干旱脅迫對(duì)燕麥幼苗葉片光合特性及活性氧清除系統(tǒng)的影響
2022-10-24 07:42:24吳雨涵劉文輝劉凱強(qiáng)張永超
草業(yè)學(xué)報(bào) 2022年10期
關(guān)鍵詞:影響
吳雨涵,劉文輝,劉凱強(qiáng),張永超
(青海省青藏高原優(yōu)良牧草種質(zhì)資源利用重點(diǎn)實(shí)驗(yàn)室,青海大學(xué)畜牧獸醫(yī)科學(xué)院,青海西寧 810016)
水分是影響作物生長(zhǎng)發(fā)育的主要因素之一,水分虧缺對(duì)植株形態(tài)結(jié)構(gòu)和生理生化會(huì)產(chǎn)生消極影響,嚴(yán)重缺水甚至?xí)鹬仓臧l(fā)生凋亡[1]。葉片作為響應(yīng)水分脅迫最敏感的器官之一,它既是光合作用的主要器官,同時(shí)也是植株內(nèi)部互相連接及植株與外界進(jìn)行物質(zhì)交換和能量傳遞的紐帶[2-3]。研究表明,干旱脅迫導(dǎo)致植株葉片相對(duì)含水量下降,葉綠素分解加快而合成速度受到抑制,從而引起葉片失綠,干物質(zhì)積累遞減[4],進(jìn)而產(chǎn)生大量的活性氧物質(zhì),如單線態(tài)氧(singlet oxygen,1O2)、超氧化物(superoxide,O2·-)、超氧化物陰離子自由基(superoxide anion,和過(guò)氧化氫(hydrogen peroxide,H2O2)等。過(guò)量的氧自由基使細(xì)胞膜遭受氧化毒害,會(huì)造成光合速率降低、葉綠素和類(lèi)胡蘿卜素發(fā)生降解等一系列生理生化現(xiàn)象[5]。研究發(fā)現(xiàn),嚴(yán)重干旱甚至?xí)鹑~片功能器官發(fā)生不可逆的機(jī)械損傷,如葉綠體、類(lèi)囊體、線粒體等的結(jié)構(gòu)皺縮形變、功能衰退,最終導(dǎo)致整個(gè)植株發(fā)生病變。
在植物長(zhǎng)期的進(jìn)化過(guò)程中,形成了一套自我防御策略,通過(guò)體內(nèi)的酶類(lèi)抗氧化劑和非酶類(lèi)抗氧化劑相互協(xié)調(diào),清除過(guò)量的活性氧物質(zhì),前者含有超氧化物歧化酶(superoxide dismutase,SOD)、過(guò)氧化物酶(peroxidase,POD)、過(guò)氧化氫酶(catalase,CAT)、抗壞血酸過(guò)氧化物酶(aseorbate peroxidase,APX)、谷胱甘肽還原酶(glutathione reductase,GR)、谷胱甘肽S-轉(zhuǎn)移酶(glutathione S-transferase,GST)、單脫氫抗壞血酸還原酶(monodehydroascorbate reductase,MDHAR)和脫氫抗壞血酸還原酶(dehydrogenase,DHAR),后者則由谷胱甘肽(glutathione,GSH)、抗壞血酸(ascorbic acid,ASA)和類(lèi)胡蘿卜素(carotenoids,Car)組成[6],保護(hù)細(xì)胞處于一個(gè)良好的生境,維持機(jī)體的正常運(yùn)轉(zhuǎn),保證植物在遭受水分脅迫后仍能繼續(xù)正常生長(zhǎng)發(fā)育。目前關(guān)于干旱脅迫對(duì)燕麥(Avena sativa)幼苗活性氧清除系統(tǒng)的研究多集中在抗氧化酶活性的響應(yīng)方面[7-8],而關(guān)于非酶類(lèi)抗氧化劑對(duì)水分脅迫響應(yīng)及在不同水分脅迫程度下酶類(lèi)抗氧化劑及非酶類(lèi)抗氧化劑在活性氧清除系統(tǒng)中發(fā)揮作用方面的研究較少。探究作物在不同干旱脅迫程度下葉片的光合特性及其防御機(jī)制對(duì)于了解作物生長(zhǎng)發(fā)育及指導(dǎo)農(nóng)業(yè)生產(chǎn)具有重要意義。
