華北亞高山土壤細(xì)菌群落及氮循環(huán)對退耕還草的響應(yīng)
2022-10-13 05:14:46王禮霄劉晉仙柴寶峰
生態(tài)環(huán)境學(xué)報(bào) 2022年8期
王禮霄,劉晉仙,柴寶峰
山西大學(xué)黃土高原研究所/黃土高原生態(tài)恢復(fù)山西省重點(diǎn)實(shí)驗(yàn)室,山西 太原 030006
亞高山草地具有水源涵養(yǎng)、生物多樣性保育、水土保持和碳匯功能(趙鵬宇,2019)。受全球變化和人類活動的影響,山西亞高山草地植被退化,水土流失,生態(tài)系統(tǒng)功能嚴(yán)重受損。基于生態(tài)環(huán)境的保護(hù)和改善,我國實(shí)施了“退耕還林/草”工程(常慶瑞等,1999)。山西亞高山草甸面積約33.33×104hm2,占全省草地總面積的10%。這一重要的政策實(shí)施以來,植被得以迅速恢復(fù)。植物是生態(tài)系統(tǒng)結(jié)構(gòu)和功能自然演化的主要驅(qū)動力(史利江等,2021),凋落物和根系分泌物的輸入影響土壤的理化性質(zhì)(Kuypers et al.,2018),進(jìn)而驅(qū)動了土壤微生物群落的演替過程(Kardol et al.,2010;Bardgett et al.,2014)。亞高山生態(tài)系統(tǒng)對氣候變化敏感,恢復(fù)較為困難,植被和土壤微生物群落的演替規(guī)律尚不清楚。
目前關(guān)于退耕還草地恢復(fù)過程中植物多樣性和土壤特征的變化規(guī)律已有了一定的研究。隨著演替的進(jìn)行,植物生物量和物種多樣性會隨著時間的推移而增加(樊博等,2020)。自然植被的恢復(fù)可以改善退化土壤的性質(zhì)并增加肥力(Cao et al.,2021)。植物演替對土壤養(yǎng)分有積極的作用,并增強(qiáng)了植物的多樣性(Deng et al.,2014;Matetskaya et al.,2021)。Wang et al.(2011)發(fā)現(xiàn)黃土高原農(nóng)田棄耕后,土壤有機(jī)碳、全氮、速效氮和速效鉀以及土壤酶活性隨著植被恢復(fù)年限的增加而提高。
土壤微生物在生態(tài)系統(tǒng)物質(zhì)循環(huán)過程中發(fā)揮著關(guān)鍵作用,微生物群落結(jié)構(gòu)和功能的變化會影響植物殘?bào)w分解、土壤碳氮轉(zhuǎn)變和地上植被養(yǎng)分供給等生態(tài)過程(Wang et al.,2011)。同時,微生物多樣性、結(jié)構(gòu)和功能也受到植被、土壤特性等因素的影響(Fan et al.,2016;Bier et al.,2014)。有越來越多的研究關(guān)注于土壤微生物群落對地上植被演替的響應(yīng),地上植被的演替過程會對土壤微生物類群和功能多樣性等造成重要影響(Lee-Cruz et al.,2013;Nacke et al.,2014)。Li et al.(2014)和Zuo et al.(2016)對植被恢復(fù)礦區(qū)和沙地草原土壤微生物群落的分析發(fā)現(xiàn),細(xì)菌的多樣性隨著植被恢復(fù)時間的增長而顯著提高,呈現(xiàn)出明顯的時間演替趨勢。多樣性的增加對微生物群落的功能產(chǎn)生重要的影響,功能基因豐度可以反映微生物群落的功能對環(huán)境變化的響應(yīng)(Li et al.,2014)。目前關(guān)于地上植被如何影響土壤微生物氮循環(huán)功能仍有爭議,Blaud et al.(2017)報(bào)道,在從農(nóng)田到森林的次生演替過程中,氮循環(huán)相關(guān)基因的豐度增加;而Zhong et al.(2018)觀察到,在從草原到森林的次生演替過程中,氮循環(huán)相關(guān)基因的相對豐度則是先升高后降低。因此,退耕還草過程中植物群落與微生物群落結(jié)構(gòu)、多樣性和功能之間的關(guān)系是恢復(fù)生態(tài)學(xué)有待回答的問題。
