雞IGF1R基因遺傳變異與其體尺和屠宰性狀的相關性
2022-10-21 11:13:16楊禮楊蛟陳磊杜勇張勝國牟慧蓉冉隆權張依浴
畜牧獸醫科學 2022年7期
關鍵詞:分析
楊禮,楊蛟,陳磊,杜勇,張勝國,牟慧蓉,冉隆權,張依浴
(1.貴州省沿河縣農業農村局,貴州 銅仁 565300;2.貴州省銅仁市農業農村局,貴州 銅仁 554300;3.貴州大學,貴州 貴陽 550000)
0 引言
世界各地的許多雞品種不僅需要通過選擇培育去提高它的生長特性,而且要提高屠宰和肉質屬性。傳統的育種方法得到了各種分子遺傳學工具的支持,以提高生產性狀的遺傳增益率,加速育種目標的實現。在標記輔助選擇中,使用對這些特性具有顯著和積極影響的適當分子標記可以改善胴體和肉質性狀。
胰島素樣生長因子1受體(Insulin-like growth factor receptor 1,IGF1R)是一種酪氨酸激酶受體,由異四聚體跨膜糖蛋白的兩個細胞外α亞基(135 kDa)和2個跨膜β亞基(90 kDa)組成[1-2]。IGF1R屬于細胞調節劑家族,由3種激素組成,包括胰島素樣生長因子1(Insulin-like growth factor 1,IGF1),IGF2和胰島素(Insulin),以及另一種稱為胰島素樣生長因子2受體的受體(IGF2R)和6個胰島素樣受體生長因子結合蛋白(Six insulin-like growth factor binding proteins,IGFBP1-6)[3-4]。這些蛋白質充當內分泌、自分泌和旁分泌刺激物,在胚胎發生、細胞生長和分化、有絲分裂、存活和代謝調節中起重要作用[5]。當IGF1,IGF2或胰島素中的任何一種與受體結合,就會發生IGF1R信號,而這種信號可介導并促進肌肉肥大的途徑,抑制有利于肌肉萎縮的生物途徑,進而對動物的生長發育起調控作用。由于具有上述功能,IGF1R基因已被認為多種動物生長和胴體性狀的功能候選基因[6]。
在雞體內,IGF1R基因定位于10號染色體上,含有21個外顯子和20個內含子,跨域基因組140 315個bp(NCBI RefSeq:NC_006097.5,區域范圍為17,124,349-17,264,663)編碼一個900個氨基酸多肽(NCBI RefSeq:NM_205032.1)。目前有關IGF1R基因對雞的體尺性狀和大多數屠宰性狀的影響的相關研究較為匱乏,特別是以貴州沿河鐵叫雞為研究動物的研究信息則不存在。本研究目的是進行IGF1R基因的多態性調查,并且評估其多態性及其基因型對雞體尺性狀和屠宰性狀的影響,以期為雞的科學選育培育提供參考。
1 材料與方法
1.1 試驗動物
研究在貴州省沿河縣鐵叫雞育種場選取同批出殼、健康無病和同一飼養環境下的20周齡雞100只,每只雞翅靜脈采取血液2 mL用于總DNA的提取。
1.2 體重與體尺測定
