辣椒CAMTA 基因家族生物信息學(xué)分析
2022-10-01 11:35:40蔣宏華李雪峰
中國蔬菜 2022年9期
關(guān)鍵詞:分析
蔣宏華 李 麗 李雪峰*
(1 湖南省農(nóng)業(yè)科學(xué)院蔬菜研究所,湖南長沙 410125;2 湖南省農(nóng)業(yè)科學(xué)院科研基地管理中心,湖南長沙 410125)
植物主要是通過分子機(jī)制來優(yōu)化生長和改善對環(huán)境約束的耐受性(Iqbal et al.,2020)。在生物和非生物脅迫下的植物生長發(fā)育中,鈣離子(Ca)作為第二信使在真核生物中普遍存在(Galon et al.,2010),且發(fā)揮重要作用(Kudla et al.,2010;Reddy et al.,2011)。鈣信號被認(rèn)為是許多適應(yīng)性過程和發(fā)育過程的核心調(diào)節(jié)信號,由鈣調(diào)素(CaM)等一系列鈣結(jié)合蛋白進(jìn)行傳輸和解析。這些蛋白質(zhì)通過調(diào)節(jié)轉(zhuǎn)錄因子(TFs)進(jìn)一步傳播信號以產(chǎn)生特定的下游反應(yīng)(Xiao et al.,2021)。目前,至少有90 種轉(zhuǎn)錄因子被鑒定為CaM結(jié)合蛋白,如CAMTA、MYB、WRKY、NAC、bZIP 和MADS-box 蛋 白(Reddy et al.,2002;Popescu et al.,2007;Kim et al.,2009;Galon et al.,2010)。在這些轉(zhuǎn)錄因子中,鈣調(diào)素結(jié)合轉(zhuǎn)錄激活因子(CAMTA)是1 個保守家族,也是與鈣調(diào)素相關(guān)的最具特征的轉(zhuǎn)錄因子(Bouché et al.,2005;Finkler et al.,2007;Xiao et al.,2021),在Ca/CaM 驅(qū)動的模式中通過調(diào)節(jié)植物應(yīng)激反應(yīng)和整體發(fā)育發(fā)揮了重要作用(Bouché et al.,2002;Galon et al.,2010;Liu et al.,2015;Shkolnik et al.,2019)。
CAMTA 轉(zhuǎn)錄因子家族具有相同的功能結(jié)構(gòu)域CG-1,這是1 個與DNA 結(jié)合有關(guān)的特異性結(jié)構(gòu)域,可以激活下游基因的轉(zhuǎn)錄。Tig(transcription factor immunoglobulin)參與TFs 中的非特異性DNA 相互作用(Aravind &Koonin,1999),也參與蛋白質(zhì)二聚化(Müller et al.,1995)。ANK 重復(fù)序列(ankyrin repeat)是多種真核生物蛋白質(zhì)中約33 個氨基酸的串聯(lián)重復(fù)模塊,與蛋白質(zhì)間的相互作用有關(guān)(Sedgwick &Smerdon,1999;Rubtsov &Lopina,2000)。