本研究以山西省寧武云中山亞高山退耕還草區(qū)不同恢復(fù)年限(15、20、30 a)的草地為研究對象,進(jìn)行草地植物群落調(diào)查和土壤樣本的采集,利用16S rRNA基因的Illumina MiSeq測序和qPCR技術(shù),通過多元統(tǒng)計(jì)的方法分析草甸恢復(fù)過程中土壤細(xì)菌群落的結(jié)構(gòu)與功能特征及其驅(qū)動因素,闡明土壤細(xì)菌群落對地上植被恢復(fù)時間序列的響應(yīng)。本研究旨在回答以下問題:(1)亞高山草草甸退耕還草過程中,植物多樣性、土壤性質(zhì)和土壤細(xì)菌群落的演變規(guī)律;(2)退耕還草過程中土壤細(xì)菌群落結(jié)構(gòu)和氮循環(huán)功能演變的驅(qū)動機(jī)制。
1 材料與方法
1.1 樣品采集
研究地點(diǎn)位于山西省呂梁山脈北段分支寧武云中山,呈東北—西南走向,海拔約 2160 m。本區(qū)年平均氣溫2.0—9.0 ℃,無霜凍期84—135 d,年降水量430—700 mm。該地區(qū)自上世紀(jì)90年代開始實(shí)施退耕還草措施,農(nóng)田植被開始自然恢復(fù)。1個農(nóng)田樣地(對照)和3個不同恢復(fù)年限的草甸(15、20、30 a)作為試驗(yàn)樣地。樣地具有相似的坡度和海拔,并經(jīng)歷了類似的耕作方式。退耕前,農(nóng)田的主要作物是莜麥,退耕后,植被自然演替。選擇對照農(nóng)田種植的也是莜麥,在春季施用了農(nóng)家肥。
在每個樣地,設(shè)置3個1 m×1 m的樣方,樣方之間間隔約20 m。用土鉆采集表層土壤樣品(0—10 cm),每個樣方內(nèi)采集5個子樣(每個角1個,中心1個),在聚乙烯袋中混合成一個樣品。經(jīng)2 mm網(wǎng)篩,將大部分根、動物和石頭去除,然后將樣本分成兩部分。其中一部分風(fēng)干后進(jìn)行土壤理化性質(zhì)測定,另一部分保存在-80 ℃冰箱中,用于DNA的提取。在每個樣方中調(diào)查植被參數(shù),記錄每種植物的名稱、高度、蓋度及多度等指標(biāo)。
1.2 土壤理化性質(zhì)測定
烘干法測定土壤含水量(SWC);土壤pH值用電位法(HANNA,意大利)測定(土水質(zhì)量比為1∶2.5);總碳(TC)和總氮(TN)通過元素分析儀(ElementarVario MACRO,德國)測定;銨態(tài)氮(NH4+-N)、硝態(tài)氮(NO3--N)采用間斷元素分析儀(CleverChem 380,德國)測定。
脲酶采用苯酚鈉-次氯酸鈉比色法,其活性以37 ℃下培養(yǎng)24 h后1 g土壤產(chǎn)生的NH3的mg數(shù)表示;蔗糖酶采用3,5-二硝基水楊酸比色法,其活性以37 ℃培養(yǎng)24 h后1 g土壤產(chǎn)生的葡萄糖的mg數(shù)表示;過氧化氫酶活性(H2O2)采用KMnO4滴定法,其活性以20 min內(nèi)每克土壤分解的過氧化氫的毫克數(shù)來表示。
1.3 DNA提取、高通量測序及生物信息學(xué)分析
稱取0.5 g土壤樣品,使用E.Z.N.A.土壤DNA試劑盒(Omega Bio-tek,USA)提取和純化土壤微生物DNA。將每個采樣點(diǎn)的3個土壤樣品等體積混合,一共12個DNA樣品,送往上海美吉生物醫(yī)藥科技有限公司進(jìn)行高通量測序。采用338F(正向引物 5′-ACTCCTACGGGAGGCAGCA-3′)和806R(反向引物 5′-GGACTACHVGGGTWTCTAAT-3′)對細(xì)菌16S rRNA的V3—V4高可變區(qū)進(jìn)行PCR擴(kuò)增,并通過Illumina Miseq測序平臺對擴(kuò)增產(chǎn)物進(jìn)行測序。剔除嵌合序列后,剩下的序列使用UPARSE以97%的相似性作為閾值劃分分類操作單元(operational taxonomic units,OTUs)。
1.4 氮循環(huán)功能基因的定量PCR