在雞禁食12 h后稱量其活重,按照NY/T823—2004規定測定體尺[7]。
1.3 屠宰性能測定
在測定完體尺指標后將雞頸靜脈放血致死,利用濕拔毛法,根據NY/T823—2004規定測定沿河鐵叫雞的屠宰性狀。
1.4 引物的設計和基因組DNA提取
按照大連寶生物血液基因組DNA提取試劑盒(TaKaRa,Dalian,China)說明書提取基因組DNA。根據NCBI GeneBank已公布雞IGF1R基因(NCBI RefSeq:NC_006097.5)序列。Primer 3(美國Applied Biosystem公司)設計多態引物[8],由上海生工生物工程股份有限公司合成,見表1。

表1 試驗所用引物
1.5 DNA池PCR擴增及測序分型
PCR反應體系30.0 mL:15.0 mL PCR MasterMix,正、反向引物各2.0 mL,DNA模板1.0 mL,12.0 mL ddH2O。按預變性、退火、延伸、終延伸程序進行PCR擴增,產物經檢測后送至生工生物工程(上海)股份有限公司測序。結果經比對分析,篩選出單核苷酸多態性(SNP)位點。通過PCR擴增獲取產物繼續送至生工生物工程(上海)股份有限公司測序,進行基因分型。
1.6 統計分析
在線軟件SHEsis(http://analysis.bio-x.cn/)構建單倍型和雙倍型;PopGene 軟件(ver.1.32)計算等位基因頻率和等位基因型;PowerMarker 軟件(ver.3.25)對多態信息含量(PIC)進行分析;采用PASW Statistics 18.0軟件對屠宰性能和體尺測定數據進行有效性檢測,剔除不符合正態分布個體。一般線性模型(GLM)分析屠宰性能和體尺與IGF1R基因的關聯性,差異顯著(P<0.05)。
2 結果與分析
2.1 雞IGF1R基因多態性分析結果
使用MegAlign分析雞IGF1R基因多態性,發現在雞IGF1R基因的第3外顯子(Exon-3)有3個SNPs位點,即g.105384 C>T、g.105333 A>C和g.110681 A>C。3個SNPs位點均存在3種基因型,突變點為同義突變,編碼氨基酸未改變。見圖1、圖2、圖3。
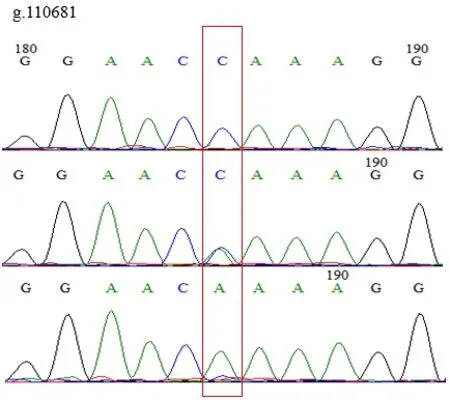
圖1 g.110681
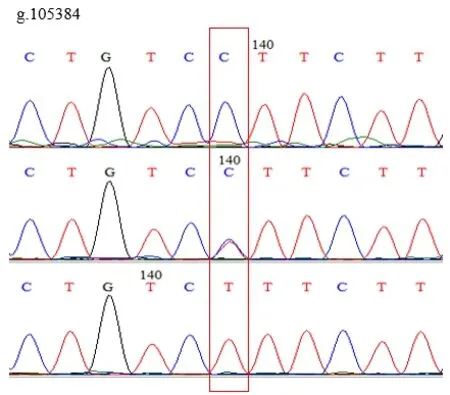
圖2 g.105384

圖3 g.105333
2.2 雞IGF1R基因群體遺傳特性分析
雞IGF1R基因群體遺傳特性分析見表2。g.105384 C>T、g.105333 A>C和g.110681 A>C這3個SNP位點均為中度多態(0.25<PIC<0.5);優勢基因型分別為CT、AC和AC,頻率均是0.580;優勢等位基因和頻率分別為C(0.647)、C(0.647)、A(0.735)。χ2檢驗結果顯示,g.105384 C>T、g.105333 A>C和g.110681 A>C均未偏離Hardy-Weinberg平衡(P>0.05)。

