH3-N濃度降低,從而造成蛋白降解與菌體蛋白合成之間失衡,而合理提高精氨酸水平能夠有效促進瘤胃對飼料的消化吸收,進而提高仔鹿瘤胃微生物活性,促進仔鹿生長。
3.2 精氨酸水平對細菌豐富度和多樣性的影響
新生斷奶仔鹿在特定飼糧結構的影響下,最終構成特定的瘤胃微生物群落,同時造成仔鹿的采食偏好,影響各細菌群落的生長效率及數量[29-31]。通過比較Chao1,ACE指數,可以反映一個群落或生境中物種數目的多少,即群落豐富度;比較Shannon,Simpson指數,可以反映該樣品中的分類總數和每個分類所占的比例,即群落多樣性。群落多樣性越高,物種分布越均勻,Shannon指數值越大,Simpson與之相反。群落覆蓋率(goods_coverage)能夠反映是否取樣充分。本研究中,由表4可知,3組的覆蓋率均為99%,說明本研究取樣充分,能較好地反映出瘤胃液中微生物菌群的多樣性;精氨酸水平為1.08%組的Chao1指數值顯著高于其他組,Simpson指數值極顯著高于其他組,Shannon指數值與其相反,說明精氨酸水平為1.08%時,仔鹿瘤胃細菌較其他組豐富,但各物種分布不均,推測其優勢菌群占總細菌群落比例較高,需要結合瘤胃細菌豐度分析的結果來驗證;另外,本研究通過UPGMA聚類樹和PCoA分析,結合Amova組間差異分析,結果發現,精氨酸水平為1.08%和1.26%時,仔鹿瘤胃細菌具有較相似的群落結構,但與精氨酸水平為0.80%組的仔鹿瘤胃細菌存在明顯差異。這些結果說明,瘤胃液中的細菌群落因精氨酸水平的差異而發生了變化。Dias等[32]、Jami等[33]和Koringa等[34]對犢牛的研究顯示,微生物群落在犢牛生命早期變化很大,隨著固體飼料的攝入,大量微生物逐漸引入并以一種特定的、漸進的順序定殖于瘤胃形成菌群對飼料進行消化利用,隨后逐漸建立穩定的瘤胃微生物區系。早期特定的飼糧結構也會影響定殖的微生物群落豐富度和多樣性。由此可知,本研究飼糧中精氨酸水平能夠影響仔鹿瘤胃細菌的多樣性以及改變細菌群落結構,適當提高精氨酸水平能夠提高細菌豐富度,改變瘤胃中優勢菌群所占的比例。
3.3 精氨酸水平對瘤胃細菌組成的影響
飼料中的碳水化合物被瘤胃內各種細菌發酵,并被轉化成各種揮發性脂肪酸。因此,瘤胃內產生的乙酸、丙酸和丁酸的比例除了受到飼料結構和類型的影響外,還與瘤胃細菌的種類和數量有關[35-36]。本研究對瘤胃菌群門水平下的豐度分析中發現,擬桿菌門、厚壁菌門和變形菌門在所有仔鹿的瘤胃細菌中占主導地位。這與前人對幼齡反芻動物的研究結果一致[37-38],說明仔鹿瘤胃菌群中優勢菌門不會因精氨酸水平的變化而發生改變,與其他幼齡反芻動物一致;但隨著飼糧的變化其相對豐度會發生適應性改變[39]。本研究中,精氨酸水平為1.08%時,與另外兩組相比,仔鹿瘤胃中擬桿菌門豐度有所降低,厚壁菌門豐度則顯著提高;與擬桿菌門主要分解非結構多糖不同,厚壁菌門與結構多糖的降解有關[40]。在飼糧精粗比一致的條件下,結合本研究未發表有關NDF營養消化率數據顯示,精氨酸水平為1.08%時,NDF營養消化率顯著高于其他組,表明適當提高精氨酸水平,可以改變擬桿菌門和厚壁菌門的比例,增加的厚壁菌門促進了飼料中纖維素的消化率;Turnbaugh等[41]指出,厚壁菌門更易攝取飼料中可發酵碳水化合物,從而加快機體對營養物質的吸收利用和能量代謝。本研究還發現,精氨酸水平為0.80%和1.26%時,擬桿菌門與厚壁菌門之和均高于精氨酸水平為1.08%組,造成此結果的原因可能是擬桿菌門與厚壁菌門降解飼糧的速率較慢,需要更多細菌參與。來自擬桿菌門的普雷沃氏菌屬已被證明是反芻動物瘤胃內豐度最高的菌屬[42-43],而本研究中,并未檢測到該菌屬,但檢測到與其相關的未分類普雷沃氏菌科可作為仔鹿的優勢菌屬,在瘤胃內降解飼料中的碳水化合物、蛋白質、氨基酸等營養物質并通過發酵產生乙酸、丙酸等為機體供能[44]。這與Si等[45]的研究結果類似。另外,精氨酸水平為1.08%時,變形菌門豐度顯著高于其他組,該門類下的琥珀酸弧菌屬也是該組仔鹿的絕對優勢菌屬,有研究報道[46-48],琥珀酸弧菌屬通過與產甲烷菌爭奪H作為底物產生琥珀酸進而通過蘋果酸-琥珀酸途徑產生丙酸,從而限制甲烷的生成;精氨酸水平為0.80%組的甲烷短桿菌屬豐度極顯著高于其他兩組,是該組仔鹿的優勢菌屬,該菌屬在其他反芻動物瘤胃微生物的研究中也有報道[49-51]。以上結果可知,適宜水平的精氨酸能夠增加琥珀酸弧菌屬豐度,從而促進瘤胃發酵向丙酸發酵模式轉變,還可以降低甲烷的產生;而精氨酸水平過低,會促使甲烷短桿菌屬豐度增加,導致甲烷排放量增加,從而對環境造成負面影響。值得注意的是,在精氨酸水平為1.08%組的仔鹿瘤胃內檢測到Dialister,為該組菌屬豐度排名第3的菌屬;據報道[52],Dialister是一種專性厭氧的革蘭氏陰性細菌,能夠利用琥珀酸脫羧基生成丙酸;Cui等[53]在牦牛犢瘤胃中也檢測到Dialister且證實該菌種與牦牛犢的生長性能呈正相關。精氨酸水平為1.26%組檢測到的未分類普雷沃氏菌科和琥珀酸弧菌屬的豐度相比于其他兩組并不占優勢,但解琥珀酸菌屬的豐度有所增加,該菌屬主要功能是將琥珀酸轉化為丙酸,為宿主提供能量,但不能發酵飼料中的碳水化合物和氨基酸[54]。因此,精氨酸水平過高可為宿主提供少量丙酸,但并不能促進飼料的消化吸收。
4 結論
在本試驗條件下,飼糧精氨酸水平為1.08%時,能夠顯著提高梅花鹿仔鹿瘤胃菌群豐富度并改變細菌群落結構和優勢菌比例;提高厚壁菌門和變形菌門豐度,降低擬桿菌門豐度;提高未分類普雷沃氏菌科、琥珀酸弧菌屬和Dialister豐度,能夠促使仔鹿的瘤胃發酵方式轉變為丙酸型發酵,增強機體供能。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
學苑創造·A版(2015年11期)2016-01-14 09:03:27
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
山東女子學院學報(2014年6期)2014-03-01 02:24:55
