基于線蟲生物多樣性的稻田鎘污染土壤修復成效研究
2022-10-19 14:28:08鄧普榮劉勇波
環境科學研究 2022年10期
關鍵詞:污染
鄧普榮,姚 志,劉勇波
中國環境科學研究院,國家環境保護區域生態過程與功能評估重點實驗室,北京 100012
近年來,農田土壤重金屬污染問題備受關注.農田的重金屬積累使得土壤肥力下降、土壤退化和可利用耕地面積減少[1-2],且受污染農田的重金屬通過食物鏈在植物、動物和人體內富集,威脅人體健康[3].為修復重金屬污染的耕地,已采用物理、化學、生物和農業生態修復等技術方法,如客土法、熱解吸法、土壤淋洗、植物和微生物修復、調整農田耕作制度等一系列農田重金屬修復技術[4-5].通過施用鈍化劑吸附、沉淀或絡合重金屬被廣泛用于農田重金屬的修復,降低重金屬的流動性和生物利用率,從而減少重金屬在食物鏈上的傳遞[6].
土壤重金屬污染的修復技術發展日漸成熟,但對受污染農田的修復后評價,特別是針對土壤動物的群落特征及種群動態的監測常常被忽視[7].20 世紀80年代,荷蘭線蟲學家Bongers[8]將土壤線蟲作為環境指示生物應用到污染土壤監測及恢復過程中.土壤線蟲是土壤動物的一個重要類群,其種類繁多,分布廣泛,參與土壤中有機質分解、植物營養礦化和養分循環等生態過程,在土壤生態系統中占有重要地位[9-10].土壤線蟲的物種多樣性、食性多樣性和生活史策略多樣性使其在變動的環境因子中,尤其是污染脅迫中作出迅速反應[11-12].土壤線蟲不同營養類群的組成、數量及線蟲群落結構的動態分析可直接反映土壤物質的代謝途徑和食物鏈/食物網結構的變化,指示土壤生態系統中物質和能量的流動情況,進而在生態系統功能水平上揭示土壤環境健康狀態[13-16].
該文在鎘污染稻田土壤中采用施加鈍化劑、鈍化劑+凈水和噴施葉面肥三種修復措施,分析水稻農田生態系統中土壤線蟲的物種組成、豐富度、群落結構、食物網動態及群落的結構變化,評價修復措施對Cd 污染稻田土壤修復的生態效應,以期確定能否采用土壤線蟲多樣性指數來指示農田重金屬污染的修復成效.
1 材料與方法
1.1 試驗地概況
試驗地位于湖南省長沙市長沙縣農田(28°15′N、113°14′E),占地面積約0.37 hm2.表層土壤中Cd 的濃度范圍為0.40~0.80 mg/kg,超過《土壤環境質量標準》(GB 15618—2018)農用地土壤Cd 污染風險篩選值(0.4 mg/kg),而其他重金屬含量均低于GB 15618—2018 相關限值.
試驗田種植了早稻“中早39”和晚稻“玉針香”,水稻種子由湖南省農業科學院提供.育苗前用5%的次氯酸鈉溶液消毒15 min,用自來水沖洗.稻田其他管理遵循長沙縣當地水稻常規操作流程.
1.2 土壤修復技術與線蟲取樣方法
該試驗采用3 種土壤修復方法,即施用鈍化劑、鈍化劑+凈水、噴施葉面肥[17].鈍化劑的主要成分是赤泥、磷礦粉、海泡石,三者質量比為2∶1∶1;使用時按1 kg 農田土添加2 g 鈍化劑(包括赤泥0.10%、磷礦粉0.05%、海泡石0.05%),即鈍化劑含量為0.2%;以1 hm2農田施用300 kg 鈍化劑的比例向農田中施用鈍化劑.葉面肥中1.0 g/L 的硅來自Na2SiO3,0.5 g/L的磷來自NaH2PO4,1.5 g/L 的硒來自Na2SeO3,0.5 g/L的硫來自Na2S,0.5 g/L 鎂的來自MgCl2.3 種土壤修復方法分別在3 塊農田中進行,并設置蒸餾水噴施作為對照組(CK),共4 種處理,每種處理均設3 個重復地塊,地塊面積40~135 m2.在水稻種植的苗期(25~35 d)、分蘗期(60~70 d)和開花期(80~90 d)分別施用鈍化劑和噴施葉面肥.