串聯(lián)重復(fù)的IQ 基序(IQ motif)以不依賴于Ca的方式與CaM 相互作用。在擬南芥()受到低溫脅迫時,基因可誘導(dǎo)100 多個基因的表達(dá)(Vogel et al.,2005;Maruyama et al.,2010),其 中CAMTA 的CG-1 結(jié)構(gòu)域可以與基因啟動子中的CM2 順式作用元件特異性結(jié)合,調(diào)控的表達(dá),當(dāng)和發(fā)生功能突變時,會抑制的表達(dá);此 外,、和均可以誘導(dǎo)、和表達(dá),并增強(qiáng)植物的抗凍性(Doherty et al.,2009)。擬南芥中的已被證實(shí)與種子萌發(fā)早期的鈉(Na)穩(wěn)態(tài)有關(guān),突變體積累較少的NaCl,并表現(xiàn)出對鹽分和ABA 的耐受性(Shkolnik et al.,2019)。突變體損害了光合作用效率和水分利用效率,植物含水量相對較低,生長遲緩,易受干旱脅迫影響(Pandey et al.,2013)。在其他物種中,也廣泛參與了非生物脅迫的反應(yīng),在玉米()中,響應(yīng)低溫脅迫、鹽脅迫、干旱脅迫(Yue et al.,2015);在小麥()中,響應(yīng)低溫脅迫、高溫脅迫、鹽脅迫、干旱脅迫(Yang et al.,2020);草莓()在高溫、低溫、鹽脅迫下的表現(xiàn)也受到的調(diào)控(Leng et al.,2015)。
在擬南芥、水稻()等模式植物中均有報道,但在辣椒(L.)中還沒有進(jìn)行全面的研究。本試驗通過PlantTFDB 數(shù)據(jù)庫的信息獲得5 個辣椒CAMTA 家族基因成員,通過分析這5 個CAMTA 家族成員的蛋白理化性質(zhì)、蛋白保守結(jié)構(gòu)域、基因結(jié)構(gòu)及構(gòu)建系統(tǒng)進(jìn)化樹,同時分析其在幾種非生物脅迫與組織發(fā)育過程中表達(dá)量的變化等信息,為辣椒CAMTA家族基因在非生物脅迫與組織發(fā)育過程中信號轉(zhuǎn)導(dǎo)與調(diào)控表達(dá)的功能研究提供理論基礎(chǔ),為辣椒抗逆研究提供新思路。
1 材料與方法
1.1 試驗材料與基因表達(dá)量數(shù)據(jù)來源
試驗材料為辣椒高代自交系6421,由湖南省農(nóng)業(yè)科學(xué)院蔬菜研究所提供,對炭疽病、菌斑病和青枯病具有抗性。基因在非生物脅迫與不同組織發(fā)育階段的表達(dá)量數(shù)據(jù)下載于PepperHub(http://pepperhub.hzau.edu.cn/;Liu et al.,2017)。
1.2 試驗方法
1.2.1 基因表達(dá)量分析 ①激素和非生物脅迫處理。6421 種子用5%次氯酸鈉溶液表面消毒15 min,清水洗滌,然后播于裝滿蛭石的200 孔育苗盤中。將育苗盤放置在30 L 的避光塑料箱上,箱內(nèi)裝滿日本花園試驗營養(yǎng)液,pH 值6.0。幼苗生長溫度為25 ℃/18 ℃(晝/夜),光照/暗周期為16 h/8 h,相對濕度為60%~70%,光照強(qiáng)度為6 000 lx。