燕麥?zhǔn)呛瘫究蒲帑湆伲ˋvena)一年生草本植物,具有喜涼耐寒,對(duì)土壤適應(yīng)性強(qiáng),病蟲(chóng)害少,耐瘠薄、抗雜類(lèi)草性能較強(qiáng)等特點(diǎn)[9],適宜在青藏高原高寒牧區(qū)和半農(nóng)半牧區(qū)種植,燕麥作為高產(chǎn)優(yōu)質(zhì)、糧飼兼用作物[10],對(duì)青海省畜牧業(yè)生產(chǎn)的穩(wěn)定發(fā)展起到了重要作用[11-12]。青海地處青藏高原,年降水量少且分布不均勻,燕麥播種時(shí)期大多集中在4-5月,而降水多集中于6-8月,苗期是燕麥需水的關(guān)鍵期,種子萌發(fā)過(guò)程中內(nèi)藏物質(zhì)已經(jīng)消耗殆盡,此時(shí)燕麥莖稈相對(duì)較弱,有效葉片數(shù)相對(duì)較少,植株存儲(chǔ)的能量和營(yíng)養(yǎng)物質(zhì)較少,光合作用積累的養(yǎng)分十分有限,此時(shí)受到干旱脅迫燕麥光合能力下降,進(jìn)而對(duì)幼苗生長(zhǎng)造成致命影響,導(dǎo)致其后期籽粒產(chǎn)量下降。前人對(duì)燕麥苗期干旱的研究大多集中在植株體內(nèi)糖分積累、內(nèi)源激素轉(zhuǎn)運(yùn)、光合特性變化及外源物質(zhì)對(duì)干旱的緩解作用方面[13-15],而關(guān)于燕麥苗期遭受干旱脅迫后葉片光合特性及其抗氧化防御調(diào)控策略尚不清楚。因此,本研究擬以青海本地推廣品種‘青燕1號(hào)’為研究對(duì)象,采用不同的水分脅迫梯度,比較分析干旱脅迫對(duì)燕麥苗期葉片的影響,以期為青藏高原地區(qū)抗旱燕麥品種評(píng)價(jià)提供理論基礎(chǔ)和科學(xué)依據(jù)。
1 材料與方法
1.1 供試材料
供試品種為青海本地推廣品種:青燕1號(hào)(Avena sativa‘Qingyan No.1’),供試燕麥種子由青海省畜牧獸醫(yī)科學(xué)院提供。
1.2 試驗(yàn)設(shè)計(jì)
試驗(yàn)于2019年12月于青海省畜牧獸醫(yī)科學(xué)院省級(jí)重點(diǎn)實(shí)驗(yàn)室進(jìn)行。挑選顆粒飽滿、大小一致的燕麥種子,用2% NaClO滅菌15 min,去離子水洗滌3~5次晾干備用。采用沙培法將消毒后的種子點(diǎn)播于培養(yǎng)穴中,放置于晝夜溫度為20℃/15℃的人工氣候培養(yǎng)箱中,光周期為16 h/8 h,相對(duì)濕度為65%。每周使用1/2 Hoagland營(yíng)養(yǎng)液澆灌植物兩次。
待幼苗兩葉期時(shí),采用干旱脅迫處理燕麥幼苗:1)CK:澆灌1/2 Hoagland營(yíng)養(yǎng)液;2)P10:10%的PEG(polyethylene glycol,-0.53 MPa)干旱脅迫處理;3)P20:20%的PEG(-1.10 MPa)干旱脅迫處理。每個(gè)處理3次重復(fù),分別在脅迫4、7和10 d時(shí)采集葉片樣品,液氮冷凍后保存在-80℃超低溫冰箱中,用于后期生理生化指標(biāo)分析。
1.3 測(cè)定指標(biāo)與方法
1.3.1氣體交換參數(shù)的測(cè)定 每個(gè)處理選取3株長(zhǎng)勢(shì)均勻的幼苗,用便攜式光合儀(Li-6800,LI-COR,USA)于9:00-12:00測(cè)定每株幼苗旗葉的光合參數(shù),每個(gè)葉片重復(fù)3次。葉室各參數(shù)指標(biāo)如下:光照強(qiáng)度1200 μmol·m-2·s-1、CO2濃度(400±5.0)μmol·mol-1(CO2小鋼瓶提供)、相對(duì)濕度(60%±1.0%)、溫度25℃。待葉室穩(wěn)定讀取凈光合速率(net photosynthetic rate,Pn)、蒸騰速率(transpiration rate,Tr)、胞間CO2濃度(intercellular CO2concentration,Ci)和氣孔導(dǎo)度(stomatal conductance,Gs)。