于5 月22 日、7 月16 日、9 月23 日和10 月22 日在長沙縣試驗地收集土壤樣品,為隨機取樣.每次在各地塊中的3 個樣點采集3 份重復土壤,取樣深度為0~30 cm.利用重糖離心法分離土壤線蟲,稱取100 g土樣,放入1 000 mL 燒杯內,加水至300 mL,浸泡30 min;過60 目(0.25 mm)、100 目(0.15 mm)和500 目(0.025 mm)篩,收集篩上的線蟲和卵,倒入離心管中離心(分離線蟲,3 000 r/min,離心5 min;分離卵,3 000 r/min,離心3 min);把上清液倒掉,加蔗糖(分離線蟲加70%的蔗糖液,分離卵加35%~40%的蔗糖),離心(同上),過500 目(0.025 mm)篩,收集篩上的線蟲和卵,在解剖鏡下記數并鑒定到屬[18].
1.3 生物多樣性指數和生態功能性指數計算
1.3.1 土壤線蟲生物多樣性指數
物種多樣性指數選用多樣性指數、優勢度指數、均勻度指數和豐富度指數.
Shannon-Wiener 多樣性指數(H′):
Simpson 優勢度指數(C):
Pielou 均勻度指數(E):
SR 豐富度指數:
式中,Pi為第i種線蟲的個體數占調查個體總數的比例,N為所調查到的總個體數,S為所調查的物種總數.
1.3.2 土壤線蟲生態功能性指數
生態功能指數選用成熟指數(MI 指數)、植物寄生線蟲成熟指數(PPI 指數)、線蟲富集指數(EI 指數)、線蟲結構指數(SI 指數)、線蟲通路比值(NCR 值)和瓦斯樂斯卡指數(WI 指數).
MI 指數:
式中,Cpi為第i種線蟲的c-p 值(colonizer-persister value),c-p 值根據自由生活線蟲在生態演替中的不同生活策略,分別賦值為1~5.根據線蟲在r 對策到k 對策的生活史策略連續譜中的位置,將線蟲劃分為5 個c-p 類群:c-p 1 為典型的機會主義者,世代時間極短,卵量巨大,在食物豐富的條件下種群暴發,極耐污染和環境壓力;c-p 2 也是機會主義者,世代時間短,卵量大,較耐污染和環境壓力;c-p 3 世代時間較長,對環境壓力較敏感;c-p 4 世代時間長,對環境壓力敏感;c-p 5 則為典型的k 對策者,世代時間很長,卵量小,對污染和環境壓力極為敏感[8,19].
PPI 指數:
式中,Pi′為植物寄生性線蟲在線蟲種群中的占比,m為所調查植物寄生性線蟲的物種總數.
MI 指數和PPI 指數反映土壤線蟲群落功能結構特征,用以評價人類干擾活動對土壤線蟲群落的影響,MI 指數反映自由生活線蟲的成熟度,PPI 指數反映植物寄生線蟲的成熟度,較低的MI 指數表明土壤生態系統擾動強烈,反之則表明土壤生態系統趨于穩定.
MI25 指數:
式中,Cpi25為c-p 值在2~5 之間的第i種線蟲的c-p值,Pi25為第i種線蟲的個體數占總個體數的比例,MI25 為c-p 值在2~5 之間的自由生活線蟲的總成熟指數,n為c-p 值在2~5 之間線蟲的物種總數.
EI 指數:
式中:EI 代表土壤線蟲富集指數;根據線蟲食性并結合線蟲生活史特征,將線蟲分為若干類群〔Bax、Fux、Ppx和Omx分別代表食細菌、食真菌、植食性以及雜食/捕食線蟲(x取值為1~5,代表線蟲的c-p 值)[20]〕,b代表食物網中的基礎成分,主要指Ba2和Fu2這兩個類群;e代表食物網中的富集成分,主要指Ba1和Fu2這兩個類群;kb和ke為各類群所對應的加權數(在0.8~5.0 之間);nb和ne則為各類群的豐度.
