廣東花生叢枝病植原體的分子鑒定
2022-10-13 11:29:38湯亞飛佘小漫李正剛藍(lán)國(guó)兵何自福
植物保護(hù) 2022年5期
關(guān)鍵詞:一致性
湯亞飛, 林 祺, 佘小漫, 李正剛, 于 琳, 藍(lán)國(guó)兵, 何自福*
(1. 廣東省農(nóng)業(yè)科學(xué)院植物保護(hù)研究所, 廣東省植物保護(hù)新技術(shù)重點(diǎn)實(shí)驗(yàn)室, 廣州 510640;2. 華南農(nóng)業(yè)大學(xué)植物保護(hù)學(xué)院, 廣州 510642)
植原體phytoplasma是一類專性寄生、不能人工培養(yǎng)的植物病原物,隸屬于細(xì)菌界Bacteria軟壁菌門Tenericutes柔膜菌綱Mollicutes無(wú)膽甾原體目Acholeplasmatales無(wú)膽甾原體科Acholeplasmataceae植原體暫定屬Ca.genus Phytoplasma[1-2]。植原體主要通過(guò)具有刺吸式口器的昆蟲傳播,也可通過(guò)嫁接或菟絲子傳播,寄主范圍廣泛,可侵染1 000多種植物,常引起叢枝、花變?nèi)~、黃化、小葉、衰退、矮化等典型癥狀,給農(nóng)林業(yè)生產(chǎn)造成較大的損失[3]。中國(guó)已報(bào)道了100多種植原體相關(guān)病害[4],如棗瘋病(jujube witches’-broom disease)[5-6]、小麥藍(lán)矮病(wheat blue dwarf disease)[7-8]、馬鈴薯僵頂病(potato stolbur disease)[9]、櫻桃叢枝或花變?nèi)~病(cherry witches’-broom or phyllody disease)[10-11]等。國(guó)際上,依據(jù)16S rRNA 基因核苷酸序列一致性、限制性片段長(zhǎng)度多態(tài)性(restriction fragment length polymorphism,RFLP)結(jié)果對(duì)植原體進(jìn)行分類,目前植原體包括52個(gè)暫定種(‘CandidatusPhytoplasma’),34個(gè)組(group)和100多個(gè)亞組(subgroup)[3]。
花生是我國(guó)重要的油料和經(jīng)濟(jì)作物之一,是廣東省第二大農(nóng)作物,全省種植面積常年保持在30萬(wàn)~35萬(wàn)hm2,僅次于水稻[12]。花生叢枝植原體(peanut witches’-broom phytoplasma,PnWB)引起花生叢枝病[13],在我國(guó)南方花生產(chǎn)區(qū)發(fā)生較為普遍。花生叢枝病田間主要表現(xiàn)為植株節(jié)間縮短,產(chǎn)生大量腋芽,叢生枝條上葉片變小,大多數(shù)植株不接莢果,民間俗稱“花生公”,嚴(yán)重影響花生的產(chǎn)量。