1.3.2葉綠素?zé)晒鈪?shù)的測(cè)定 測(cè)定時(shí)間與氣體交換參數(shù)同步,幼苗在黑暗條件下適應(yīng)1 h后,每個(gè)處理選取3株長(zhǎng)勢(shì)均勻的幼苗,測(cè)定每株幼苗的旗葉,每個(gè)葉片重復(fù)3次。飽和脈沖設(shè)為8000 μmol·m-2·s-1,記錄初始熒光產(chǎn)量(minimal fluorescence,F(xiàn)o)、最大暗適應(yīng)態(tài)熒光產(chǎn)量(maximal fluorescence yield of the dark-adapted state,F(xiàn)m)、最大光適應(yīng)態(tài)熒光產(chǎn)量(maximal fluorescence yield of the light-adapted state,F(xiàn)m')、可變熒光(variable fluorescence,F(xiàn)v)和穩(wěn)態(tài)熒光產(chǎn)量(steady state fluorescence yield,F(xiàn)s)等參數(shù)。參考王迪[16]的計(jì)算方法,計(jì)算光系統(tǒng)Ⅱ(PSII)的最大光化學(xué)效率(maximal photochemistry efficiency,F(xiàn)v/Fm):Fv/Fm=(Fm-Fo)/Fm;PSII實(shí)際光化學(xué)效率(practical efficiency,ΦPSⅡ):ΦPSⅡ=(Fm'-Fs)/Fm';非光化學(xué)猝滅(non-photochemical quenching,NPQ):NPQ=(Fm-Fm')/Fm'。
1.3.3測(cè)定葉片生理指標(biāo) 葉綠素(chlorophyll,Chl)和類(lèi)胡蘿卜素(carotenoids)含量采用分光光度計(jì)法進(jìn)行測(cè)定;超氧陰離子(O2-)含量參考Schneider等[17]的方法測(cè)定;過(guò)氧化氫(H2O2)含量參考Liu等[18]的方法進(jìn)行測(cè)定。
1.3.4測(cè)定酶類(lèi)和非酶類(lèi)抗氧化劑的含量 超氧化物歧化酶活性(SOD)參考Asish等[19]的方法,過(guò)氧化氫酶(CAT)測(cè)定參考Zhang等[20]的方法,抗壞血酸過(guò)氧化物酶活性(APX)及脫氫抗壞血酸還原酶活性(DHAR)采用Yoshiyuki等[21]的方法進(jìn)行測(cè)定,谷胱甘肽過(guò)氧化物酶(GPX)采用Elia等[22]的方法進(jìn)行測(cè)定,谷胱甘肽還原酶(GR)活性采用Foyer等[23]的方法進(jìn)行測(cè)定,抗壞血酸(ASA)、谷胱甘肽(GSH)、脫氫抗壞血酸(dehydroascorbic acid,DHA)及氧化型谷胱甘肽(oxidative glutathione,GSSG)含量的測(cè)定參考牛奎舉[24]的方法。
1.4 數(shù)據(jù)分析
采用Microsoft Excel 2016軟件進(jìn)行數(shù)據(jù)處理與作圖,采用SPSS 16.0軟件進(jìn)行方差分析、Duncan多重比較、多元回歸分析及通徑分析,采用Origin 2018作圖。
2 結(jié)果與分析
2.1 不同干旱脅迫對(duì)燕麥葉片生理指標(biāo)的影響
從表1可以看出,干旱脅迫程度和干旱脅迫時(shí)間對(duì)Chl、Car、H2O2、O2-含量的影響均達(dá)到極顯著水平(P<0.01),干旱時(shí)間和干旱程度的互作效應(yīng)對(duì)Chl、Car、H2O2、O2-含量的影響達(dá)到極顯著水平(P<0.01)。其中,各因素對(duì)Chl、Car、H2O2含量影響大小表現(xiàn)為:干旱程度>干旱時(shí)間>干旱時(shí)間和程度互作,O2-含量表現(xiàn)為干旱時(shí)間>干旱時(shí)間與程度互作>干旱程度。