采用CFX 96 touch實(shí)時定量 PCR系統(tǒng)(Bio-Rad,Hercules,CA,USA)測定參與土壤氮循環(huán)過程的主要功能基因(nifH、amoA-AOA、amoA-AOB、nirK、nirS)豐度。操作過程和引物序列見課題組之前的報(bào)道(Luo et al.,2020)。反應(yīng)體系、擴(kuò)增程序及熔解曲線見文獻(xiàn)(徐白璐等,2017)。以含有目的基因片段的重組質(zhì)粒為模板,10倍倍比進(jìn)行稀釋,根據(jù)情況選取4—5個稀釋梯度制作標(biāo)準(zhǔn)曲線。將樣品與標(biāo)準(zhǔn)品同時進(jìn)行 qPCR檢測,每個樣品設(shè)置 3個重復(fù)和一個陰性對照。qPCR的擴(kuò)增效率是 80%—95%,標(biāo)準(zhǔn)曲線的R2>0.990。所有功能基因的豐度最終計(jì)算為每克干土的拷貝數(shù)。
1.5 數(shù)據(jù)分析
基于R studio(v.3.4.3)vegan包,計(jì)算細(xì)菌和植物的α多樣性和β多樣性,采用非度量多維尺度分析(Non-metric multidimensional scaling,NMDS)和相似性分析(Analysis of similarity,ANOSIM)對農(nóng)田與不同恢復(fù)年限草地細(xì)菌群落結(jié)構(gòu)進(jìn)行比較。采用冗余分析(Redundancy analysis,RDA)評價細(xì)菌群落結(jié)構(gòu)與環(huán)境變量之間的相關(guān)性。利用IBM SPSS statistics 20進(jìn)行皮爾遜相關(guān)性分析(Pearson correlation test)以及采用單因素方差分析(One-way analysis of variance)和Duncan多重比較分析進(jìn)行顯著性差異分析。所有統(tǒng)計(jì)分析的顯著性水平均為P<0.05。
2 結(jié)果
2.1 土壤理化性質(zhì)和植物多樣性特征
不同恢復(fù)年限草地的土壤理化參數(shù)和植物多樣性數(shù)據(jù)見表1。所有土壤樣品的pH都小于7,為弱酸性土壤。土壤總碳、總氮、碳氮比、硝態(tài)氮、銨態(tài)氮、土壤含水率、過氧化氫酶、脲酶和蔗糖酶活性在農(nóng)田和不同年限恢復(fù)年限的草地間差異顯著(P<0.05),而土壤pH的差異不顯著(P>0.05)。農(nóng)田土壤總碳、總氮、銨態(tài)氮、硝態(tài)氮含量、脲酶活性顯著高于草地,而在草地恢復(fù)過程中土壤總碳和總氮含量穩(wěn)步提升;退耕30年草地土壤碳氮比、含水率和蔗糖酶活性顯著高于其他樣地。不同恢復(fù)年限的草地地上植被多樣性指數(shù)(Shannon index)和豐富度指數(shù)(Richness index)差異顯著(P<0.05),但均勻度差異不顯著(P>0.05)。恢復(fù)15年的草地植物生物多樣性最高,隨著恢復(fù)年限的加長,草地優(yōu)勢種發(fā)生變化,生物多樣性降低。
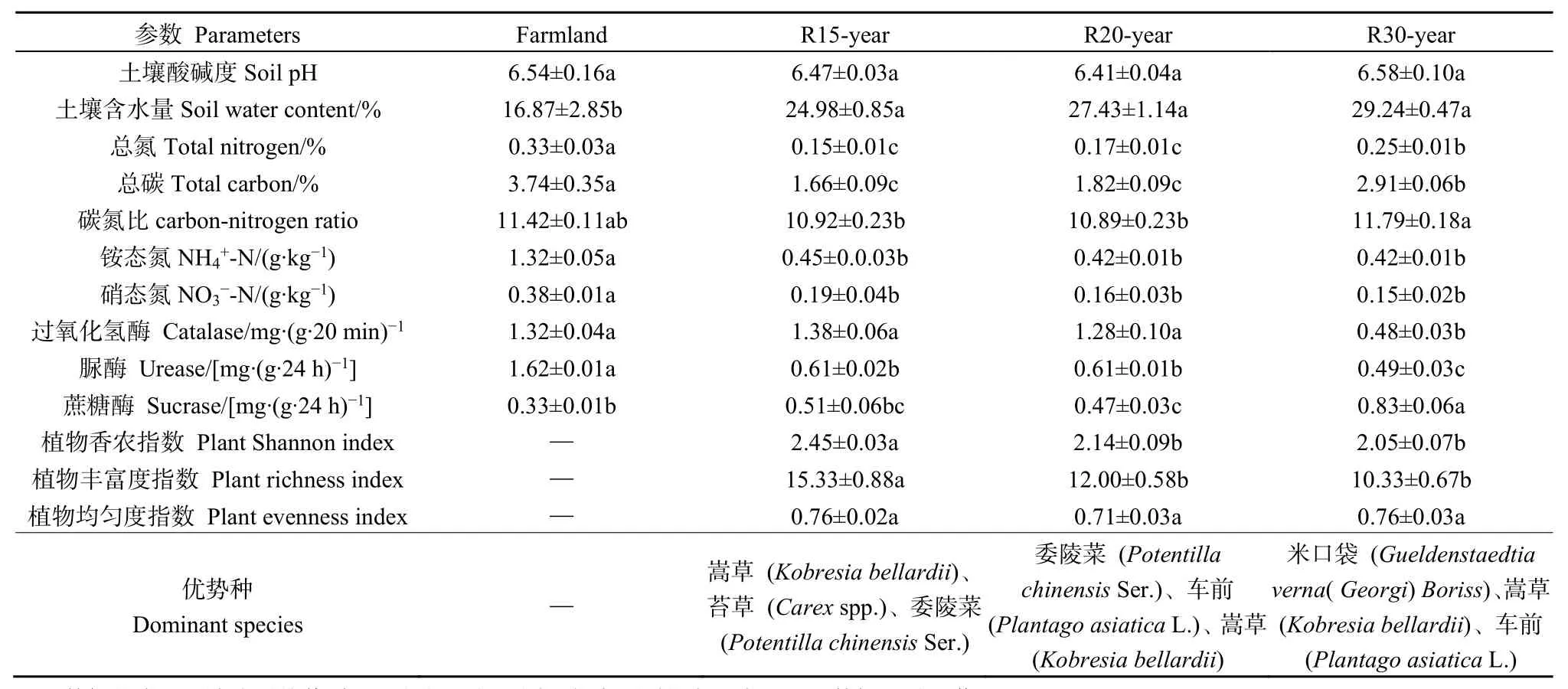
表1 農(nóng)田與不同恢復(fù)年限草地土壤理化性質(zhì)與植物群落參數(shù)Table 1 Soil physicochemical properties and plant community parameters under different revegetation habitats and farmland
2.2 細(xì)菌群落組成和多樣性
通過高通量測序,在12個樣本中共檢測到358488個細(xì)菌序列。按最小樣本序列數(shù)抽平,共鑒定出5410個OTU(>97%序列相似性水平),屬于36個門、118個綱、295個目、449個科、808個屬和1709個種。36個門中有11個門定義為優(yōu)勢細(xì)菌門(樣本中相對豐度>1%),相對豐度從高到低,依次為放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、酸桿菌門(Acidobacteria)、綠彎菌門(Chloroflexi)、芽單胞菌門(Gemmatimonadetes)、厚壁菌門(Firmicutes)、疣微菌門(Verrucomicrobia)、擬桿菌門(Bacteroidetes)、髕骨細(xì)菌門(Patescibacteria)、甲基肌酐門(Methylomirabilota)和粘球菌門(Myxococcota)(圖1a)。在不同恢復(fù)年限退耕草地的土壤中,這些優(yōu)勢細(xì)菌門占據(jù)了95%以上的細(xì)菌序列。綠彎菌門的相對豐度在不同恢復(fù)年限退耕草地之間存在顯著差異(P<0.05)(圖1b)。