對播種40 d 后的幼苗進(jìn)行激素和脅迫處理,在營養(yǎng)液中添加最終濃度為10 μmol · L茉莉酸甲酯(MeJA)作為激素處理,在營養(yǎng)液中添加最終濃度為200 mmol · L的NaCl 作為鹽脅迫,在營養(yǎng)液中添加最終濃度為30 mmol · L的HO作為氧化脅迫,對照植株添加相應(yīng)量的營養(yǎng)液。低溫脅迫處理,將幼苗轉(zhuǎn)移到10 ℃恒溫的生長室中,光照/暗周期、相對濕度和光照強(qiáng)度與對照植株相同。采集脅迫處理后0、0.5、1、3、6、12、24 h 和對照植株葉片和根組織。考慮到生物鐘對植物基因表達(dá)的影響,所有處理和對照植株的樣本采集分別從脅迫處理第1 天的8:00、8:30、9:00、11:00、14:00、20:00 和第2 天的8:00 開始。每個樣品4 次生物學(xué)重復(fù),取樣后迅速放入液氮中冷凍,并在-80 ℃下保存,用于RNA 提取。
② 組織不同發(fā)育時期。幼苗生長溫度為白天25~29 ℃,夜間16~20 ℃。采用盆栽方式,每天通過滴灌系統(tǒng)補(bǔ)充復(fù)合肥料(N、P 和K)。為了采集不同發(fā)育階段的果實(shí)樣品,在開花當(dāng)天對第4 分杈處的花進(jìn)行標(biāo)記。開花后3、7、10、15、20、25、30、35、40、45、50、55、60 d 采集標(biāo)記花生長的果實(shí)。對于在開花后3、7 d 采集的果實(shí),將整個果實(shí)研磨以提取RNA;而對于在開花后10 d 及其后采集的果實(shí),將其切割并分為果肉、胎座和種子進(jìn)行研磨提取RNA。每個樣品由10個果實(shí)組成。根據(jù)花蕾的大小,在花蕾出現(xiàn)后2、5、10、15、20、25、30、40、50 d 收集花蕾,并將花蕾出現(xiàn)后60 d 時完全開放的花分為花瓣、帶柱頭的子房和雄蕊。在葉片出現(xiàn)后2、5、10、15、20、25、30、40、50、60 d 采集相同葉位的葉片樣本。每個樣品4 次生物學(xué)重復(fù),取樣后迅速放入液氮中冷凍,并在-80 ℃下保存,用于RNA 提取。
1.2.2 RNA 測序取樣本總RNA 0.5~5.0 μg,用作RNA 樣本制備的輸入材料。使用NEBNext Ultra Directional RNA Library Prep Kit(Illumina,#E7420L,NEB,USA)生成測序庫,并將索引代碼添加到每個樣本的屬性序列中。使用Qubit 3.0中的Qubit DNA 檢測試劑盒測定文庫濃度,然后將其稀釋至1 ng · μL。使用Bioanalyzer 2100 system(Agilent Technologies)評估文庫中的插入大小,并使用StepOnePlus Real-Time PCR System(Applied Biosystems,Carlsbad,CA,USA)準(zhǔn)確定量具有合格插入大小的文庫。使用HiSeq SR Cluster Kit v4 cBot HS(Illumina,San Diego,CA,USA)在cBot 群集生成系統(tǒng)上對索引庫進(jìn)行聚類。聚類生成后,使用150 bp 配對末端模型在Illumina Hiseq 4000 平臺上對文庫進(jìn)行測序。