我國(guó)花生叢枝病于1952年首次在廣州發(fā)現(xiàn)[14]。該病害被發(fā)現(xiàn)以來(lái),我國(guó)不少學(xué)者對(duì)其開展了相關(guān)研究,20世紀(jì)80年代初,通過(guò)昆蟲介體傳病試驗(yàn)證明華南花生叢枝病是由小綠葉蟬EmpoascaflavescensFab.傳播的類菌原體(現(xiàn)稱植原體)病[15];同年,通過(guò)電子顯微鏡從海南花生叢枝病病樣中觀察到類菌原體[16]。1994年,對(duì)華南花生叢枝病的發(fā)病規(guī)律及防治方法開展了相關(guān)研究[14]。2000年,開展了花生叢枝病病原的血清學(xué)研究[17]。隨著分子生物學(xué)技術(shù)的發(fā)展,不少學(xué)者已對(duì)引起海南、云南、臺(tái)灣等省花生叢枝病的相關(guān)植原體的16S rRNA、SecY、rp等多個(gè)基因進(jìn)行克隆和序列分析[13,18-21]。尚未見(jiàn)廣東花生叢枝病的植原體分子特征及相關(guān)基因序列分析的報(bào)道。本研究采用分子生物學(xué)方法對(duì)廣東采集的花生叢枝病進(jìn)行了病原鑒定,明確了植原體株系分類地位,為進(jìn)一步開展花生植原體病害防治提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
1.1.1樣品采集
2020年10月,在廣東省湛江市遂溪縣花生種植地隨機(jī)采集了7份花生叢枝病樣,病株田間癥狀表現(xiàn)典型叢枝、小葉(圖1)。

圖1 廣東省花生叢枝病田間癥狀Fig.1 Symptoms of peanut witches’-broom disease in fields in Guangdong province
1.1.2主要試劑和儀器設(shè)備
植物基因組DNA抽提試劑盒(EasyPure Plant Genomic DNA Kit)和大腸桿菌EscherichiacoliT1感受態(tài)細(xì)胞購(gòu)自北京全式金生物技術(shù)有限公司;PremixTaqTM(ExTaqTMVersion 2.0 plus dye)和載體pMD 19-T購(gòu)自寶生物工程(大連)有限公司;瓊脂糖凝膠回收試劑盒(GeneJET Gel Extraction Kit)購(gòu)自美國(guó)Thermo(賽默飛)公司;氨芐青霉素、X-Gal、蔗糖、瓊脂糖和胰蛋白胨購(gòu)自生工生物工程(上海)股份有限公司;GoldView核酸染料購(gòu)自湖南艾科瑞生物工程有限公司;其他常規(guī)分析純?cè)噭┵?gòu)自廣州市芊薈化玻儀器有限公司。T100TMThermal cycler PCR儀、Universal HoodⅡ凝膠成像系統(tǒng)以及電泳儀均購(gòu)自美國(guó)BIO-RAD公司;植物組織自動(dòng)研磨儀MM400購(gòu)自德國(guó)Retsch(萊馳)公司。
1.2 試驗(yàn)方法
1.2.1病樣總DNA提取
取待測(cè)花生病株葉片100 mg置于裝有鋼珠的2 mL圓底離心管,將其裝入植物組織自動(dòng)研磨儀的適配器,然后浸入液氮中冷卻2 min左右,取出后移至主機(jī)快速固定即可開始研磨,以頻率1 200次/min,時(shí)間為90 s,進(jìn)行振蕩破碎,結(jié)束后迅速取出粉末,按照植物DNA提取試劑盒(EasyPure Plant Genomic DNA Kit)說(shuō)明書上的步驟進(jìn)行總DNA抽提,DNA沉淀溶解于50 μL ddH2O中,于-20℃冰箱保存?zhèn)溆谩?/p>
1.2.216S rRNA基因的擴(kuò)增