SI 指數:
式中:SI 為結構指數;s代表食物網中的結構成分,包括Ba3~Ba5、Fu3~Fu5、Om3~Om5和Pp2~Pp5類群;ks為各類群所對應的加權數;ns為各類群的豐度.
NCR 值:
式中:NBa為食細菌線蟲數量,個;NFu為食真菌線蟲數量,個.
WI 指數:
式中,NPp為植食性線蟲數量,個.
EI 指數和SI 指數均在0~100 之間變化,二者結合分析可以較好地指示土壤環境的干擾程度和土壤食物網的變化.EI 指數主要用于評估食物網對可利用資源的響應,SI 指數指示在干擾或恢復過程中土壤食物網結構的變化.NCR 值主要用于指示土壤食物網的降解途徑[14].
2 結果與討論
2.1 土壤線蟲群落組成
土壤線蟲分析數據如表1 所示,共鑒定出線蟲13 599 條,隸屬于32 屬,包括食細菌線蟲13 屬,占14.62%;食真菌線蟲3 屬,占1.72%;植食性線蟲8 屬,占22.10%;雜食/捕食線蟲8 屬,占61.55%.矛線屬(Dorylaimus)、潛根屬(Hirschmanniella)和托布利屬(Tobrilus)為此水稻農田生態系統的優勢屬.
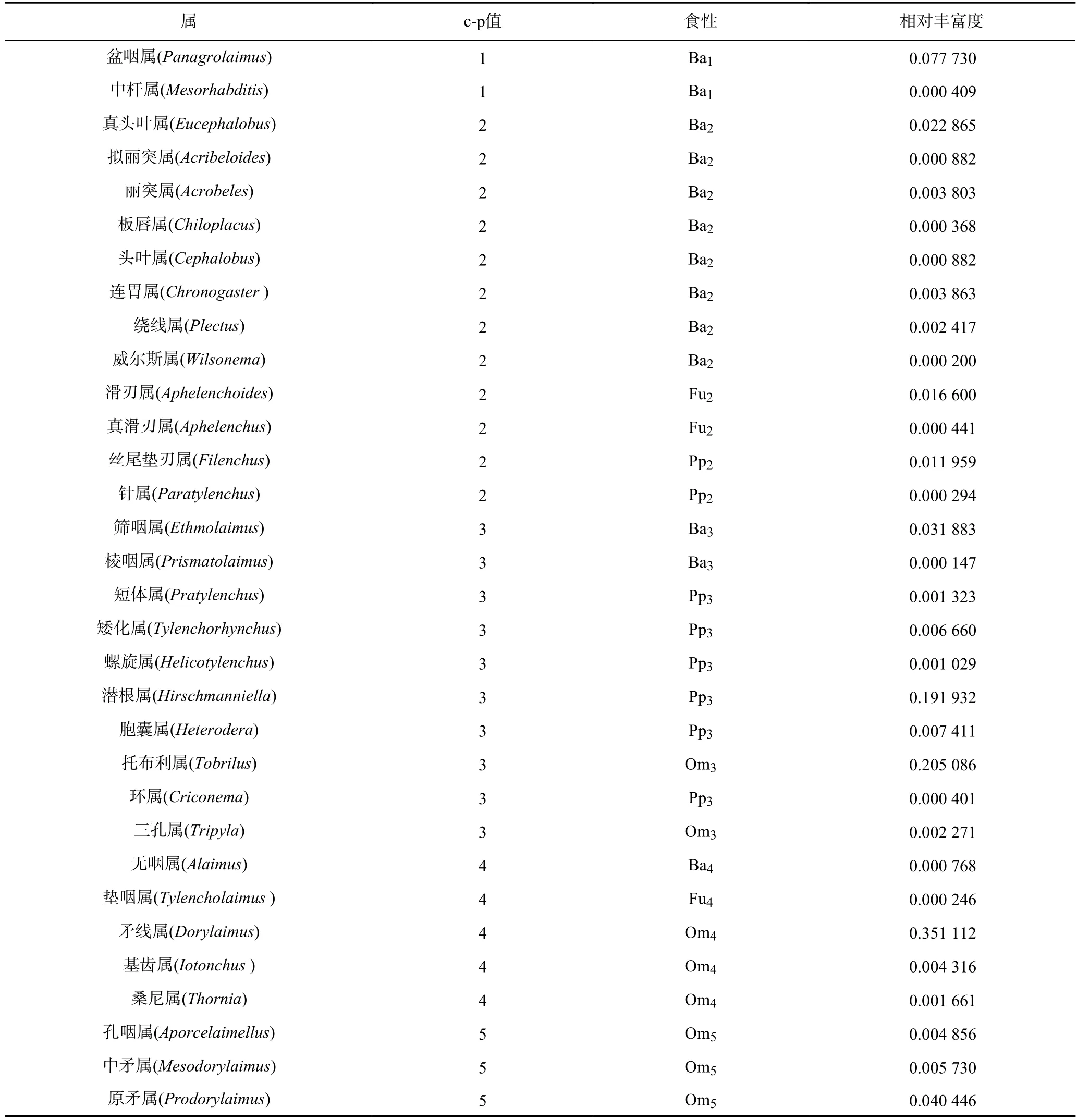
表1 試驗稻田樣地土壤線蟲群落組成Table 1 Community composition of soil nematode in paddy fields