1.2.3 mRNA 序列數(shù)據(jù)處理質(zhì)量控制 使用FastQC 軟件(Babraham Bioinformatics,http://www.Bioinformatics.Babraham.ac.uk)評估RNA 序列數(shù)據(jù)的質(zhì)量。
1.2.4 映射和表達(dá) 使用hisat2 程序(Kim et al.,2015)將所有干凈讀取的片段映射到辣椒的參考基因組(http://peppersequence.genomics.cn;Qin et al.,2014)Zunla 和CM334(http://peppergenome.snu.ac.kr;Kim et al.,2014)中,設(shè)置為“max intronlen 15000—dta-cuラinks—rna-strandness RF”(Kim et al.,2015)。就映射到染色體錨定基因組的讀取率而言,與基因組Zunla 的比對結(jié)果略好于CM334,因此在后續(xù)分析中選擇Zunla 基因組作為參考。提取所有與Zunla 對齊的唯一映射讀取,然后使用samtools 將其轉(zhuǎn)換為排序的bam 文件(Li et al.,2009)。通 過HTSeq(Anders et al.,2015)計算映射到每個基因外顯子的片段數(shù)量來量化基因表達(dá)。使用內(nèi)部perl 程序?qū)⒒蜷L度估計為所有外顯子的長度之和,使用每千個堿基每百萬個映射片段的片段數(shù)(fragments per kilobase million,F(xiàn)PKM)對每個基因的片段計數(shù)進(jìn)行歸一化,每個基因的表達(dá)水平計算為3 次生物復(fù)制的平均值。
1.2.5 鄰近法系統(tǒng)進(jìn)化分析 在PlantTFDB 網(wǎng)站(http://planttfdb.gao-lab.org/)得到辣椒5 個、擬南芥10 個、玉米10 個、水稻7 個、番茄()7 個CAMTA 基因家族的成員,使用MEGA 11 對得到的CAMTA 基因家族成員的氨基酸序列構(gòu)建系統(tǒng)進(jìn)化樹,方法為鄰近法(neighborjoining,NJ)。進(jìn)行Clustal W 多重序列比對,并采用自展法(Bootstrap)進(jìn)行檢驗,其中Bootstra 設(shè)置為1 000。
1.2.6 順式作用元件分析 使用Plant Care 網(wǎng)站(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)對辣椒的5 個CAMTA 家族基因的啟動子進(jìn)行在線預(yù)測,并篩選相關(guān)的順式作用元件,使用TBtools 對順式作用元件進(jìn)行可視化。