利用擴(kuò)增植原體16S rRNA基因的通用引物P1/P7[22-23]、R16mF2/R16mR1[24],對(duì)待測(cè)病樣總DNA進(jìn)行PCR檢測(cè),預(yù)期擴(kuò)增目的片段大小分別為1.8 kb和1.4 kb。反應(yīng)體系25 μL:待測(cè)樣品總DNA 1 μL(約20 ng),ExTaqTMPremix 12.5 μL,上、下游引物(10 μmol/L)各 1 μL,滅菌水9.5 μL。反應(yīng)程序:95℃ 預(yù)變性4 min;95℃ 1 min,55℃ 1 min,72℃ 2 min,35個(gè)循環(huán);72℃ 延伸10 min。1%瓊脂糖凝膠電泳檢測(cè)PCR產(chǎn)物。
1.2.3SecY基因的擴(kuò)增
利用擴(kuò)增植原體SecY基因的通用引物L(fēng)15F1/MapR1和SecYF2(Ⅱ)/SecYR1(Ⅱ)[19],對(duì)待測(cè)病樣總DNA進(jìn)行巢式PCR擴(kuò)增,目的片段大小為1.7 kb。PCR反應(yīng)體系25 μL:待測(cè)樣品總DNA 1 μL (約20 ng),ExTaqTMPremix 12.5 μL,引物L(fēng)15F1/MapR1 (10 μmol/L)各1 μL,滅菌水9.5 μL。反應(yīng)程序:95℃ 預(yù)變性4 min;95℃ 1 min,48℃ 1 min,72℃ 2 min,30個(gè)循環(huán);72℃延伸10 min。巢式PCR反應(yīng)體系25 μL:第一輪PCR產(chǎn)物 1 μL,ExTaqTMPremix 12.5 μL,引物SecYF2(Ⅱ)/SecYR1(Ⅱ) (10 μmol/L)各1 μL,滅菌水9.5 μL。反應(yīng)程序:95℃ 預(yù)變性4 min;95℃ 30 s,55℃ 1 min,72℃ 90 s,35個(gè)循環(huán);72℃延伸10 min。1%瓊脂糖凝膠電泳檢測(cè)PCR產(chǎn)物。
1.2.4基因克隆與測(cè)序
采用瓊脂糖凝膠回收試劑盒(Gene JET Gel Extraction Kit)回收目的條帶,將回收的PCR產(chǎn)物連接到pMD-19T載體上。具體操作:pMD-19T 1 μL、2×solution I連接反應(yīng)緩沖液5 μL、回收DNA目的片段4 μL混勻,在16℃條件下反應(yīng)過(guò)夜。采用熱激法將連接產(chǎn)物轉(zhuǎn)化到大腸桿菌T1感受態(tài)細(xì)胞,在含100 μg/mL Amp、40 μL/mL X-Gal的LB固體培養(yǎng)基上,37℃倒置培養(yǎng)過(guò)夜;從每個(gè)平板隨機(jī)挑取3個(gè)陽(yáng)性白色單菌落送至生工生物工程(上海)股份有限公司進(jìn)行測(cè)序。
1.2.5序列分析