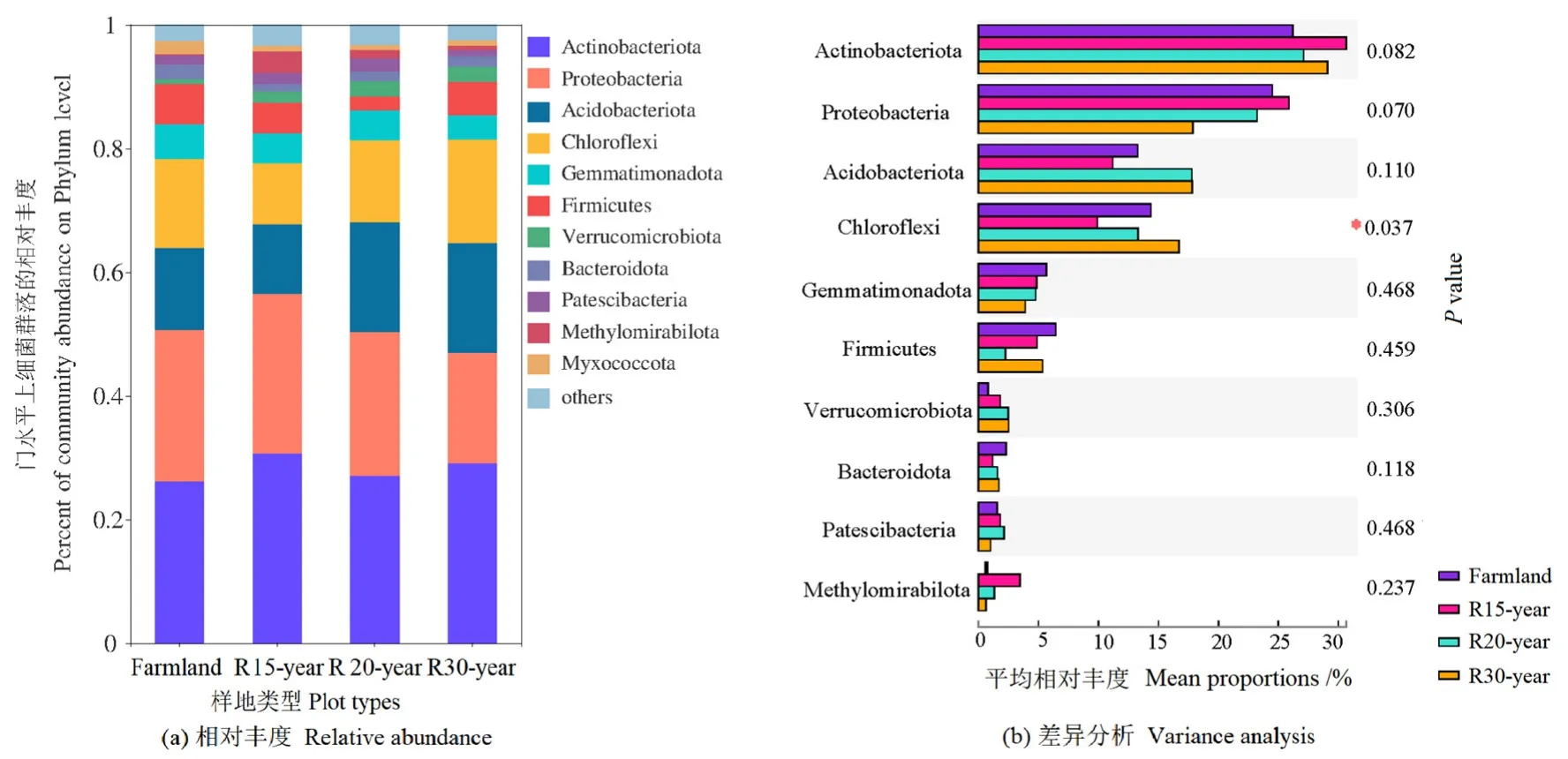
圖1 農(nóng)田和不同退耕年限草地細(xì)菌群落在門水平的相對豐度及差異分析Figure 1 The relative abundance of dominant bacteria phylum and variation analysis from different revegetation habitats and farmland
12個相對豐度最高的細(xì)菌綱、科和屬,在亞高山不同恢復(fù)年限退耕草地間存在差異(圖2)。α變形菌綱(Alphaproteobacteria)、γ變形菌綱(Gammaproteobacteria)、Blastocatellia、綠彎菌綱(Chloroflexia)的豐度差異顯著;在科水平,黃色桿菌科(Xanthobacteraceae)、Pyrinomonadaceae、Roseiflexaceae的差異顯著;RB41和norank_f_Roseiflexaceae屬在不同恢復(fù)年限退耕草地間存在顯著差異(P<0.05)。
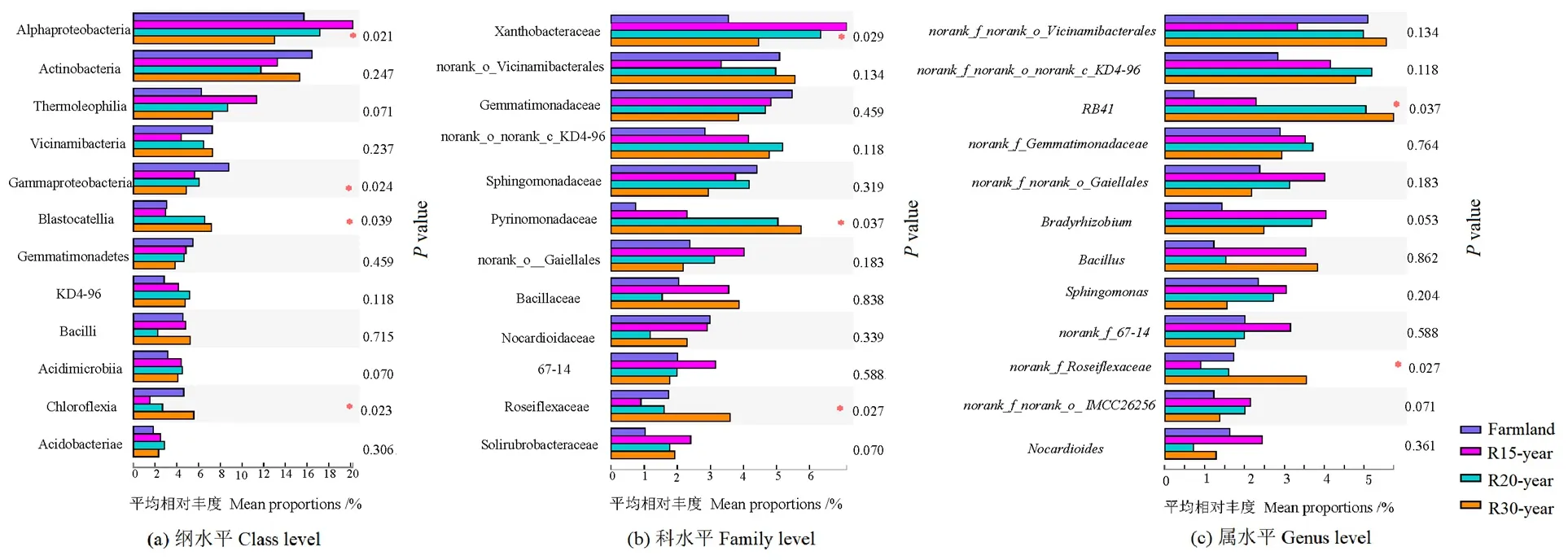
圖2 農(nóng)田和不同退耕年限草地細(xì)菌群落在綱、科、屬水平的差異分析Figure 2 The variation analysis of dominant bacteria class,family and genus from different revegetation habitats and farmland
不同恢復(fù)年限退耕草地的土壤細(xì)菌群落α多樣性分析結(jié)果見表2。其中農(nóng)田土壤細(xì)菌群落Shannon指數(shù)顯著高于退耕還草地,Simpson指數(shù)顯著低于退耕還草地,即農(nóng)田土壤細(xì)菌群落多樣性最高,而均勻度較低。自然恢復(fù) 15年的退耕地的細(xì)菌群落Shannon指數(shù)顯著低于其他樣地,Simpson指數(shù)顯著高于其他樣地。隨著自然恢復(fù)年限的增長,細(xì)菌群落Shannon指數(shù)增加,Simpson指數(shù)降低。農(nóng)田土壤細(xì)菌群落Sobs指數(shù)、ACE指數(shù)和Chao1指數(shù)最大,其土壤細(xì)菌群落種類數(shù)目最多,在退耕地自然恢復(fù)過程中,細(xì)菌群落種類數(shù)目呈增加趨勢。
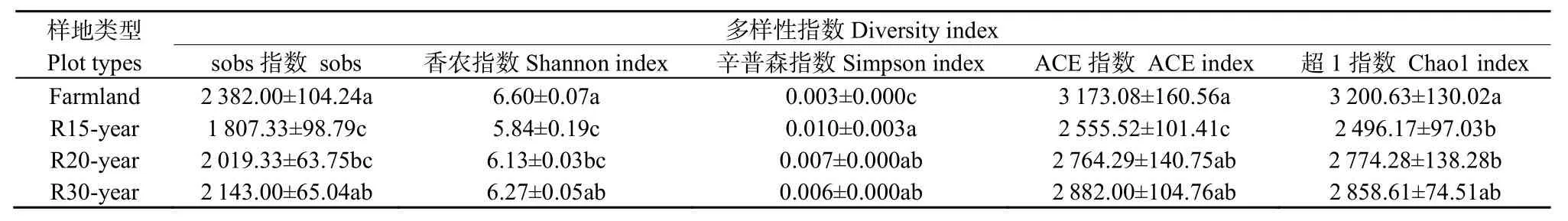
表2 農(nóng)田和不同恢復(fù)年限草地土壤細(xì)菌群落多樣性Table 2 Diversity of bacteria communities of different revegetation habitats and farmland
基于Bray-Curtis距離的非度量多維尺度分析(NMDS)和ANOSIM結(jié)果表明,不同恢復(fù)年限退耕草地的土壤細(xì)菌群落結(jié)構(gòu)發(fā)生了顯著的變化(R2=0.5014,P<0.01)(圖3a)。統(tǒng)計(jì)分析結(jié)果顯示,細(xì)菌群落的Bray-Curtis距離符合正態(tài)分布(P>0.05),土壤細(xì)菌群落 β多樣性(Bray-Curtis距離)與組間時間距離呈顯著正相關(guān)(r=0.449,P<0.001),土壤細(xì)菌群落之間的相異性隨著恢復(fù)時間的增加而增加(圖3b)。
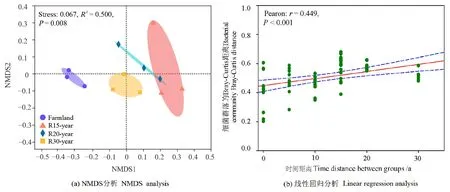
圖3 農(nóng)田和不同恢復(fù)年限草地細(xì)菌群落基于Bray-Curtis距離的NMDS分析及其與線性回歸分析Figure 3 NMDS and linear regression analysis of bacteria communities based on Bray-Curtis distance among different revegetation habitats and farmland