1.2.7 蛋白理化性質(zhì)與基因結(jié)構(gòu)分析 利用ExPASy Proteomics Sever(https://www.expasy.org/)對獲得的辣椒CAMTA 家族蛋白進(jìn)行理化性質(zhì)分析。利用GSDS(http://gsds.gao-lab.org/)對基因結(jié)構(gòu)進(jìn)行分析與可視化。使用MEME 網(wǎng)站(https://meme-suite.org/meme/tools/)對蛋白的保守序列進(jìn)行分析,并使用TBtools 進(jìn)行可視化。使用NCBI 網(wǎng)站(https://www.ncbi.nlm.nih.gov/)對蛋白序列進(jìn)行分析,并使用TBtools 對功能結(jié)構(gòu)域進(jìn)行可視化。
2 結(jié)果與分析
2.1 辣椒CAMTA 基因家族信息及蛋白理化性質(zhì)分析
辣椒5 個基因通過Protparam 進(jìn)行定位分析表明(表1):5 個基因分別分布在4 條染色體上,且大小差異明顯,基因全長最小,為270 bp;基因全長最大,為13 814 bp。CaCAMTAs 氨基酸數(shù)目在89~1 023 個之間;Capana01g002305 氨基酸數(shù)目最少,為89 個;Capana00g001755 氨基酸數(shù)目最多,為1 023 個。Capana01g002305 相對分子質(zhì)量最小,為10.686 35 kD;Capana00g001755 相對分子質(zhì)量最大,為115.200 58 kD。各基因的等電點(diǎn)分布較廣,在5.81~9.10 之間,且5 個蛋白的不穩(wěn)定系數(shù)均大于40,表明其編碼產(chǎn)物并不穩(wěn)定。
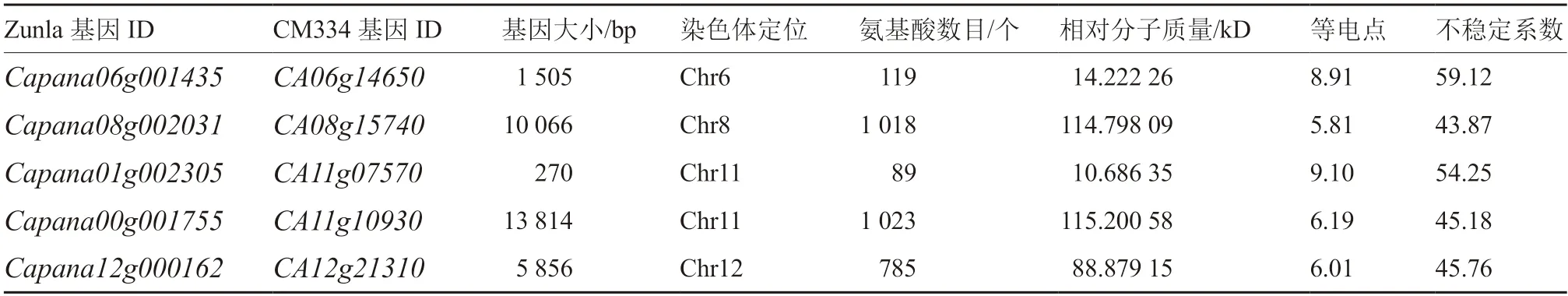
表1 辣椒CAMTA 基因家族信息及蛋白理化性質(zhì)
2.2 辣椒CAMTA 基因家族基因結(jié)構(gòu)分析
通過在線軟件GSDS 對辣椒CAMTAs 基因結(jié)構(gòu)進(jìn)行可視化分析(圖1),辣椒CAMTAs 基因不僅在長度上存在差異,而且在內(nèi)含子與外顯子數(shù)目上也存在差異。只有1 個外顯子,含有3 個外顯子,含有12 個外顯子,和均含有13 個外顯子。
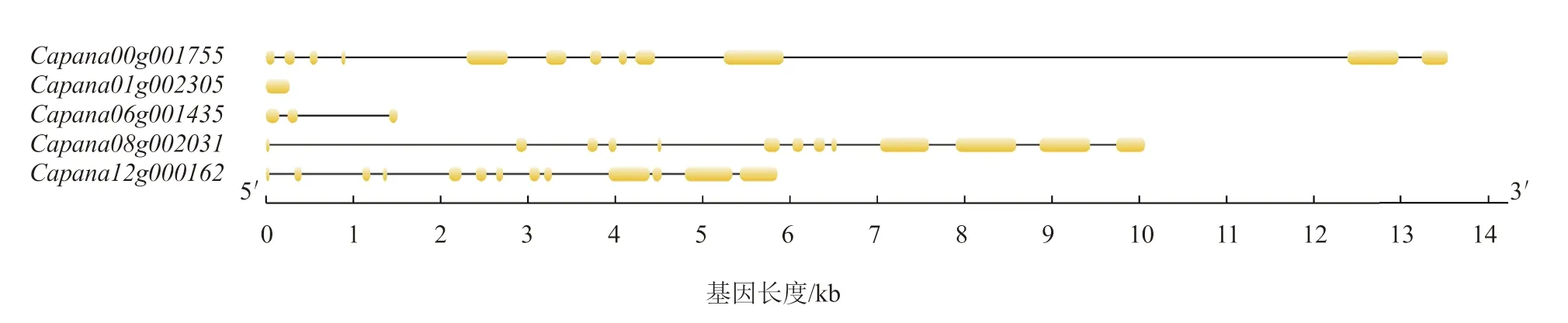
圖1 辣椒CAMTA 基因家族基因結(jié)構(gòu)分析結(jié)果
2.3 辣椒CAMTA 基因家族系統(tǒng)進(jìn)化、保守基序和結(jié)構(gòu)域分析
系統(tǒng)進(jìn)化樹分析顯示(圖2-A),辣椒CAMTA基因家族可以分為3 類,、和為I 類,為Ⅱ類,為Ⅲ類。