表2 SNP位點群體遺傳信息分析
2.3 雞IGF1R基因SNP單倍型和雙倍型分析結果
在線軟件SHEsis(http://analysis.bio-x.cn/)構建單倍型和雙倍型。結果顯示(表3),在試驗沿河鐵叫雞群體中,存在4種單倍型(H1,H2,H3,H4),9種雙倍型(H1H1、H1H2、H1H3、H1H4、H2H2、H2H4、H3H3、H3H4和H4H4)。優勢單倍型H1(73),頻率0.365,其次是H4(67),頻率0.335;優勢雙倍型為H1H4(40),頻率0.400,其次是 H1H3(12)和H3H4(12)頻率均為0.120。
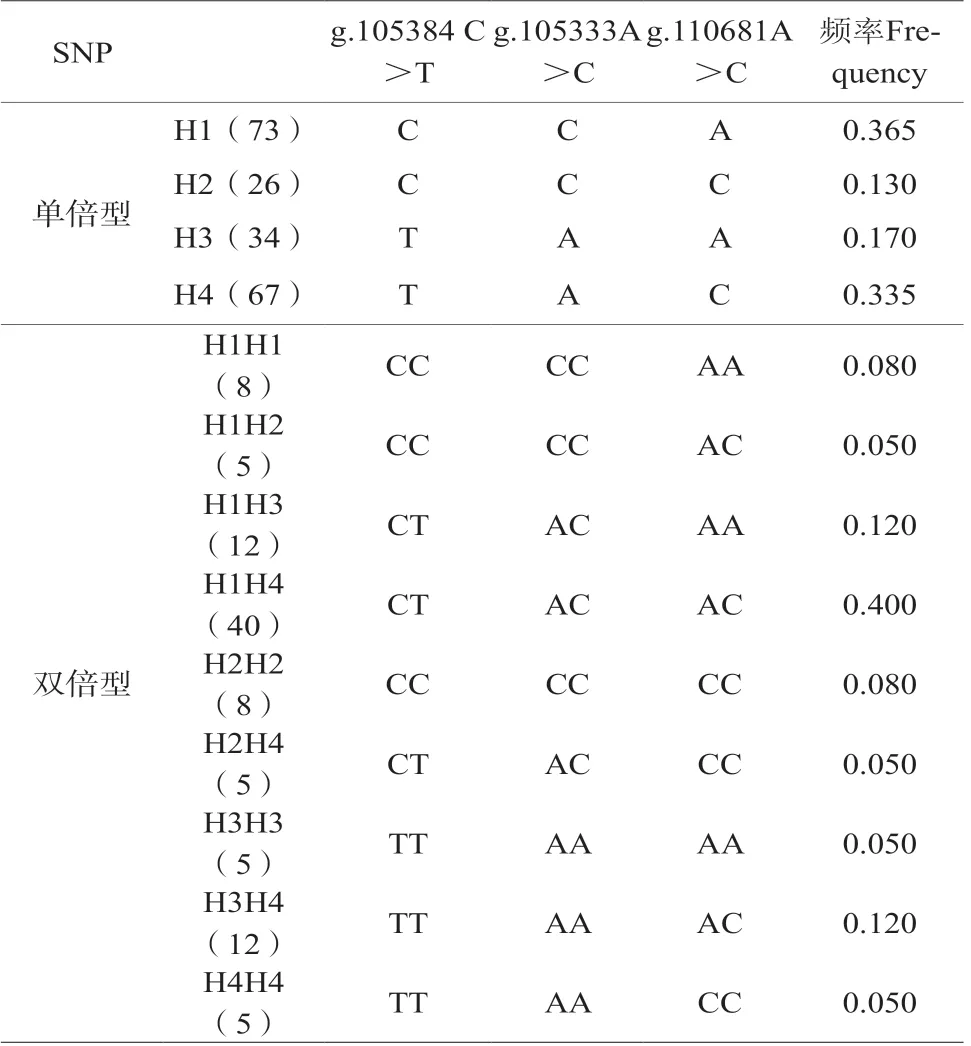
表3 SNPs的單倍型和雙倍型分析
2.4 雞IGF1R基因SNP與體尺的關聯分析結果
一般線性模型(GLM)分析了IGF1R基因多態與體尺的關聯性(表4)。結果顯示,g.105384 C>T突變位點CC和CT基因型雞的體斜長顯著大于CC基因型(P<0.05);g.105333 A>C位點AA和AC基因型體斜長顯著大于CC基因型(P<0.05),g.110681 A>C位點AA基因型髖骨寬顯著大于AC和CC基因型(P<0.05)。

表4 IGF1R基因多態與體尺的關聯性分析
2.5 雞IGF1R基因SNP與屠宰性能的關聯分析結果
試驗分析IGF1R基因多態與體尺的關聯性,詳見表5。結果表面,g.105384 C>T位點TT基因型半凈膛和全凈膛重顯著高于CC基因型(P<0.05),g.105333 A>C位點AA基因型半凈膛和全凈膛重顯著高于CC基因型(P<0.05),g.110681 A>C突變位點CC基因型腿肌重顯著高于AA和AC基因型。
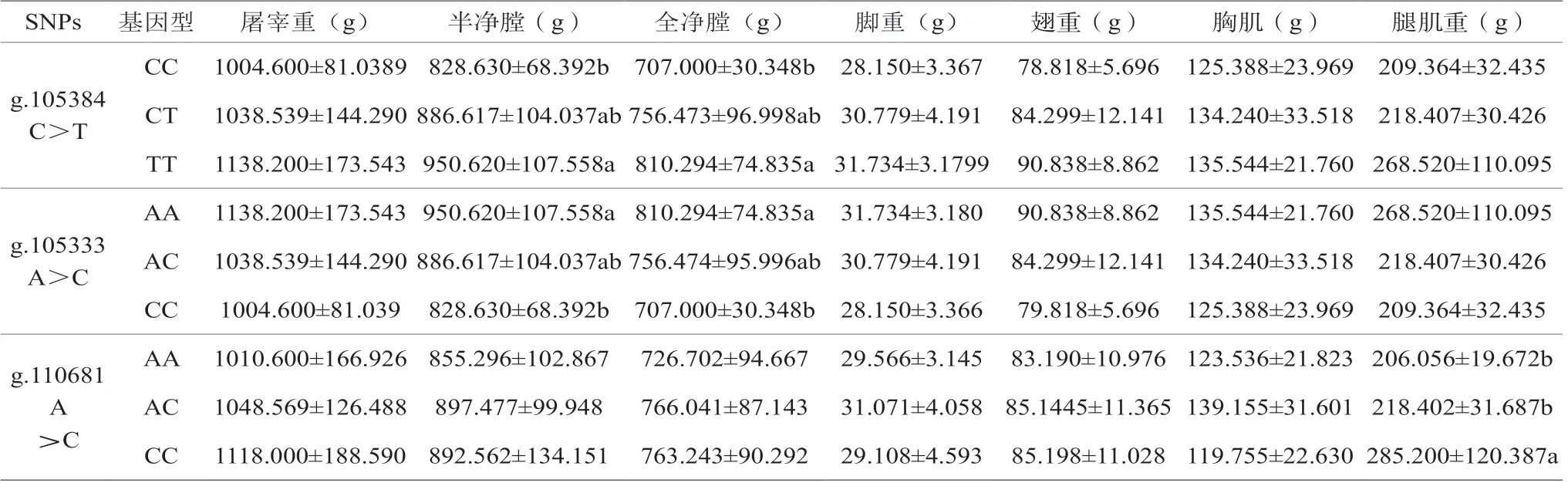
表5 IGF1R基因多態與屠宰性狀的關聯性分析
3 討論
IGF1R基因已被研究為多種動物體尺和胴體性狀的功能候選基因。在水牛、牦牛、山羊、豬和雞身上發現了與許多這些特性的顯著關聯[9-12]。雖然在雞方面,高鳳華等[13]研究IGF1R基因多態性對生長性狀的影響,但雞的衍生品種中進行研究,同時IGF1R基因多態性與雞屠宰性狀的關聯相對較少。Ekegbu[14]檢測了新西蘭羅姆尼綿羊IGF1R基因從9號外顯子到10號內含子片段中核苷酸變異,并發現其影響羊的屠宰性狀。而Byun等[15]發現了綿羊IGF1R基因核苷酸變異與壽命之間的關聯。鑒于上述IGF1R基因的生物學功能,并且IGF1R基因與許多其他候選基因一樣,對多個性狀具有多效性作用[16]。在這種情況下,雞的生長、胴體和肌肉質量的改善也可能伴隨著肉質的改善。因此,廣泛和復雜的研究,包括廣泛的生產性狀,有可能提供適當的價值的分子標記育種。