2.3 環(huán)境因子對細(xì)菌群落的影響
環(huán)境因子對不同恢復(fù)年限草地細(xì)菌群落的組成具有一定的影響。冗余分析(RDA)的前兩個軸解釋了總變量的48.42%。從圖4a可知,環(huán)境變量總體上沿著時間序列分布。在環(huán)境變量中,總碳、總氮、碳氮比與銨態(tài)氮含量是細(xì)菌群落結(jié)構(gòu)變化的重要驅(qū)動因子(P<0.05)。此外,通過環(huán)境因子對細(xì)菌主要類群影響的Pearson相關(guān)分析發(fā)現(xiàn)(圖4b),土壤總碳、總氮與綠彎菌門、粘球菌門顯著正相關(guān)(P<0.01),土壤含水量與變形菌門和蛭弧菌門(Bdellovibrionota)顯著負(fù)相關(guān)(P<0.01),與浮霉菌門顯著正相關(guān)(P<0.01),土壤銨態(tài)氮含量與黏細(xì)菌門顯著正相關(guān)(P<0.01),土壤硝態(tài)氮含量與變形菌門、厚壁菌門、擬桿菌門與粘球菌門顯著正相關(guān)(P<0.01),與浮霉菌門與甲基肌酐門顯著負(fù)相關(guān)。植物群落多樣性與土壤細(xì)菌群落之間的相關(guān)性分析結(jié)果表明(圖 5),土壤細(xì)菌和植物的Shannon指數(shù)顯著相關(guān)(P<0.05),而它們之間的β多樣性沒有顯著的相關(guān)關(guān)系(r=0.127,P>0.05)。
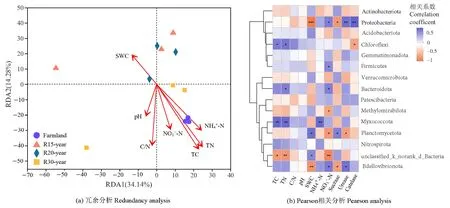
圖4 農(nóng)田和不同恢復(fù)年限草地細(xì)菌群落組成的冗余分析及優(yōu)勢門與環(huán)境因子的Pearson相關(guān)分析Figure 4 Redundancy analysis of bacterial community composition and Pearson analysis with dominant phylum in different revegetation grasslands and farmland
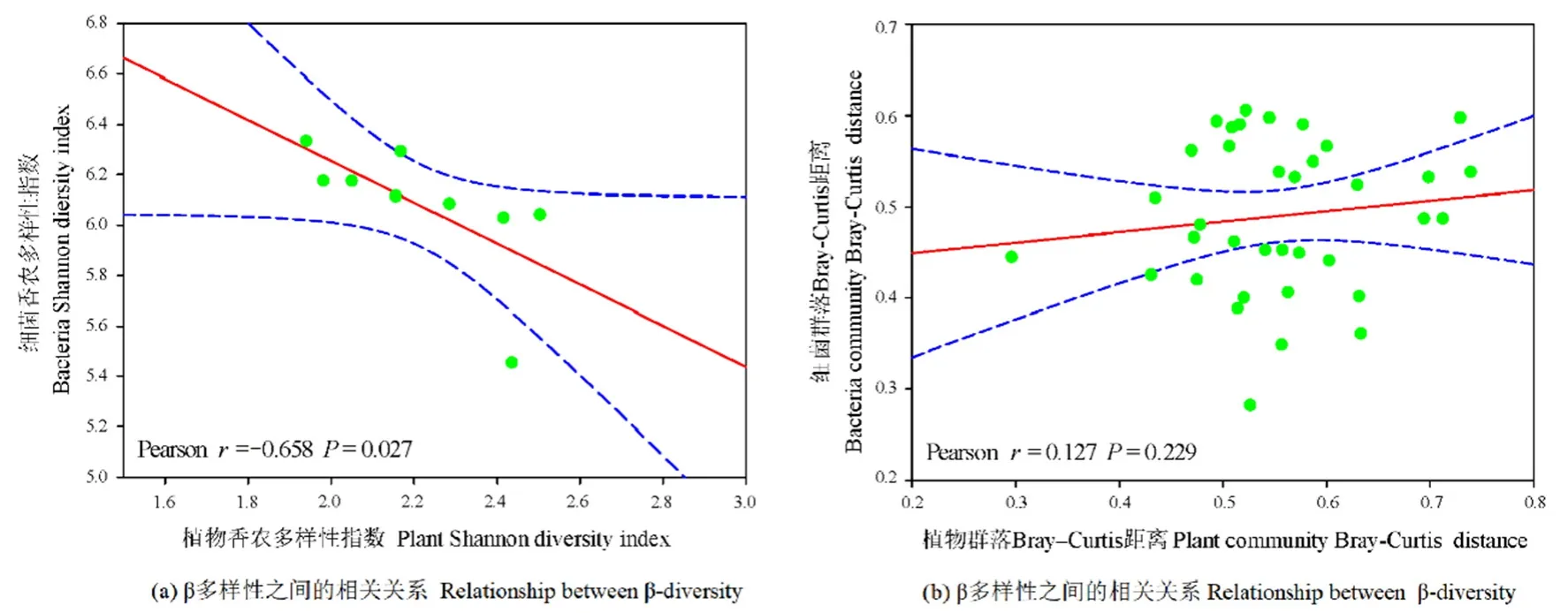
圖5 農(nóng)田和不同恢復(fù)年限草地土壤細(xì)菌群落與植物群落α多樣性和β多樣性之間的相關(guān)關(guān)系(基于Bray-Curtis距離)Figure 5 Relationship between plant and soil bacterial α-diversity and β-diversity (based on Bray-Curtis distances)in different revegetation grasslands and farmland
2.4 退耕還草過程中氮循環(huán)功能基因的變化特征
利用實(shí)時定量 PCR方法分析了土壤中與氮循環(huán)有關(guān)的關(guān)鍵功能基因,包括固氮有關(guān)的nifH基因、與硝化作用有關(guān)的amoA-AOA基因、amoA-AOB基因、反硝化作用有關(guān)的nirK和nirS基因。結(jié)果顯示,在4個樣地中土壤中nifH的拷貝數(shù)是 1.31×108—4.04×108g-1,amoA-AOA 的拷貝數(shù)是 0.33×108—2.47×108g-1,amoA-AOB 的拷貝數(shù)是0.31×107—7.28×107g-1,nirK 的拷貝數(shù)是 3.32×107—4.55×107g-1,nirS的拷貝數(shù)是0.28×107—4.30×107g-1(圖6)。農(nóng)田中5種氮循環(huán)有關(guān)的關(guān)鍵功能基因的拷貝數(shù)均顯著高于退耕還草自然恢復(fù)樣地,amoA-AOA、amoA-AOB、nifH、nirS的拷貝數(shù)在4個樣地間有顯著差異(P<0.05),且隨著恢復(fù)年限的增長,amoA-AOB與 nifH的拷貝數(shù)顯著增加(P<0.05),nirK的拷貝數(shù)在4個樣地間無顯著差異(P>0.05)。
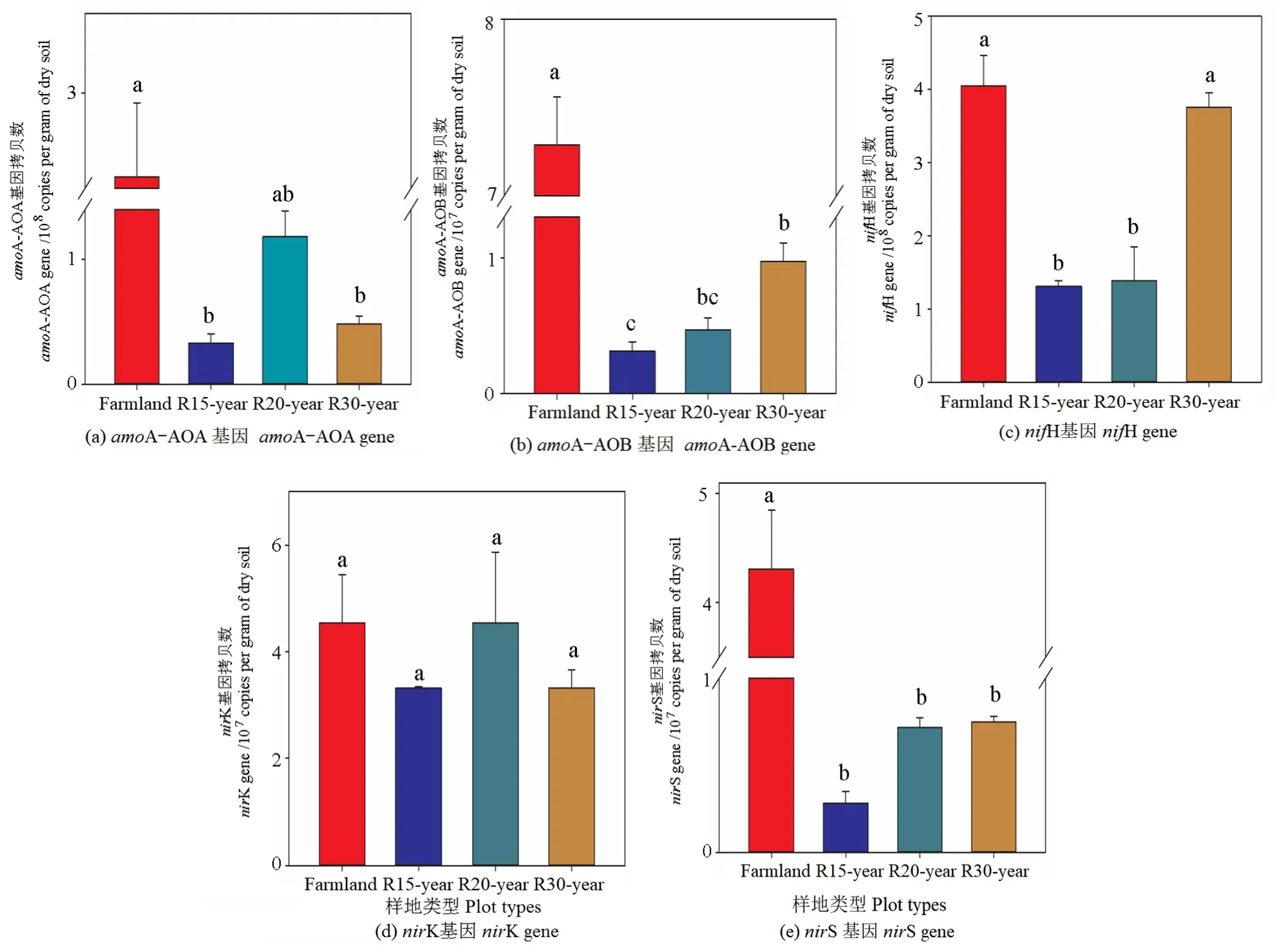