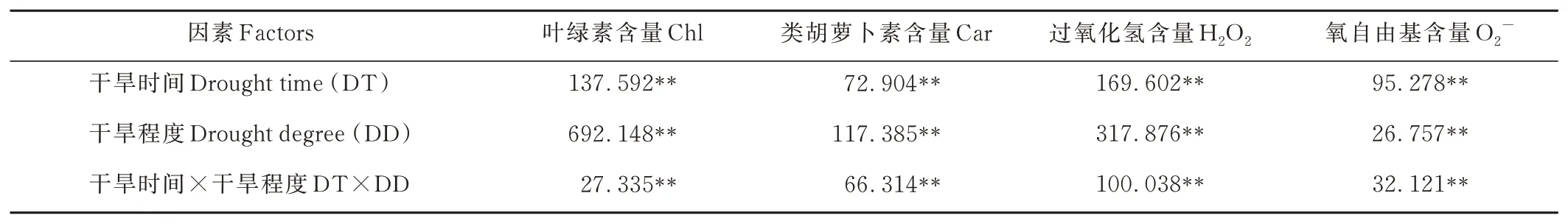
表1 干旱脅迫對(duì)燕麥葉片生理指標(biāo)影響的方差分析Table 1 The variance analysis of effects on oat leaf physiological indicators under drought stress
不同干旱時(shí)間處理對(duì)燕麥葉片Chl含量影響顯著(P<0.05)(圖1a),其中Chl含量在P10和P20處理下顯著低于CK(P<0.05)。隨著干旱時(shí)間和干旱程度的加劇,Chl含量在P20處理第10天時(shí)達(dá)到最低,相比CK降低46.9%。P10和P20處理下Car含量在第7和10天顯著增加(P<0.05),而在第4天沒(méi)有顯著變化(P>0.05)(圖1b)。
P10和P20處理下O2-和H2O2的生成速率在4 d內(nèi)變化不大(圖1c,d),但在7和10 d內(nèi)顯著增加(P<0.05)。與CK相 比,P10和P20在7~10 d O2-、H2O2含量增加幅度分別為3.18~16.51 mmol·min-1·g-1和55.20~158.30 mmol·min-1·g-1。
2.2 不同干旱脅迫對(duì)燕麥氣體交換參數(shù)的影響
從表2可以看出,干旱程度和干旱時(shí)間對(duì)Pn、Tr、Gs、Ci的影響達(dá)到極顯著水平(P<0.01),干旱時(shí)間和干旱程度的互作效應(yīng)對(duì)Pn、Tr、Gs、Ci達(dá)到極顯著水平(P<0.01)。其中,各因素對(duì)Pn、Tr、Gs、Ci的影響大小表現(xiàn)為:干旱程度>干旱時(shí)間>干旱程度與干旱時(shí)間互作。

表2 干旱脅迫對(duì)燕麥氣體交換參數(shù)影響的方差分析Table 2 The variance analysis of effects on oat gas exchange parameters under drought stress
從不同干旱時(shí)間及干旱程度對(duì)‘青燕1號(hào)’燕麥幼苗光合參數(shù)的影響來(lái)看(表3),P10和P20的Pn從第4天均開(kāi)始下降。干旱脅迫對(duì)Pn有抑制作用,4、7和10 d干旱脅迫對(duì)Pn的抑制作用明顯高于CK。
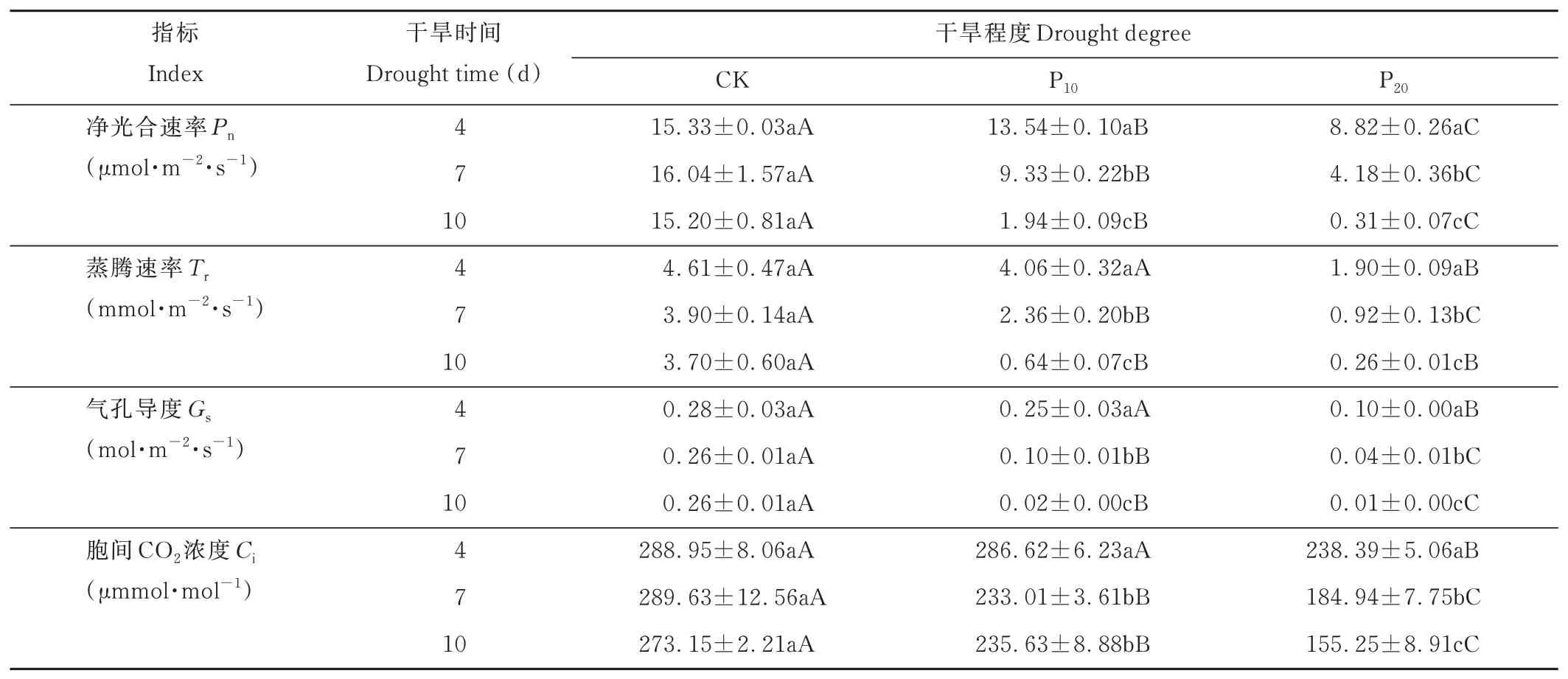
表3 不同干旱脅迫下燕麥氣體交換參數(shù)的變化Table 3 Changes of gas exchange parameters of oat under different water stress
在第4天,CK和P10處理之間的Tr、Gs和Ci均無(wú)顯著差異(P<0.05),而P20處理下的Tr和Gs在第4天開(kāi)始顯著增加(P<0.05)。干旱誘導(dǎo)Tr、Gs和Ci在4~10 d內(nèi)持續(xù)下降,在P20處理10 d后的下降幅度分別比CK下降93.0%、96.1%和43.2%。
2.3 不同干旱脅迫對(duì)燕麥幼苗熒光特性的影響
從表4可以看出,干旱時(shí)間對(duì)Fo、Fv/Fm、ΦPSⅡ、NPQ的影響達(dá)到極顯著差異水平(P<0.01),干旱程度對(duì)Fv/Fm、ΦPSⅡ、NPQ的影響達(dá)到極顯著差異水平(P<0.01),而干旱程度對(duì)Fo無(wú)顯著影響(P>0.05),干旱時(shí)間與干旱程度的互作效應(yīng)對(duì)Fo、Fv/Fm、ΦPSⅡ、NPQ的影響達(dá)到極顯著差異(P<0.01)。