利用DNAStar的SeqMan對(duì)測(cè)序獲得的基因序列進(jìn)行拼接,去掉載體序列,將所得DNA序列在GenBank數(shù)據(jù)庫(kù)(http:∥www.ncbi.nlm.nih.gov)中進(jìn)行BLASTn搜索,確定是否為植原體基因序列;進(jìn)一步將16S rRNA基因片段序列通過(guò)植原體在線分類軟件iPhyClassifier(https:∥plantpathology.ba.ars.usda.gov/cgi-bin/resource/iphyclassifier.cgi)計(jì)算相似系數(shù)和虛擬RFLP(Virtual RFLP)分析,確定其植原體組和亞組[25]。利用在線分析工具M(jìn)USCLE(https:∥www.ebi.ac.uk/Tools/msa/muscle/),將所獲得的16S rRNA和SecY基因序列與已登錄GenBank的相關(guān)序列進(jìn)行一致性分析。采用MEGA 6.06的鄰接法(neighbor joining,NJ)[26]構(gòu)建基于16S rRNA和SecY基因的系統(tǒng)進(jìn)化樹,bootstrap值設(shè)置為1 000。
2 結(jié)果與分析
2.1 植原體PCR檢測(cè)結(jié)果
利用擴(kuò)增植原體16S rRNA的通用引物P1/P7、R16mF2/R16mR1對(duì)采集的7份花生叢枝病樣進(jìn)行PCR檢測(cè),結(jié)果兩對(duì)引物均能從7份病樣的總DNA中擴(kuò)增出與預(yù)期目的片段大小一致的條帶,陰性對(duì)照中未擴(kuò)增出任何片段(圖2a、b)。進(jìn)一步利用擴(kuò)增SecY基因的通用引物L(fēng)15F1/MapR1和SecYF2(Ⅱ)/SecYR1(Ⅱ)對(duì)7份花生叢枝病樣進(jìn)行巢式PCR擴(kuò)增,5份病樣總DNA中擴(kuò)增出與預(yù)期目的片段大小一致的特異性條帶,陰性對(duì)照中未擴(kuò)增出任何片段(圖2c)。這些結(jié)果表明,采集于廣東省湛江市遂溪縣的花生叢枝病樣中存在植原體。將該株系命名為廣東花生叢枝植原體(PnWB-GDSX-2020)。
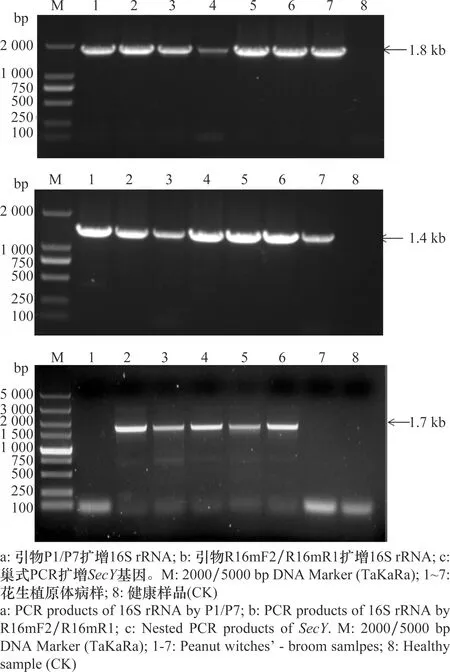
圖2 花生叢枝病樣中植原體16S rRNA、SecY基因PCR檢測(cè)結(jié)果Fig.2 PCR detection result of 16S rRNA and SecY genes of phytoplasma in peanut witches’-broom samples
2.2 16S rRNA基因序列分析
隨機(jī)挑取1個(gè)病樣的PCR產(chǎn)物進(jìn)行克隆、測(cè)序、分析,結(jié)果表明,從廣東遂溪花生病樣中獲得的16S rRNA片段大小為1 430 bp (GenBank登錄號(hào)為MZ427281),BLASTn結(jié)果顯示,與該片段有較高一致性的序列均為16SrⅡ組 (Peanut WB group)成員的16S rRNA序列。進(jìn)一步利用在線分析工具M(jìn)USCLE(https:∥www.ebi.ac.uk/Tools/msa/muscle/)對(duì)廣東花生叢枝植原體(PnWB-GDSX-2020)的16S rRNA與GenBank數(shù)據(jù)庫(kù)(https:∥www.ncbi.nlm.nih.gov/genbank/)相關(guān)序列進(jìn)行了核苷酸一致性分析(表1),結(jié)果顯示,PnWB-GDSX-2020與已報(bào)道的花生叢枝植原體組(16SrⅡ組)的16S rRNA核苷酸序列一致性在97%以上,與來(lái)自中國(guó)廣東、海南、臺(tái)灣的16SrⅡ-A、16SrⅡ-D和16SrⅡ-V亞組的11個(gè)植原體的一致性為100%,與來(lái)自中國(guó)海南和云南花生叢枝植原體的一致性為99.93%;而與其他亞組植原體的一致性較低,其中與16SrⅠ和16SrⅣ組植原體的一致性分別為89.47%和90.81%。