圖6 農(nóng)田和不同恢復(fù)年限草地參與氮循環(huán)主要過程的功能基因拷貝數(shù)(amoA-AOA,amoA-AOB,nifH,nirK and nirS)Figure 6 Gene copy numbers of functional genes involved in major steps of the nitrogen cycle (amoA-AOA,amoA-AOB,nifH,nirK and nirS) in different revegetation grasslands and farmland
2.5 氮循環(huán)功能基因與環(huán)境參數(shù)和優(yōu)勢類群的相關(guān)關(guān)系
用Pearson相關(guān)性分析環(huán)境因子對氮循環(huán)相關(guān)基因拷貝數(shù)的影響。編碼氨單加氧酶基因amoA-AOA與amoA-AOB對環(huán)境因子的響應(yīng)一致,amoA-AOA與amoA-AOB與總碳、總氮、銨態(tài)氮、硝態(tài)氮顯著正相關(guān),與土壤含水率顯著負(fù)相關(guān)(圖7)。編碼固氮酶基因 nifH、總碳、總氮、碳氮比與銨態(tài)氮顯著正相關(guān)。編碼亞硝酸還原酶基因nirS、nirK對環(huán)境因子的響應(yīng)不同,環(huán)境因子與nirK相關(guān)性不顯著,但nirS與總碳、總氮、銨態(tài)氮與硝態(tài)氮顯著正相關(guān),與土壤含水量顯著負(fù)相關(guān)。
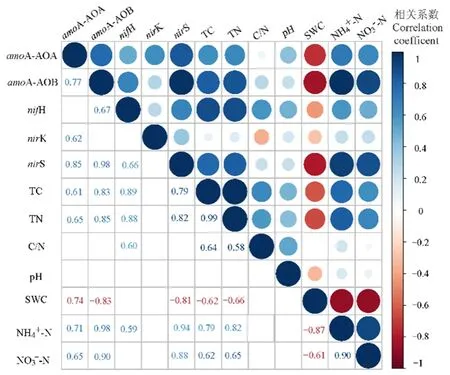
圖7 參與氮循環(huán)主要過程的功能基因與土壤理化性質(zhì)的相關(guān)關(guān)系Figure 7 Correlations between gene copy numbers of functional genes involving in nitrogen cycling and soil physicochemical properties
氮循環(huán)相關(guān)基因與細(xì)菌優(yōu)勢門的相關(guān)性(表3)表明,氮循環(huán)相關(guān)基因與優(yōu)勢門具有一定的相關(guān)性。其中,amoA-AOA基因與綠彎菌門和擬桿菌門極顯著正相關(guān)(P<0.01),與粘球菌門顯著正相關(guān)(P<0.05);amoB-AOB基因與綠彎菌門、擬桿菌門和粘球菌門極顯著正相關(guān)(P<0.01);nirK基因與擬桿菌門極顯著正相關(guān)(P<0.01);nirS基因與綠彎菌門、擬桿菌門和粘球菌門極顯著正相關(guān)(P<0.01)。
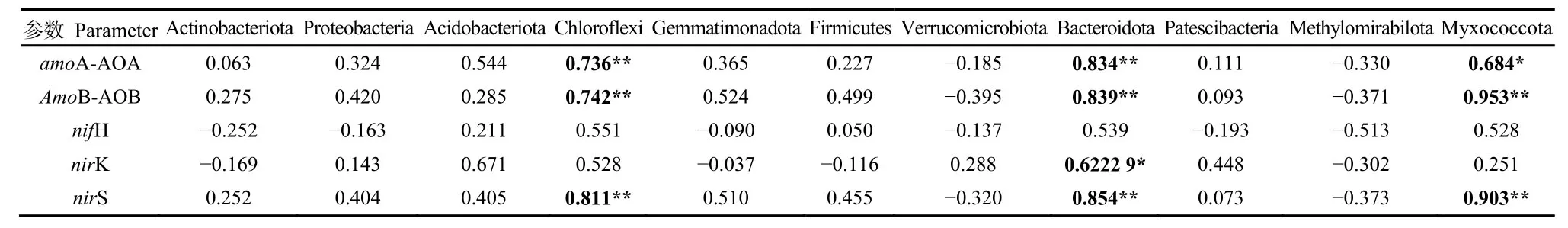
表3 參與氮循環(huán)主要過程的功能基因與優(yōu)勢門的相關(guān)關(guān)系Table 3 Correlation between dominant phylum and functional genes involved in the main processes of nitrogen cycling
3 討論
3.1 退耕還草過程中植物與土壤性質(zhì)的變化
我們的結(jié)果顯示恢復(fù) 15年的草地植物生物多樣性最高,隨著恢復(fù)年限的增長,草地優(yōu)勢種發(fā)生變化,生物多樣性降低。與Zhang et al.(2016)和Wang et al.(2009)的研究一致,植物多樣性與豐富度在退耕后呈駝峰型變化,退耕還草后植物多樣性與豐富度持續(xù)增加,在植物群落演替中期物種豐富度最高,而后隨著退耕年限的增加而減少。這是由于恢復(fù)初期機(jī)會物種先占據(jù),隨后隨著累積資源的增加,物種豐富度增加。隨著生態(tài)系統(tǒng)的成熟,強(qiáng)大的競爭對手可能會在后期占據(jù)主導(dǎo)地位,導(dǎo)致物種豐富度下降,反映了植物在草地恢復(fù)過程中由于環(huán)境變化和種間競爭而導(dǎo)致群落結(jié)構(gòu)的變化(海旭瑩等,2020)。