2.2 Cd污染土壤修復措施下線蟲群落多樣性的變化
土壤線蟲的豐富度、物種數、生物多樣性指數等在不同取樣時間表現出顯著差異(見表2).不同修復措施對水稻土壤線蟲物種數、Shannon-Wiener 多樣性指數、Simpson 優勢度指數、Pielou 均勻度指數和SR 豐富度指數均沒有顯著影響,但對土壤線蟲豐富度有顯著影響(P<0.05,見表2、圖1、圖2).
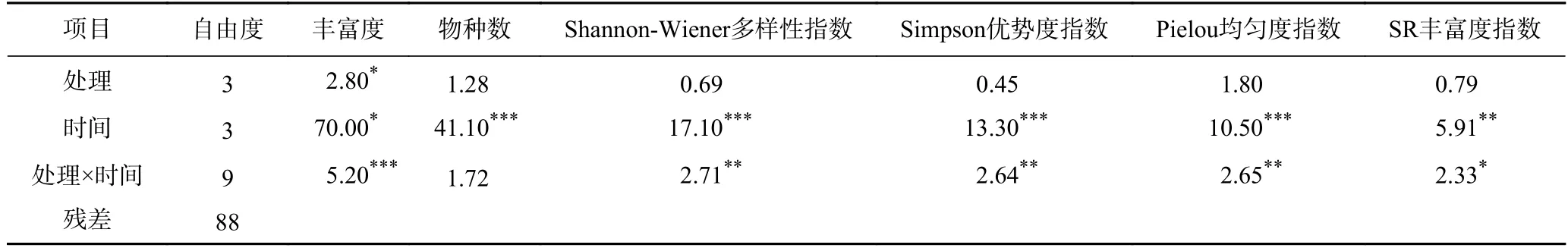
表2 3 種修復措施下稻田土壤線蟲的生物多樣性指數方差分析結果Table 2 F-values of ANOVAs for biodiversity indices of soil nematodes under three remediation methods at four sampling times in paddy fields
線蟲豐富度的影響與取樣時間有交互作用(見表2).多重比較分析不同月份3 種土壤修復措施下稻田土壤中線蟲豐富度發現,在9 月和10 月線蟲豐富度差異不顯著,但在5 月和7 月差異顯著(見圖1).5 月,鈍化劑處理的土壤中線蟲豐富度最高,顯著高于鈍化劑+凈水和葉面肥處理,但與對照組沒有顯著性差異.7 月,葉面肥處理的水稻土壤中線蟲豐富度最高,顯著高于鈍化劑處理的水稻土壤,但與鈍化劑+凈水處理和對照組差異不顯著(見圖1).