2.3 16S rRNA基因序列的系統(tǒng)進(jìn)化分析
采用MEGA 6.06的鄰接法構(gòu)建了PnWB-GDSX-2020 16S rRNA與GenBank數(shù)據(jù)庫(kù)中36個(gè)植原體的系統(tǒng)發(fā)育樹,以柑橘黃龍病菌亞洲種廣東株系(CandidatusLiberibacter asiaticus isolate Guangdong-HP)(GenBank登錄號(hào): DQ432005)的16S rRNA序列為外組。結(jié)果(圖3)顯示,PnWB-GDSX-2020與16Sr Ⅱ組的植原體株系聚集在一個(gè)大分支,與16Sr Ⅱ-A、16Sr Ⅱ-D、16Sr Ⅱ-V和16Sr Ⅱ-X亞組植原體株系形成一個(gè)小的分支,親緣關(guān)系最近;而與16Sr Ⅰ和16Sr Ⅳ組的植原體親緣關(guān)系遠(yuǎn)。

表1 PnWB-GDSX-2020與其他植原體的16S rRNA基因序列一致性1)
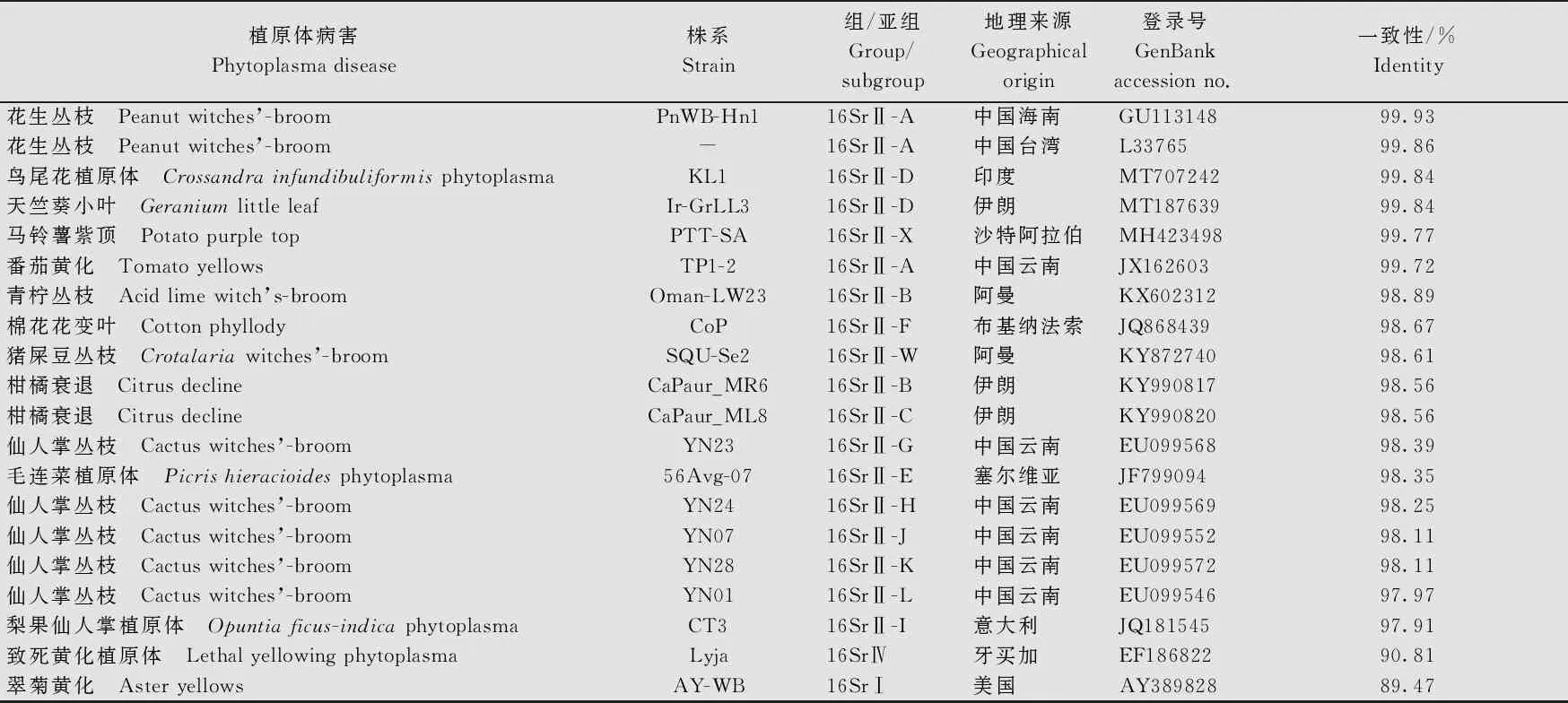
續(xù)表1 Table 1(Continued)

圖3 PnWB-GDSX-2020與其他36個(gè)植原體代表株系基于16S rRNA基因序列的系統(tǒng)進(jìn)化樹Fig.3 Phylogenetic tree constructed based on 16S rRNA gene sequences of PnWB-GDSX-2020 and 36 other phytoplasmas
2.4 16S rRNA基因序列的iPhyClassifier在線分析
將獲得的PnWB-GDSX-2020 16S rRNA基因序列利用植原體在線分類軟件iPhyClassifier進(jìn)行分析。結(jié)果顯示PnWB-GDSX-2020 16S rRNA序列與‘CandidatesPhytoplasma australasiae’ 菌株(GenBank登錄號(hào):U15442)的一致性最高,為98.5%,這說(shuō)明PnWB-GDSX-2020是‘CandidatusPhytoplasma australasiae’的相關(guān)菌株;PnWB-GDSX-2020 16S rRNA序列片段的17種限制性內(nèi)切酶虛擬RFLP 圖譜與16SrⅡ-V亞組的參照株系‘Praxelisclematidea’ phyllody phytoplasma (GenBank 登錄號(hào):KY568717) 酶切圖譜一致(相似系數(shù)1.00),表明PnWB-GDSX-2020屬于16SrⅡ-V亞組成員。
2.5 SecY基因序列分析