表4 不同干旱脅迫對(duì)燕麥幼苗熒光特性影響的方差分析Table 4 The variance analysis of effects on oat chlorophyll fluorescence characteristics under drought stress
由表5可知,在P10干旱條件下,F(xiàn)o隨干旱時(shí)間的延長(zhǎng)先增加后降低。處理7 d后,P10和P20顯著增加(P<0.05),分別比CK增加9.7%和11.1%。在干旱脅迫10 d時(shí)P10和P20處理下的Fo開(kāi)始明顯下降,其中P20下降幅度最大,為30.0%。
4 d時(shí)所有處理對(duì)Fv/Fm無(wú)明顯影響(表5)。干旱脅迫7~10 d時(shí),與CK相比,P20處理下的Fv/Fm下降高于P10處理。

表5 不同水分脅迫下燕麥熒光參數(shù)的變化Table 5 Changes of fluorescence parameters of oat under different water stress
在P20處理下,ΦPSⅡ第4天開(kāi)始下降,在第10天達(dá)到最低,在第4和10天分別比CK下降了28.3%和68.2%。P10和P20的NPQ隨著干旱時(shí)間和干旱程度的增加而顯著增加(P<0.05),其中第10天NPQ增加最多,分別比CK增加147.2%和134.7%。
2.4 不同干旱脅迫對(duì)燕麥活性氧清除系統(tǒng)的影響
植物的多種代謝過(guò)程均可產(chǎn)生活性氧自由基,體內(nèi)活性氧成分在正常水平下處于一個(gè)動(dòng)態(tài)平衡,適量的活性氧還可作為信號(hào)分子,調(diào)控細(xì)胞基因轉(zhuǎn)錄[25-26]。當(dāng)植物遭受水分脅迫后會(huì)打破這個(gè)動(dòng)態(tài),活性氧的產(chǎn)生大于其清除速率,造成植物組織遭受氧化脅迫,破壞細(xì)胞膜脂;而植物在應(yīng)對(duì)活性氧脅迫的過(guò)程中,進(jìn)化形成了一套由酶類(lèi)和非酶類(lèi)抗氧化劑組成的防御系統(tǒng),清除活性氧帶來(lái)的負(fù)面效應(yīng),來(lái)幫助植物在遭受環(huán)境變化時(shí)得以生存。
2.4.1不同干旱脅迫對(duì)燕麥抗氧化酶活性的影響 從表6可以看出,干旱程度對(duì)SOD、CAT、APX、GPX、DHAR活性的影響均達(dá)到極顯著水平(P<0.01),而對(duì)GR無(wú)影響(P>0.05),干旱時(shí)間對(duì)SOD、GPX、DHAR活性的影響均達(dá)到極顯著水平(P<0.01),而對(duì)CAT、APX、GR無(wú)影響(P>0.05),干旱時(shí)間與干旱程度的互作效應(yīng)對(duì)SOD、CAT、APX、GPX、DHAR、GR的影響均達(dá)到極顯著(P<0.01)差異水平。
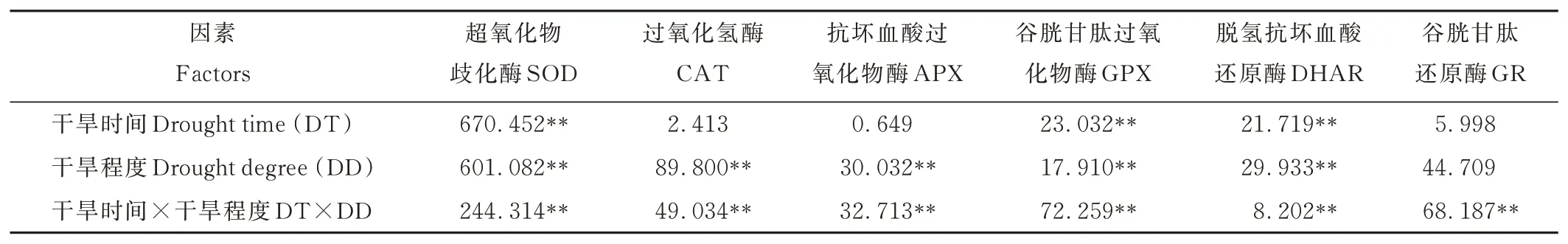
表6 不同干旱脅迫對(duì)燕麥抗氧化酶活性的方差分析Table 6 The variance analysis of effects on activity of enzymatic antioxidants under different drought stress
不同干旱脅迫對(duì)燕麥葉片抗氧化酶活性的影響結(jié)果表明(圖2),隨著干旱脅迫程度及持續(xù)時(shí)間的增加,SOD活性持續(xù)升高。其中,P20的SOD活性升高幅度最大,P10次之。說(shuō)明隨著水分脅迫加劇其清除活性氧能力逐漸下降(圖2a)。
干旱脅迫4 d的CAT活性相比CK顯著增高(P<0.05),P10和P20處理分別增加7.9%和17.0%。隨著干旱持續(xù)時(shí)間的增加,P10處理在7~10 d比CK顯著增加(P<0.05),但P20并未出現(xiàn)此規(guī)律(圖2b)。