2.3 Cd 污染土壤修復措施下土壤線蟲生活史策略指數的變化
不同修復措施對水稻農田土壤線蟲生活史策略指數—MI 指數、PPI 指數、MI25 指數等均有顯著影響(見表3).土壤線蟲MI 指數在不同處理之間有顯著性差異(見表3),多重比較分析發現,在5 月和9 月沒有顯著差異,但在7 月和10 月有顯著差異,其中鈍化劑+凈水處理的土壤中線蟲MI 指數最高,顯著高于葉面肥處理的土壤,但與鈍化劑處理和對照組的土壤均無顯著差異(見圖3).
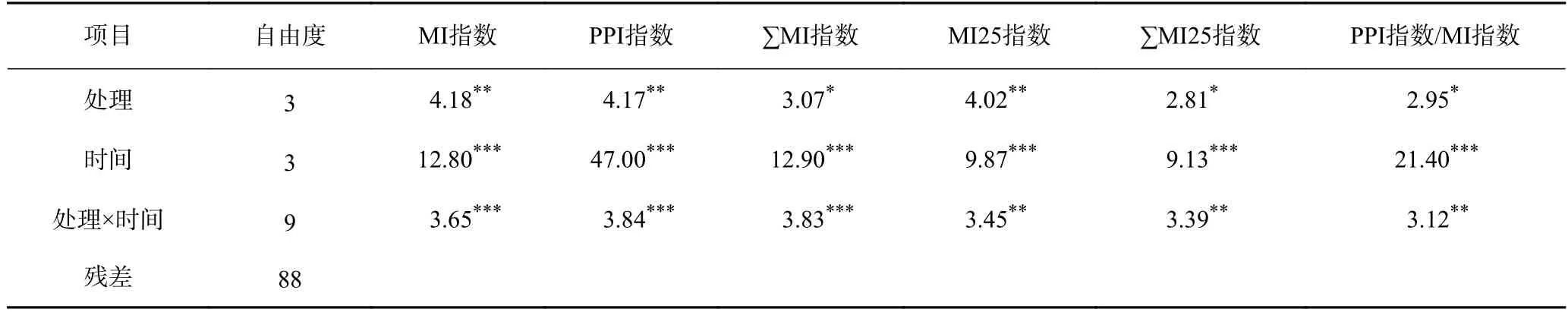
表3 3 種修復措施下稻田土壤線蟲的生活史策略指數方差分析結果Table 3 F-values of ANOVAs for life history strategy indices of soil nematodes under three remediation methods at four sampling times in paddy fields
植物寄生線蟲PPI 指數在不同處理之間具有顯著差異(見表3),多重比較分析發現,在5 月、9 月和10 月差異不顯著,但在7 月有顯著差異(見圖3).7 月,葉面肥處理的土壤中線蟲PPI 指數顯著低于鈍化劑+凈水處理的土壤,但與鈍化劑處理和對照組的土壤均無顯著差異(見圖3).
由圖3 可見:5 月,使用鈍化劑處理的土壤中線蟲MI 指數最低,顯著低于對照組土壤,但與鈍化劑+凈水及葉面肥處理的土壤之間沒有顯著差異;7 月和10 月,葉面肥處理的土壤中線蟲MI 指數最低,顯著低于鈍化劑+凈水處理的土壤,但與對照組和鈍化劑處理的土壤均無顯著差異(見圖3).
c-p 值在2~5 之間的自由生活線蟲成熟指數(MI25 指數)能指示土壤狀況的變化.由圖3 可見,在7 月和10 月,不同處理間稻田土壤線蟲群落MI25 指數有顯著差異,但在5 月和9 月無顯著差異.在7 月和10 月,葉面肥處理的土壤中線蟲MI25 指數最低,顯著低于鈍化劑+凈水處理的土壤,但與對照組及鈍化劑處理的土壤均沒有顯著差異.
2.4 Cd 污染土壤修復措施下線蟲生態功能指數的變化特征
不同土壤修復措施下線蟲的生態功能指數(EI指數、SI 指數、NCR 指數和WI 指數)均沒有顯著差異(見表4 和圖4).NCR 指數接近0,代表土壤有機質分解依靠真菌分解途徑;若NCR 指數接近1,代表土壤有機質分解依賴細菌分解途徑[21].所有樣地的NCR 指數在0.6~1.0 之間,表明細菌是農田土壤有機質的主要分解分解者.該農田多數土壤樣品的WI 指數>1,表明土壤健康程度較好.