利用擴(kuò)增SecY基因的通用引物,從廣東遂溪花生病樣總DNA中PCR擴(kuò)增獲得該基因片段,其大小為1 709 bp (GenBank登錄號(hào)為MZ437794),其中該片段序列318-1580 nt為完整SecY基因序列,編碼420個(gè)氨基酸。BLASTn結(jié)果顯示,與該片段有較高一致性的序列均為16SrⅡ組成員的SecY基因序列。進(jìn)一步利用在線分析工具M(jìn)USCLE對(duì)PnWB-GDSX-2020的SecY基因與GenBank數(shù)據(jù)庫(kù)(https:∥www.ncbi.nlm.nih.gov/genbank/)相關(guān)序列進(jìn)行核苷酸一致性分析,結(jié)果(表2)表明,PnWB-GDSX-2020與已報(bào)道的16SrⅡ組的SecY基因核苷酸序列一致性在98%以上,其中與來(lái)自中國(guó)云南、中國(guó)臺(tái)灣、印度的15個(gè)植原體的SecY基因核苷酸一致性為98.83%~99.82%;而與其他亞組植原體的SecY基因核苷酸序列一致性較低,其中與16SrⅠ和16SrⅤ組植原體的一致性分別為64.05%和69.74%。進(jìn)一步采用MEGA 6.06的鄰接法,構(gòu)建了PnWB-GDSX-2020SecY與GenBank數(shù)據(jù)庫(kù)中17個(gè)植原體的系統(tǒng)發(fā)育樹,結(jié)果(圖4)顯示,PnWB-GDSX-2020與16SrⅡ組的植原體株系聚集在一個(gè)大分支,親緣關(guān)系最近;與16SrⅠ和16SrⅤ組的植原體親緣關(guān)系遠(yuǎn)。

表2 PnWB-GDSX-2020與其他植原體的Sec Y基因序列一致性1)

圖4 PnWB-GDSX-2020與其他17個(gè)植原體代表株系基于SecY 基因序列的系統(tǒng)進(jìn)化樹Fig.4 Phylogenetic tree constructed based on SecY gene sequences of PnWB-GDSX-2020 and 17 other phytoplasmas
3 結(jié)論與討論
本研究通過(guò)PCR擴(kuò)增、克隆、測(cè)序獲得廣東花生叢枝病相關(guān)植原體(PnWB-GDSX-2020)的16S rRNA和SecY基因序列,進(jìn)一步通過(guò)對(duì)16S rRNA基因序列的iPhyClassifier在線分析、序列一致性和系統(tǒng)進(jìn)化分析,明確廣東花生叢枝病相關(guān)植原體為16SrⅡ-V亞組植原體。同時(shí),SecY基因序列分析結(jié)果也顯示,PnWB-GDSX-2020與花生叢枝植原體組的一致性最高,親緣關(guān)系最近。廣東花生叢枝病于20世紀(jì)50年代就有發(fā)生,至今僅見(jiàn)通過(guò)電子顯微鏡觀察、介體和嫁接傳病來(lái)鑒定其病原的報(bào)道,本文從分子水平明確了廣東花生叢枝病相關(guān)植原體,并確定了其分類地位,這些結(jié)果為開展植原體16S rRNA和SecY基因序列變異與進(jìn)化等研究提供了基礎(chǔ)數(shù)據(jù)。