在干旱脅迫 下,APX活 性 在P10和P20處理下4和7 d較 高(圖2c),P10處理在4和7 d分別達(dá)到5.02和4.70 μmol·min-1·g-1,P20處理在4和7 d分別達(dá)到4.91和5.12 μmol·min-1·g-1。第10天時(shí),P10處理下APX活性仍然高于CK,而P20處理下APX活性開(kāi)始急劇下降。
在干旱脅迫處理期間,GPX在第4和7天的變化與APX相似(圖2d)。GPX活性在P10和P20處理下4和7 d與CK相比均明顯增加(P<0.05)。10 d后,P10處理下GPX活性然高于CK,P20處理下則低于CK。
干旱脅迫4~7 d時(shí),DHAR活性在P10處理下高于CK,而P20始終保持下降趨勢(shì),且在干旱持續(xù)時(shí)間內(nèi)顯著低于CK(P<0.05)(圖2e)。
GR活性在P10和CK處理下在第4和7天無(wú)明顯變化(圖2f),隨著干旱持續(xù)時(shí)間的增加,P10處理下GR活性在第10天逐漸增加。P20處理7 d時(shí)GR活性最高,在第10天時(shí)急劇下降。
2.4.2不同干旱脅迫對(duì)燕麥非酶類(lèi)抗氧化劑的影響 從表7可以看出,干旱時(shí)間和脅迫程度對(duì)ASA、GSH、DHA、GSSG、ASA/DHA、GSH/GSSG的影響均達(dá)到顯著水平(P<0.05)或極顯著水平(P<0.01)。干旱時(shí)間和干旱程度互作效應(yīng)對(duì)ASA、GSH、GSSG、ASA/DHA的影響達(dá)到極顯著(P<0.01)水平,而對(duì)DHA、GSH/GSSG的影響無(wú)顯著差異(P>0.05)。
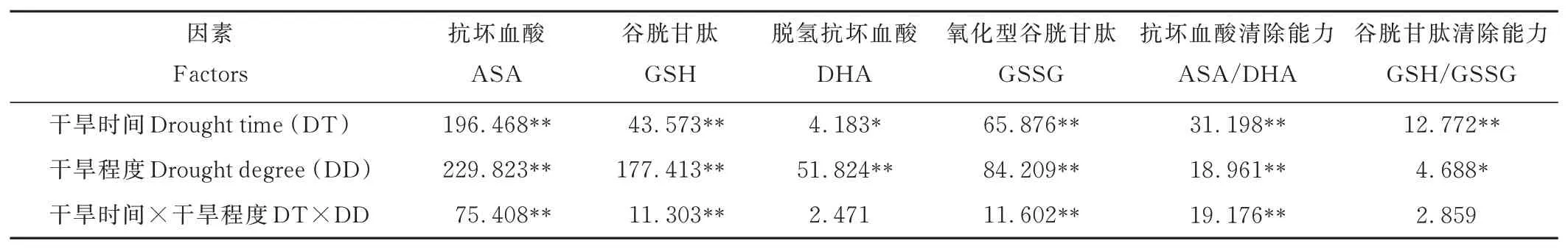
表7 不同干旱脅迫對(duì)燕麥非酶抗氧化劑影響的方差分析Table 7 The variance analysis of effects on activity of non-enzymatic antioxidants under different drought stress
由圖3a,b可知,干旱脅迫處理后ASA和GSH活性顯著增加(P<0.05)。P20處理第10天ASA含量處于最高水平,相比CK增加了589.6%,而GSH含量在P10和P20處理4~7 d均保持較高水平。
氧化型產(chǎn)物DHA和GSSG含量對(duì)干旱脅迫的響應(yīng)相似(圖3c,d)。在4~10 d,P10處理下的DHA含量比CK分別增加了66.7%、160.0%和146.8%,P20的DHA含量也分別增加了107.9%、168.1%和100.3%。在干旱4~10 d,P10處 理 下 的GSSG含 量 比CK分 別 增 加 了12.3%、52.8%和69.5%,P20的GSSG含 量 也 分 別 增 加 了17.5%、93.7%和51.4%。
GSH/GSSG和ASA/DHA大小可以反映非酶類(lèi)抗氧化劑清除活性氧的能力(圖3e,f),在所有處理4~7 d時(shí),ASA/DHA未發(fā)生顯著變化(P>0.05),在干旱處理10 d時(shí)顯著增加(P<0.05),P10和P20處理下ASA/DHA分別增加了8.0%和244.4%。干旱導(dǎo)致GSH/GSSG在第4天明顯增加,而隨著持續(xù)時(shí)間的增加,與CK相比,P10與P20處理分別在第10和7天開(kāi)始呈遞減趨勢(shì),說(shuō)明隨著干旱脅迫增加,其清除活性氧能力在逐漸降低。
3 討論
3.1 不同干旱脅迫對(duì)葉片光合特性的影響