辣椒基因的蛋白質(zhì)序列中共鑒定獲 得5 個Motif(Motif1~Motif5),蛋白質(zhì)序 列相對保守(圖2-B),其位置與圖2-C 結(jié)構(gòu)域吻合。辣椒CAMTA 蛋白結(jié)構(gòu)域分析結(jié)果如圖2-C所示,該家族所有蛋白在N 端均含有CG-1 結(jié)構(gòu)域,是與DNA 結(jié)合有關(guān)的特異性結(jié)構(gòu)域,可以激活下游基因的轉(zhuǎn)錄。還有其他不保守的結(jié)構(gòu) 域,如Capana08g002031 和Capana00g001755共有的TIG 結(jié)構(gòu)域,Capana08g002031 獨(dú)有的CBD 結(jié)構(gòu)域,Capana00g001755 獨(dú)有的IQ 結(jié)構(gòu) 域,Capana08g002031、Capana00g001755 和Capana12g000162 共有的ANK 結(jié)構(gòu)域。
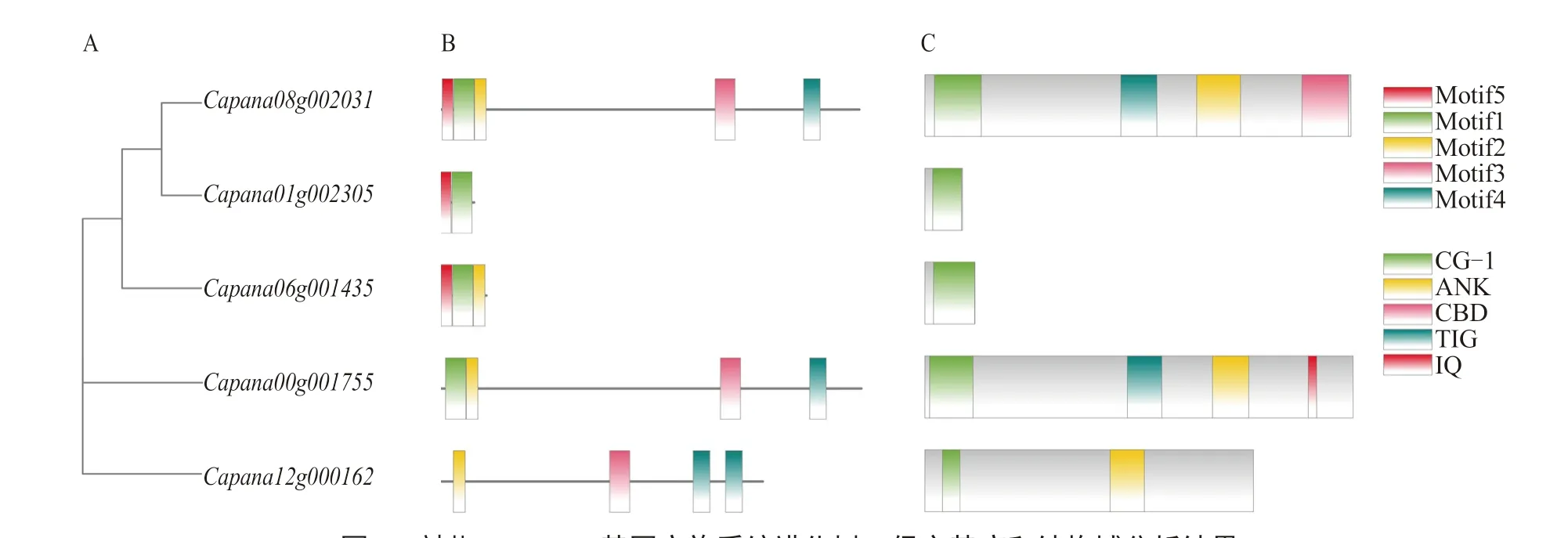
圖2 辣椒CAMTA 基因家族系統(tǒng)進(jìn)化樹、保守基序和結(jié)構(gòu)域分析結(jié)果
通過在線軟件Swiss Model 對CaCAMTAs 家族蛋白進(jìn)行三級結(jié)構(gòu)預(yù)測,結(jié)果如圖3 所示,辣椒CAMTA 家族蛋白構(gòu)象并不完全相同,這與蛋白質(zhì)的一級結(jié)構(gòu)和二級結(jié)構(gòu)有關(guān),其中兩條最小的蛋白Capana06g001435 和Capana01g002305 僅含有CG-1 結(jié)構(gòu)域,其三維構(gòu)象相仿。
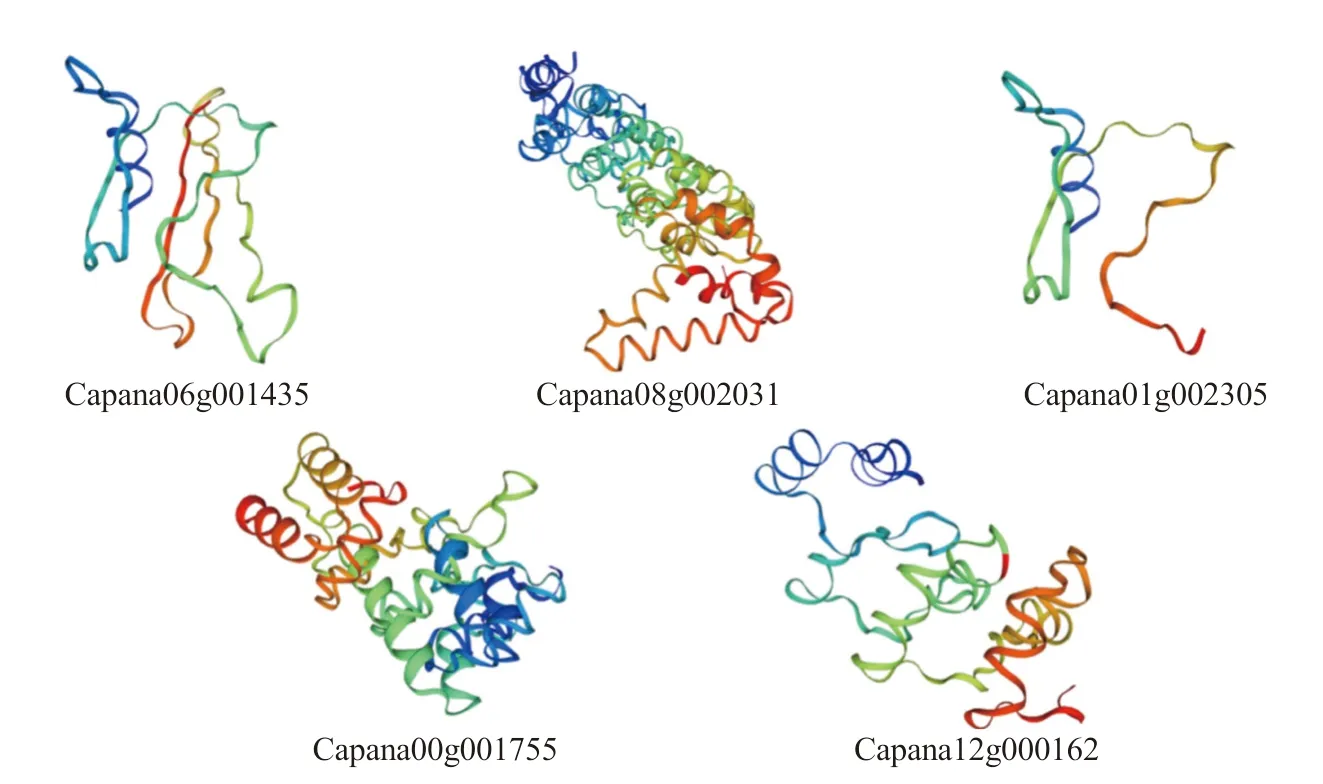
圖3 辣椒CAMTA 基因家族蛋白質(zhì)三級結(jié)構(gòu)預(yù)測
2.4 辣椒CAMTA 基因家族系統(tǒng)進(jìn)化分析
通過構(gòu)建系統(tǒng)進(jìn)化樹,將辣椒與擬南芥、玉米、水稻、番茄的CAMTA 基因家族進(jìn)行系統(tǒng)發(fā)育分析,結(jié)果如圖4 所示。辣椒的5 個CAMTA可以分為3 個亞家族,Capana08g002031、Capana06g001435 和Capana01g002305 分 為1 組,Capana00g001755與Capana12g000162各自為1組。辣椒Capana08g002031 與番茄Solyc01g105230 親緣關(guān)系最近,與擬南芥AT5G09410、AT5G64220 較近;辣椒Capana06g001435 與番茄Solyc01g057270親緣關(guān)系較近;辣椒Capana01g002305 與水稻Os07g43030、玉米GRMZM2G431243 親緣關(guān)系較近;辣椒Capana00g001755 與番茄Solyc05g015650親緣關(guān)系最近,與番茄Solyc12g035520、擬南芥AT1G67310 親緣關(guān)系較近;辣椒Capana12g000162與番茄Solyc12g099340 親緣關(guān)系最近,與擬南芥AT4G16150、AT3G16940 親緣關(guān)系較近。