植原體是一類不能人工培養(yǎng)的植物病原細(xì)菌,分類鑒定主要依據(jù)16S rRNA、rp、SecY等保守基因序列分析。本文利用擴(kuò)增16S rRNA基因的兩對(duì)通用引物從7份花生病樣中均擴(kuò)增出條帶,而經(jīng)過(guò)多次試驗(yàn)通過(guò)SecY基因的通用引物進(jìn)行巢式PCR僅能從5份病樣中擴(kuò)增出目的條帶,其他2份病樣未能擴(kuò)增出條帶。16S rRNA基因是植原體高度保守基因,也是目前植原體分類的主要分子依據(jù),而rp、SecY等其他基因遺傳變異較大。本文擴(kuò)增SecY基因序列采用的引物位于rpl15和map基因之間,擴(kuò)增的目的片段包含部分rpl15基因、完整SecY基因以及部分map基因,推測(cè)2份樣品未能擴(kuò)增出目的條帶是由于引物結(jié)合位點(diǎn)的植原體基因序列發(fā)生變異導(dǎo)致。
植原體病害種類多,寄主范圍廣,已在農(nóng)林領(lǐng)域造成嚴(yán)重危害。病原種類鑒定是指導(dǎo)相關(guān)病害防控的前提。而用于植原體檢測(cè)與鑒定的技術(shù)主要有組織學(xué)觀察、生物學(xué)方法、血清學(xué)檢測(cè)、分子檢測(cè)技術(shù)。廣東省常年高溫高濕,植物和介體昆蟲都可以周年生長(zhǎng)與繁殖,植原體等各類植物病害發(fā)生嚴(yán)重。不少學(xué)者采用不同鑒定技術(shù)對(duì)廣東相關(guān)植原體病害進(jìn)行鑒定。如20世紀(jì)80年代通過(guò)組織學(xué)觀察和生物學(xué)方法明確廣東地區(qū)的花生叢枝病、芝麻花變?nèi)~病是由植原體引起的[14,27];1994年,同樣采用組織學(xué)觀察和生物學(xué)方法明確水稻橙葉病是一種植原體引致的新病害[28]。隨著分子生物學(xué)技術(shù)的發(fā)展,2019年通過(guò)分子檢測(cè)技術(shù)明確廣東棗瘋病植原體屬于16SrⅤ-B亞組[29],廣東茄子花變?nèi)~病植原體屬于16SrⅡ-D亞組[30]。本文通過(guò)分子生物學(xué)技術(shù)確定了廣東花生叢枝病植原體屬于16SrⅡ-V亞組,且與先前報(bào)道的茄子花變?nèi)~、豆角花變?nèi)~相關(guān)植原體16S rRNA基因序列(GenBank登錄號(hào):MH667642、MW426146)一致性100%,但茄子花變?nèi)~植原體病害發(fā)生在粵東惠州、豆角花變?nèi)~植原體病害發(fā)生在粵西湛江,由此初步推測(cè),該植原體病害極有可能通過(guò)介體昆蟲的傳播在全省蔓延,這一研究結(jié)果為廣東省植原體病害的防控提供了科學(xué)依據(jù)。
猜你喜歡
遼寧教育(2022年19期)2022-11-18 07:20:42
公民與法治(2022年5期)2022-07-29 00:47:28
汽車實(shí)用技術(shù)(2022年9期)2022-05-20 05:51:26
教學(xué)考試(高考物理)(2021年5期)2021-11-08 10:31:22
歷史教學(xué)問(wèn)題(2021年4期)2021-11-05 07:02:34
中醫(yī)眼耳鼻喉雜志(2021年1期)2021-07-22 07:38:14
裝備制造技術(shù)(2020年11期)2021-01-26 00:39:12
中國(guó)公共安全(2017年11期)2017-02-06 05:28:08
電測(cè)與儀表(2016年7期)2016-04-12 00:22:18
燕山大學(xué)學(xué)報(bào)(2015年4期)2015-12-25 02:19:49