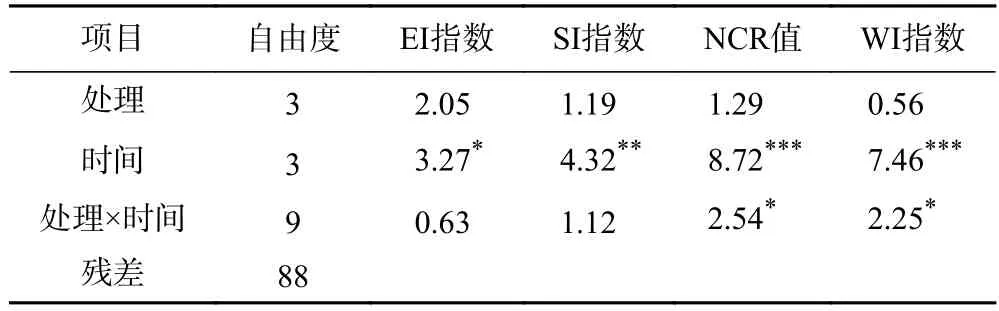
表4 3 種修復措施下稻田土壤線蟲的生態功能性指數方差分析結果Table 4 F-values of ANOVAs for ecological functions of soil nematodes under three remediation methods at four sampling times in paddy fields
EI 指數與SI 指數聯合使用時,以SI 指數(0~100)為橫坐標、EI 指數(0~100)為縱坐標,以(50,50)為中心,將SI 指數和EI 指數的分布劃分成4 個象限,這4 個象限分別代表食物網不同的特征.SI 指數指示土壤食物網連通性和食物鏈長度,其數值越大,表明相對連通性越高,食物鏈越長.EI 指數指示外界養分投入狀況,其數值越大,表明外界投入養分相對越多.由圖5 可見,在3 種稻田重金屬Cd 的修復措施下,各處理的土壤線蟲權重區系分析結果分布于B 和C 象限.分布于B 象限表明該水稻農田養分狀況較好且受脅迫程度較低,食物網穩定成熟;分布在C 象限表明水稻農田肥力狀況較差但受脅迫程度較低[22].
2.5 討論
土壤線蟲食性多樣,占據食物鏈/食物網多個營養級,是土壤生態系統中的重要組成部分.土壤線蟲因種類眾多、繁殖力強、遷移能力差和對環境脅迫敏感,常作為評價土壤質量的指示生物[23-24].筆者試驗研究了Cd 污染稻田在3 種修復措施下,土壤線蟲的多樣性及土壤線蟲群落的結構和功能的變化,發現土壤線蟲的豐富度、物種數、生物多樣性指數和生態功能指數等在不同取樣時間上有顯著性差異.Yang等[25]對連續種植多年的轉Bt基因水稻農田觀測發現,取樣年份和采樣日期對根際土壤中線蟲群落組成的影響比轉Bt基因水稻處理更明顯.這也證明土壤線蟲隨環境因子變化而變化,適合指示土壤理化性質、土壤肥力、土壤食物網結構和功能等動態變化[19,26].一般采用多種物理、化學和生物技術方法修復重金屬Cd 污染土壤,如使用鈍化劑和葉面肥等,這些可能改變土壤環境,從而影響土壤線蟲群落.鈍化劑通過升高稻田土壤pH 從而降低重金屬Cd 從土壤到水稻的遷移率[27],也能影響農田中線蟲的群落結構和動態[28].噴施混合磷硫葉面肥影響地上部分作物的次生代謝產物進而影響土壤線蟲群落[17].