葉綠素是葉片光合同化的主要色素,在酸、堿、氧化脅迫下極易發(fā)生分解[27],水分脅迫下葉綠素含量的變化可以在一定程度上反映植物的生產(chǎn)性能和抵抗逆境脅迫的能力[28],類(lèi)胡蘿卜素為天然水溶性自由基清除劑,在植物吸收光能、保護(hù)葉綠素、猝滅活性氧方面具有重要作用[29]。在本研究中,干旱脅迫下的葉綠素含量隨時(shí)間的增加而不斷下降,而類(lèi)胡蘿卜素含量則隨時(shí)間的延長(zhǎng)而增加。此外,隨著脅迫梯度的增加,O2-和H2O2含量顯著高于對(duì)照,葉綠素與O2-和H2O2呈負(fù)相關(guān),與類(lèi)胡蘿卜素則呈顯著正相關(guān)(圖1)。這表明活性氧(reactive oxygen species,ROS)的增加可能加劇了燕麥的氧化損傷程度,抑制了葉綠素含量,而類(lèi)胡蘿卜素的增加可以緩解氧化損傷。
3.2 干旱脅迫對(duì)葉片生理特性的影響
影響植物光合速率的因素主要有氣孔限制和非氣孔限制,前者多為可逆修復(fù),后者一般為不可逆損傷。本試驗(yàn)得出,不同水分脅迫下,Pn、Tr、Ci和Gs下降明顯,而氣孔限制值顯著提高,其中Gs變化最為敏感,根據(jù)氣孔限制學(xué)說(shuō)可知,導(dǎo)致Pn下降的主要原因是氣孔關(guān)閉,葉片的氣孔導(dǎo)度減小,引起胞間CO2濃度下降,最終導(dǎo)致凈光合速率下降;不同水分脅迫下,凈光合速率下降幅度大于蒸騰速率,說(shuō)明氣孔限制對(duì)蒸騰速率的影響大于凈光合速率,本研究結(jié)果與此一致。水分利用效率表示作物消耗單位質(zhì)量的水所生成的干物質(zhì)量,高水分利用效率往往是作物抵御水分脅迫的重要策略,主要原因是氣孔限制對(duì)蒸騰速率的影響大于凈光合速率,本試驗(yàn)得出同樣結(jié)論,隨著水分脅迫梯度增加,蒸騰速率的下降幅度大于凈光合速率,水分利用效率呈遞增趨勢(shì)。
光合電子傳遞產(chǎn)生活性氧,活性氧的過(guò)量積累對(duì)光合系統(tǒng)造成損害,葉綠素?zé)晒鈩?dòng)力學(xué)可以從能量代謝及轉(zhuǎn)化的角度快速、靈敏、無(wú)損傷的探測(cè)逆境對(duì)植物光合作用的影響,是植物遭受水分脅迫危害的一種理想監(jiān)測(cè)手段[30-32]。Fo代表初始熒光,是光系統(tǒng)Ⅱ(PSⅡ)反應(yīng)中心完全開(kāi)放時(shí)的熒光產(chǎn)量,逆境時(shí)Fo增加被認(rèn)為是誘導(dǎo)光系統(tǒng)Ⅱ活性中心失活的結(jié)果,其變化與葉綠素濃度有關(guān)。本試驗(yàn)得出,在短時(shí)間、輕度脅迫時(shí)Fo顯著上升,但隨著脅迫時(shí)間及程度增加,葉綠素降解速度加快,F(xiàn)o的含量呈下降趨勢(shì)。Fv/Fm和ΦPSⅡ反應(yīng)PSⅡ反應(yīng)中心原初光能轉(zhuǎn)化效率和實(shí)際光能轉(zhuǎn)化效率,當(dāng)植物遭受脅迫時(shí),PSⅡ活性中心降低,光能轉(zhuǎn)化效率下降,過(guò)剩光能對(duì)光合機(jī)構(gòu)產(chǎn)生光抑制效應(yīng),因此,F(xiàn)v/Fm和ΦPSⅡ減小常常被認(rèn)為是植物發(fā)生光抑制和光合電子傳遞速率減弱的表現(xiàn)[33-34]。本試驗(yàn)顯示,隨著脅迫濃度增加,F(xiàn)v/Fm和ΦPSⅡ明顯降低,說(shuō)明缺水造成了燕麥PSⅡ活性中心的損傷,導(dǎo)致活性氧增加造成光系統(tǒng)中心對(duì)光吸收和利用效率降低。此外,與對(duì)照相比,NPQ含量顯著升高,說(shuō)明過(guò)剩光能以熱消耗的形式散失,避免了過(guò)剩光能抑制光合機(jī)構(gòu)活性。
3.3 干旱脅迫對(duì)葉片抗氧化防御應(yīng)對(duì)策略的影響
抗氧化防御系統(tǒng)在植物緩解逆境脅迫方面具有重要作用,在植物的正常代謝過(guò)程中葉綠體、線粒體等在進(jìn)行電子傳遞的過(guò)程均可產(chǎn)生活性氧[35],少量的活性氧對(duì)植株沒(méi)有毒害作用,但活性氧大量積累會(huì)對(duì)細(xì)胞產(chǎn)生氧化脅迫[36],植物在進(jìn)化過(guò)程中建立了酶類(lèi)抗氧化和非酶類(lèi)抗氧化劑防御體系,二者協(xié)同作用能迅速清除體內(nèi)過(guò)量的活性氧物質(zhì),保持機(jī)體處于良性循環(huán)狀態(tài)。