退耕初期,因?yàn)橥V沽朔柿系妮斎耄寥鲤B(yǎng)分含量顯著降低。然而,隨著自然恢復(fù)年限的增加,研究發(fā)現(xiàn)地上植被發(fā)生次生演替,土壤有機(jī)殘?bào)w和根系分泌物的輸入量逐漸增加(Zhang et al.,2016),與該研究的結(jié)果一致,土壤中積累的養(yǎng)分水平(TC、TN)隨退耕年限逐漸增加,且蔗糖酶活性也逐漸增加,說明自然恢復(fù)導(dǎo)致了草地碳氮的積累。植被自然恢復(fù)過程中,植物的枯枝落葉可將部分養(yǎng)分歸還土壤,土壤碳含量積累可能歸因于凋落物以及根系的數(shù)量和質(zhì)量逐漸增多,促進(jìn)了土壤碳的礦化過程(Li et al.,2004);此外,隨著恢復(fù)年限的增加,與固氮作用有關(guān)的豆科植物增加促進(jìn)了氮的積累(張文彥等,2010);我們的結(jié)果均體現(xiàn)出植被自然恢復(fù)對土壤系統(tǒng)有積極影響。
3.2 退耕還草過程中細(xì)菌群落結(jié)構(gòu)的變化
地上植被演替影響植物根系的生長,引起土壤細(xì)菌群落組成和多樣性的變化。本研究中,沒有額外施肥的退耕草地放線菌門更豐富,這是由于放線菌門對營養(yǎng)物質(zhì)的貧乏有較高耐受性,且對干燥環(huán)境有較高抗性(Schimel et al.,2007)。植被自然恢復(fù)過程顯著增加了綠彎菌門的豐度,綠彎菌門是環(huán)境中C、N等元素生物地球化學(xué)循環(huán)的重要參與者,且與土壤中積累的養(yǎng)分水平(TC、TN)顯著相關(guān)。Tscherko et al.(2005)證明,在植被演替期間,土壤C和N資源對確定土壤微生物群落組成非常重要。Lozano et al.(2014)也報(bào)告了廢棄耕地細(xì)菌群落組成沿退耕年限順序的變化,顯示出植被自然恢復(fù)對地下微生物群落具有顯著作用。
隨著長期的植被自然恢復(fù),土壤營養(yǎng)資源不斷改善,為微生物提供了更多的可用生態(tài)位。關(guān)于植被演替對土壤細(xì)菌群落多樣性和結(jié)構(gòu)的影響的尚有爭議(Hossain et al.,2011;Lin et al.,2012)。我們的結(jié)果表明,隨著恢復(fù)時間的推移,細(xì)菌群落多樣性增加,且土壤細(xì)菌群落的α多樣性與植物群落的α多樣性之間有顯著相關(guān)關(guān)系,表明植物和微生物群落的演替過程是同步的,Zhang et al.(2016)的報(bào)道也支持了我們的結(jié)果。Lozano et al.(2014)發(fā)現(xiàn)了棄耕地土壤植被恢復(fù)過程中細(xì)菌群落演替階段之間的明顯分離,但與Kuramae et al.(2011)的研究結(jié)果相反,他們發(fā)現(xiàn)了大量的微生物群落重疊,在草原的時間序列上沒有明顯的差異。我們基于Bray-Curtis的NMDS結(jié)果與Lozano et al.(2014)結(jié)果一致,顯示細(xì)菌群落結(jié)構(gòu)沿時間序列發(fā)生了顯著變化。
相關(guān)分析和RDA表明,優(yōu)勢菌門(放線菌門、變形菌門、酸桿菌門和綠彎菌門)的豐度與土壤養(yǎng)分水平顯著相關(guān)。土壤總碳、總氮、碳氮比與銨態(tài)氮是影響黃土高原亞高山退耕還草自然恢復(fù)土地細(xì)菌群落結(jié)構(gòu)的主要環(huán)境因素。之前的研究表明,土壤銨態(tài)氮含量對土壤中主要細(xì)菌類群的相對豐度有正向影響(Yao et al.,2014;Yuan et al.,2014),合適的土壤碳氮比含量是微生物群落生長繁殖的驅(qū)動力,我們的結(jié)果也證實(shí)了這點(diǎn),支持了土壤碳、氮組分在細(xì)菌群落形成中的重要性。變形菌門中包含高比例的特定固氮根瘤菌,研究表明氮肥處理對細(xì)菌群落結(jié)構(gòu)的影響因研究地域不同存在差異(Bai et al.,2021)。我們的結(jié)果顯示恢復(fù)后期雖C、N積累,但變形菌門的豐度隨著自然恢復(fù)的時間序列減少,且豐度與土壤水分負(fù)相關(guān),與硝態(tài)氮和脲酶活性顯著正相關(guān),表明在亞高山干旱環(huán)境中變形菌門在氮代謝過程中產(chǎn)生了重要作用。此外,結(jié)果顯示酸桿菌門的Blastocatellia隨著恢復(fù)時間顯著增加,酸桿菌門屬于寡營養(yǎng)性型細(xì)菌(王光華等,2020),該發(fā)現(xiàn)與Campbell et al.(2010)報(bào)道的結(jié)果不一致,可能與亞高山特殊的地域環(huán)境(如低溫)有關(guān)。