鈍化劑和鈍化劑+凈水的噴施均顯著增加了稻田線蟲成熟指數,且c-p 3、c-p 4、c-p 5 的線蟲類群占據優勢,該措施下農田生態系統受到的干擾減小,向著更成熟的階段發展.葉面肥噴施處理部分降低了土壤線蟲的MI 指數,增加了PPI 指數,且c-p 2~5 類群線蟲的MI25 指數為所有處理間最低,這說明地上植被與土壤線蟲群落之間為聯動的系統.植被-土壤-土壤線蟲之間存在的互動效應[29].Zhang 等[30]以森林生態系統為例,證明森林地上植被的種群結構會影響地下土壤線蟲多樣性和種群結構的維持.Hu 等[31]以高山草甸生態系統為例,證明放牧強度對土壤線蟲群落有顯著影響.葉面肥的噴施一定程度上促進了農田地上水稻作物的生長,進而影響了土壤線蟲群落結構[32].在農田生態系統中,地上部分農作物的耕作也影響著土壤線蟲的豐度.在試驗中隨著水稻的收獲,土壤線蟲豐度于7 月和10 月達到了峰值.水稻的收獲使有機質在稻田土壤中積累,特別是在7 月,土壤線蟲豐富度、物種數和Shannon-Wiener 多樣性指數均達到峰值.Zhu 等[32-34]證明,適量的有機肥或秸稈還田都增加了土壤線蟲的多樣性,改變了土壤線蟲群落的組成.
3 種修復措施對水稻土壤線蟲的生態功能指數均沒有顯著影響.Park 等[35]對0~2.6 mg/kg 濃度梯度下Cd 污染土壤的短期生態效應評估發現,Cd 對生態指數影響顯著,這可能與土壤Cd 的含量相關.但在試驗中鈍化劑和葉面肥的使用導致線蟲NCR 指數降低,有機物分解途徑向真菌分解轉移,而WI 指數增加,土壤健康狀況向好發展[36].氮和磷的有效性限制了生態系統的生產力,土壤養分的有效性和碳氮比是稻田土壤線蟲群落的主要調控因子[9,36].這種養分驅動對土壤微生物和線蟲的影響可能超過了重金屬有效性的負面影響[34,36].
土壤管控模式影響了土壤線蟲群落種群動態.例如,農田耕作方式、土壤有機肥使用和殺蟲劑噴施等均會對土壤線蟲群落多樣性和結構產生影響[37-43].在使用土壤鈍化劑的農田管理模式下,農田生態系統趨于穩定,且土壤健康狀況得到改善,因此鈍化劑可用于微量Cd 重金屬超標稻田管控[5].土壤線蟲群落動態研究對監測農業生態系統、改善農田營養結構、提高人類干預恢復和保護耕地具有重要意義.
3 結論
a) 土壤線蟲的豐富度、物種數、生物多樣性指數和生態功能指數等在不同時間有顯著性差異,說明土壤線蟲對環境因素較敏感,適合用于指示土壤理化性質、土壤肥力、土壤食物網結構和功能等變化.
b) 3 種修復措施對水稻土壤線蟲豐富度有顯著影響,不同試驗處理對水稻土壤線蟲Shannon-Wiener多樣性指數、Simpson 優勢度指數、Pielou 均勻度指數的影響與取樣時間有交互作用.
c) 3 種修復措施對水稻土壤線蟲的生態功能指數—EI 指數、SI 指數、NCR 值和WI 指數均沒有顯著影響,說明這3 種重金屬土壤修復措施對土壤線蟲群落的結構和功能沒有顯著影響.
d) Cd 污染修復措施對水稻土壤線蟲生活史策略指數系列(MI 指數、PPI 指數、MI25 指數等)有顯著影響.MI 系列指數為線蟲群落的加權c-p 值,較好地指示了土壤生態系統的演替和恢復等過程,說明土壤重金屬修復模式顯著影響了土壤線蟲群落的生態系統演替.土壤線蟲生活史策略指數可以作為重金屬污染土壤修復的成效評價指標.
猜你喜歡
小學科學(學生版)(2021年8期)2021-08-30 09:16:18
小學科學(學生版)(2021年7期)2021-07-28 06:44:32
小學科學(學生版)(2020年6期)2020-08-13 07:47:12
當代陜西(2019年7期)2019-04-25 00:22:18
領導決策信息(2018年26期)2018-10-12 02:18:26
新高考(英語進階)(2017年10期)2017-12-23 09:14:50
食品界(2016年4期)2016-02-27 07:36:15
都市麗人(2015年5期)2015-03-20 13:33:49
裝備環境工程(2015年4期)2015-02-28 01:20:10
江蘇年鑒(2014年0期)2014-03-11 17:09:46