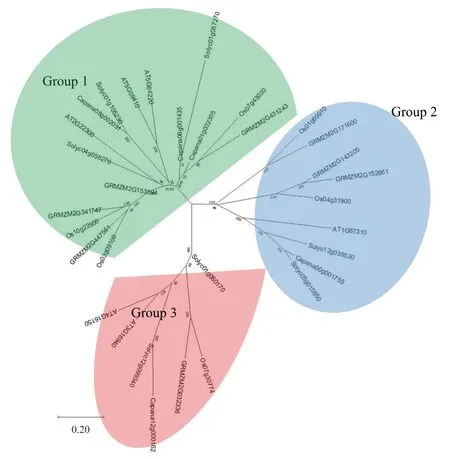
圖4 辣椒、擬南芥、玉米、水稻和番茄CAMTA 基因家族系統(tǒng)進(jìn)化分析結(jié)果
2.5 辣椒CAMTA 基因啟動子順式作用元件分析
通過辣椒CAMTA 基因啟動子順式作用元件分析,找出常見的順式作用元件,包括防御應(yīng)激反應(yīng)元件和脅迫響應(yīng)元件,結(jié)果如圖5 所示。辣椒CAMTA 家族基因有9 種順式調(diào)控元件,激素類的響應(yīng)元件主要包括水楊酸響應(yīng)元件(Salicylic acid responsiveness)、赤霉素響 應(yīng)元 件(Gibberellin-responsiveness)、茉莉酸甲 酯響應(yīng)元件(MeJA-responsiveness)、脫落酸響應(yīng)元件(Abscisic acid responsiveness)、生長素響應(yīng)元件(Auxin responsiveness),脅迫響應(yīng)元件主要包括厭氧誘導(dǎo)響應(yīng)元件(Anaerobic induction responsiveness)、防御和應(yīng)激響應(yīng)元件(Defense and stress responsiveness)、低溫響應(yīng)元件(Low temperature responsiveness)、干旱誘導(dǎo)的MYB 結(jié)合位點(diǎn)(MYB binding site)。表明辣椒基因與響應(yīng)辣椒的非生物脅迫關(guān)系密切。
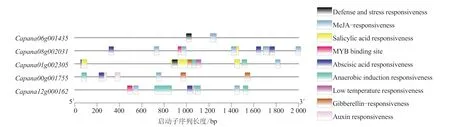
圖5 辣椒CAMTA 基因啟動子順式作用元件分析結(jié)果
2.6 激素和脅迫處理下辣椒CAMTA 基因表達(dá)分析
通過定量分析辣椒CAMTA 家族基因在激素和非生物脅迫處理下的表達(dá)情況,基因表達(dá)量變化如圖6 所示。在低溫脅迫下,與對照相比,表達(dá)量下調(diào),、在葉片中表達(dá)量上調(diào),在根中處理12 h 時下調(diào)表達(dá)。NaCl 脅迫下,在葉片中處理6 h 后就開始上調(diào)表達(dá),在根中處理24 h 時下調(diào)表達(dá),在葉片中處理3 h 后開始上調(diào)表達(dá),在葉片中處理6 h 后上調(diào)表達(dá),在根中處理1.5 h 后上調(diào)表達(dá)。HO脅迫下,表達(dá)量上 調(diào),表達(dá)量在葉片中處理6 h 后上調(diào),在根中處理3 h 時下調(diào)表達(dá),處理12 h 后上調(diào)表達(dá),變化不顯著。MeJA處理后,、與表達(dá)量均下調(diào),而與表達(dá)量低且變化不明顯。辣椒在低溫、NaCl、HO與MeJA處理后,、與表達(dá)量均有不同程度的變化,表明辣椒CAMTA 基因家族參與了脅迫后的信號轉(zhuǎn)導(dǎo)過程。