3.3 退耕還草過程中細(xì)菌功能類群的變化
氮是土壤中重要的養(yǎng)分元素,土壤氮循環(huán)對土壤健康有重要的影響,土壤微生物在氮循環(huán)過程中發(fā)揮著重要的作用(賀紀(jì)正等,2013)。本研究中,由于農(nóng)田有額外的氮肥輸入,農(nóng)田土壤比恢復(fù)草地土壤中的氮代謝相關(guān)的基因更豐富,表明農(nóng)田土壤中固氮、硝化、反硝化作用較棄耕地土壤更強(qiáng)。氮主要通過生物固定進(jìn)入陸地生態(tài)系統(tǒng),nifH基因是固氮作用的標(biāo)志性基因,編碼固氮酶鐵蛋白組分(Wang et al.,2017;Nie et al.,2019)。在自然恢復(fù) 30年達(dá)到最高值,表明固氮能力隨著植被自然恢復(fù)時間的推移而加強(qiáng),我們的結(jié)果顯示土壤全氮含量與nifH基因豐度顯著相關(guān),這與在西藏高山草地土壤和長白山溫帶森林土壤中的研究結(jié)果一致(Yang et al.,2013;Tang et al.,2018)。amoA-AOA與amoA-AOB基因驅(qū)動土壤中氨氧化過程,該過程是硝化作用的第一步(Schmidt et al.,2019)。本研究發(fā)現(xiàn)amoA-AOA基因在草甸自然恢復(fù)20年達(dá)到最高值,amoA-AOB基因隨自然恢復(fù)顯著增加,且與總碳、總氮含量顯著正相關(guān),大部分研究結(jié)果均認(rèn)為底物濃度是影響氨氧化微生物在土壤中生長并發(fā)揮氨氧化功能的主要因素(Martens et al.,2009),本研究也證實(shí)了這一結(jié)果。同時,amoA-AOA和amoA-AOB基因與綠彎菌門、擬桿菌門和粘球菌門顯著正相關(guān),表明這3個優(yōu)勢門可能參與了氨氧化過程,nirS和nirK基因是將亞硝酸鹽還原為NO的功能基因,是反硝化作用的標(biāo)志性反應(yīng)(Wang et al.,2020)。本研究中隨著草甸自然恢復(fù)nirS和nirK基因增加不顯著((P>0.05)),表明亞高山草甸的自然恢復(fù)沒有顯著增加土壤的反硝化作用。研究發(fā)現(xiàn)土壤含氮量等是微生物反硝化過程的重要影響因素(Blaud et al.,2017;王楊,2014),本研究也發(fā)現(xiàn)nirS與總氮、硝態(tài)氮和銨態(tài)氮含量有顯著的相關(guān)關(guān)系。因此,亞高山退耕還草自然恢復(fù)過程不僅會改變微生物群落結(jié)構(gòu),還會通過微生物功能基因豐度的變化而顯著影響到土壤中的物質(zhì)循環(huán)過程(Zhang et al.,2007)。
4 結(jié)論
亞高山草甸自然恢復(fù)顯著改變了地上植被的多樣性,土壤的養(yǎng)分條件也隨著土壤有機(jī)殘?bào)w的積累顯著改善。植被恢復(fù)改善了亞高山的土壤養(yǎng)分資源,為微生物提供提了更多生態(tài)位,土壤細(xì)菌群落的多樣性沿自然恢復(fù)年限順序顯著增加,群落結(jié)構(gòu)也發(fā)生了顯著變化。同時,隨著地上植被的恢復(fù)演替,固氮作用的標(biāo)志性基因 nifH基因豐度顯著增加,參與氨氧化過程的amoA-AOA和amoA-AOB基因豐度也顯著增加,表明亞高山草甸自然恢復(fù)加強(qiáng)了土壤的固氮與硝化過程。通過對環(huán)境因子與細(xì)菌群落的結(jié)構(gòu)和功能基因分析證實(shí),土壤碳氮含量是影響亞高山退耕還草自然恢復(fù)土壤細(xì)菌群落的主要環(huán)境因素。本研究全面關(guān)聯(lián)了植被、土壤性質(zhì)、微生物群落結(jié)構(gòu)與氮循環(huán)相關(guān)功能基因,為制定亞高山退耕還草措施提供了數(shù)據(jù)依據(jù),并為亞高山草地生態(tài)恢復(fù)策略提供了理論支撐。