研究對IGF1R基因跨越部分內含子和全部外顯子的片段進行多態性分析,3個SNP位點g.105384 C>T、g.105333 A>C和g.110681 A>C分別在IGF1R基因的外顯子3被檢測到,3個SNPs基因型分布符合Hardy-Weinberg平衡。某個群體未來世代的基因型和等位基因頻率,當前種群的平衡狀態常用Hardy-Weinberg平衡預測,當一個符合孟德爾群體之間交配隨機,并且無選擇、突變、遷移和遺傳漂變等事件發生時,那么基因頻率將保持不變,即這個群體被稱為處于Hardy-Weinberg平衡[17]。根據關聯分析法,3個SNPs可聯合組成8種單倍型和36雙倍型,但本試驗只找到了4種單倍型和9種雙倍型,與理論有差距,可能與試驗群體小或其他單倍體個體在人工選育中被淘汰有關。
關聯分析結果顯示g.105384 C>T突變位點CC和CT基因型雞的體斜長顯著大于CC基因型(P<0.05);g.105333 A>C位點AA和AC基因型體斜長顯著大于CC基因型(P<0.05),g.110681 A>C位點AA基因型髖骨寬顯著大于AC和CC基因型(P<0.05)。g.105384 C>T位點TT基因型半凈膛和全凈膛重顯著高于CC基因型(P<0.05),g.105333 A>C位點AA基因型半凈膛和全凈膛重顯著高于CC基因型(P<0.05),g.110681 A>C突變位點CC基因型腿肌重顯著高于AA和AC基因型。前人研究證實,同義突變SNP在IGF1R基因中的雖不直接改變蛋白的一級序列[18]。然而,它可以引起IGF1R蛋白表達、構象和功能的改變。而在羊中,3號外顯子核苷酸c.654G > A突變,它編碼了IGF1R的尿道亞基的一部分,子亞基包含1個富含半胱氨酸的區域,參與配體結合。因此,該區域的任何變異,例如由IGF1R基因特定片段的核苷酸替換引起的變異,可能會影響IGF1R與IGF1或胰島素的相互作用,從而導致信號通路的改變。在綿羊IGF1R基因中,同義突變,c.654G > A突變可擾亂轉錄、剪接、共翻譯折疊、mRNA穩定,從而導致其他功能相關的變化[19]。此外,同義替換可以影響蛋白質編碼區域內的轉錄和剪接調節因子,從而調節基因表達。此外,最近研究揭示了非同義編碼SNP和同義編碼SNP在性狀方面表現出相似的似然性和效應大小。g.105384 C>T、g.105333 A>C和g.110681 A>C位點一些基因型對雞的體尺和屠宰性狀有顯著影響可能與此相關,不可忽視。在今后的選育培養過程中可根據實際需求選擇培育。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