酶類(lèi)抗氧化劑被譽(yù)為最有效、最直接的活性氧清除系統(tǒng)[33],包括SOD、CAT、APX、GPX、DHAR和GR等。SOD是清除活性氧的第一道防御,它能催化O2-歧化生成H2O2,而產(chǎn)生的H2O2可被CAT、APX、GPX清除。其中CAT能直接清除H2O2生成H2O,而APX、GPX必須要有ASA和GSH的參與,二者催化H2O2氧化ASA和GSH生成氧化型物質(zhì)DHA和GSSG[37]。
從不同干旱脅迫對(duì)燕麥葉片抗氧化酶活性的影響結(jié)果來(lái)看(圖2),隨著干旱脅迫程度及持續(xù)時(shí)間的增加,SOD活性持續(xù)升高。其中,P20的SOD活性升高幅度最大,P10次之。說(shuō)明隨著水分脅迫加劇其清除活性氧能力逐漸下降。本研究中,以P20處理下SOD和CAT活性最高,隨著脅迫濃度增加酶活性顯著下降;而APX和GPX活性隨著脅迫濃度增加呈上升趨勢(shì),其中P20處理下,APX活性逐漸下降,而GPX則未發(fā)生變化,說(shuō)明輕度脅迫主要以SOD和CAT清除活性氧為主,而重度脅迫下以APX和GPX依賴ASA、GSH清除占主導(dǎo)地位。DHAR和GR能將DHA和GSSG還原為ASA和GSH,源源不斷地為APX和GPX提供底物,因此,APX、GPX、DHAR和GR的完美協(xié)同是機(jī)體高效清除活性氧的重要信號(hào)。本研究中,DHAR和GR受水分脅迫影響變化明顯,在重度脅迫下DHAR和GR活性顯著低于對(duì)照。
類(lèi)胡蘿卜素、ASA和GSH是非酶類(lèi)抗氧化劑的重要組成部分,類(lèi)胡蘿卜素能減輕活性氧對(duì)細(xì)胞遺傳物質(zhì)和細(xì)胞膜的損傷[38],而ASA和GSH是ASA-GSH循環(huán)的重要參與者。本研究中,水分脅迫下類(lèi)胡蘿卜素、ASA、GSH含量顯著增加,這與董守坤等[39]的研究結(jié)果一致,水分脅迫促使非酶類(lèi)抗氧化劑增加,解除或減輕燕麥遭受的氧化脅迫。APX、GPX、DHAR和GR四種酶對(duì)ASA-GSH循環(huán)起重要調(diào)控作用,APX、GPX將ASA、GSH氧化后,DHAR和GR將氧化型產(chǎn)物DHA和GSSG又還原成ASA和GSH,形成“氧化-還原-氧化”的循環(huán),因此,有研究也常將ASA/DHA和GSH/GSSG的大小作為非酶類(lèi)抗氧化劑清除能力的體現(xiàn)。
本研究中,ASA、GSH、GSSG和DHA均隨著水分脅迫程度加深有不同程度的增加,ASA/DHA和GSH/GSSG隨水分脅迫增加或持續(xù)下降或先升高后下降,說(shuō)明雖然水分脅迫引起了ASA、GSH等抗氧化劑的增加,但其氧化型物質(zhì)同樣也在增加,而DHAR和GR的還原力是有限的,活性氧物質(zhì)積累最終導(dǎo)致燕麥清除速度小于其生成速度,其清除活性氧能力在逐漸降低,最終抗氧化系統(tǒng)開(kāi)始衰變。
4 結(jié)論
1)干旱脅迫能引起植株短時(shí)間內(nèi)水分利用效率增加,但氣孔關(guān)閉以及光反應(yīng)中心遭到破壞是植株光合性能減弱的主要原因。
2)在輕度脅迫下,‘青燕1號(hào)’燕麥主要通過(guò)酶類(lèi)抗氧化劑清除活性氧毒害物質(zhì),而在重度脅迫下以非酶類(lèi)抗氧化劑清除系統(tǒng)為主。
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(xiāo)(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級(jí)版)(2011年3期)2011-01-01 00:00:00