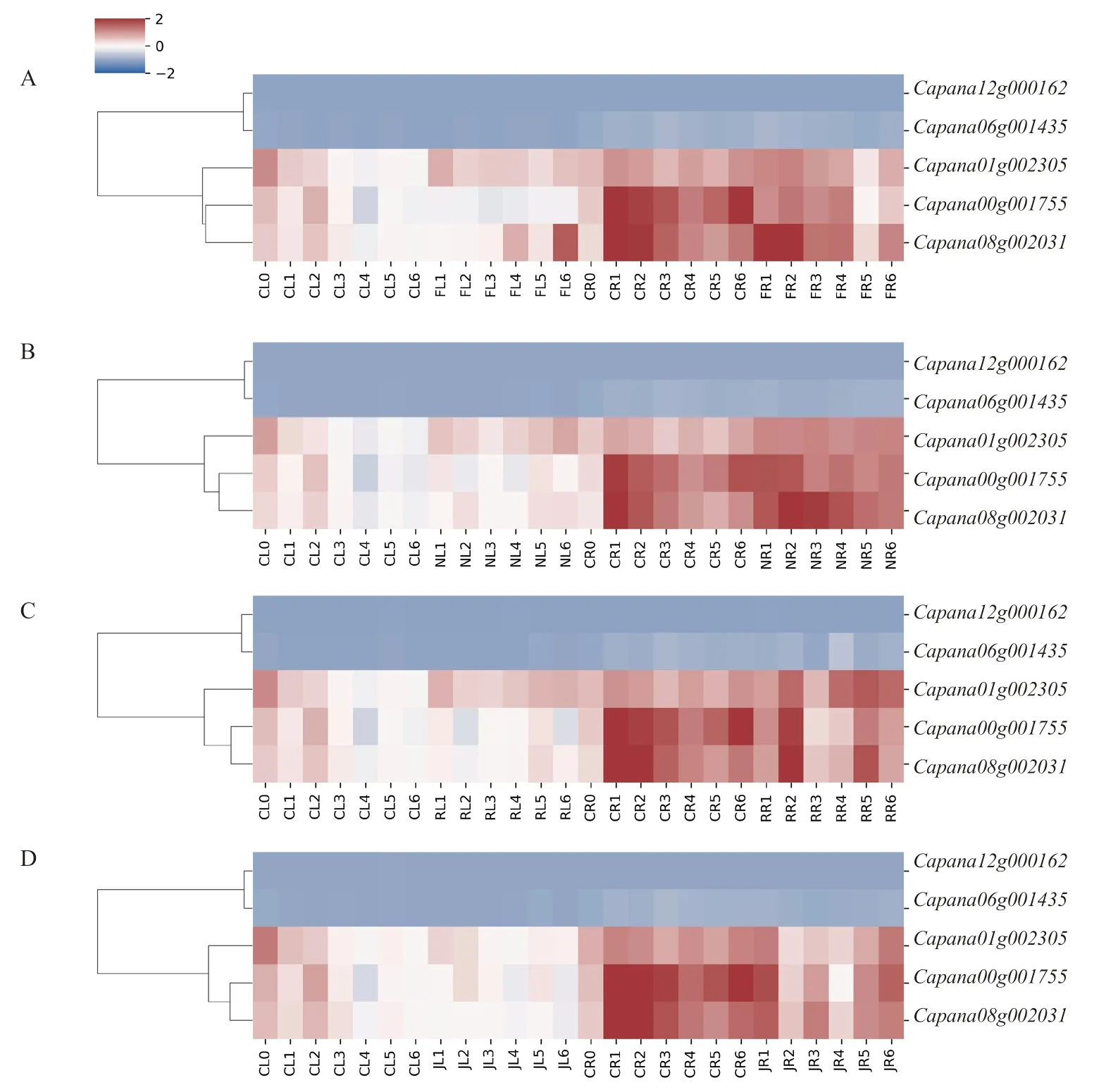
圖6 CaCAMTA 基因在不同處理后的表達(dá)量變化
2.7 組織不同發(fā)育時期辣椒CAMTA 基因表達(dá)分析
為探究辣椒CAMTA 基因家族是否在組織發(fā)育過程中發(fā)揮了作用,對辣椒在葉片、花、果實(shí)不同發(fā)育時期的表達(dá)量進(jìn)行了分析。在整個發(fā)育階段表達(dá)量低且無明顯變化,只在花蕾發(fā)育的2~10 d(圖7-A)、胎座發(fā)育的40 d(圖7-B)上調(diào)表達(dá),而在其他組織的發(fā)育過程中變化不明 顯。、和表達(dá)模式類似,在葉片、花蕾發(fā)育過程中有明顯的表達(dá)量變化,在子房、花瓣、雄蕊中的表達(dá)量依次降低(圖7-A)。在果肉、胎座、種子、早期的種子與胎座的發(fā)育過程中均有明顯的表達(dá)量變化,種子發(fā)育的前期表達(dá)量較高,隨后下調(diào)表達(dá),這表明、和對種子發(fā)育的調(diào)節(jié)作用主要發(fā)揮在早期(圖7-B)。
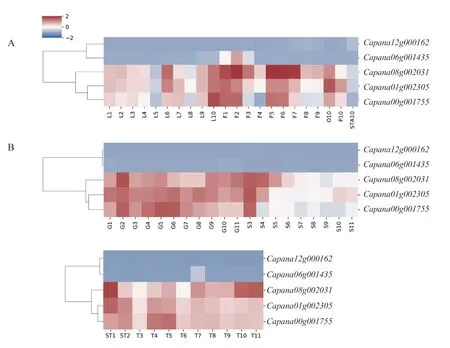
圖7 CaCAMTA 基因在葉片、花、果實(shí)中不同時期的表達(dá)量變化
3 結(jié)論與討論
了解植物對脅迫響應(yīng)的分子機(jī)制對分析植物的生命活動具有重要意義,并進(jìn)一步影響農(nóng)業(yè)生產(chǎn)。CAMTA 家族作為鈣調(diào)素轉(zhuǎn)錄激活因子,在植物受到脅迫信號轉(zhuǎn)導(dǎo)過程中必然發(fā)揮著重要的作用。5 個辣椒CAMTA 基因分布在4 條染色體上,基因大小差異極大,內(nèi)含子外顯子數(shù)目也不盡相同,表達(dá)的蛋白質(zhì)等電點(diǎn)變化范圍較大且不穩(wěn)定系數(shù)大于40,表明其表達(dá)產(chǎn)物并不穩(wěn)定,唯一共同點(diǎn)是其共有的CG-1 結(jié)構(gòu)域,可與DNA 特異性結(jié)合,并激活下游基因轉(zhuǎn)錄。對其啟動子分析發(fā)現(xiàn),CAMTA基因家族含有多個與脅迫相關(guān)的順式作用元件,表明其在脅迫響應(yīng)過程中的重要作用。在其他物種中的大量研究也已有證明。例如,在擬南芥中,、與協(xié)同工作,直接結(jié)合在的啟動子上誘導(dǎo)表達(dá),從而提高植物的抗凍性(Kim et al.,2013);也通過調(diào)節(jié)一些應(yīng)激反應(yīng)基因來積極應(yīng)對干旱反應(yīng),包括、(Pandey et al.,2013)。本試驗通過測定低溫、鹽、過氧化氫與茉莉酸甲酯處理后辣椒基因表達(dá)量的變化,表明在辣椒中CAMTA同樣參與了多種非生物脅迫的響應(yīng)。CAMTA 基因家族對脅迫的響應(yīng)并不局限于非生物脅迫,在非生物脅迫產(chǎn)生的調(diào)控過程中同樣發(fā)揮著重要作用,如可以作為植物免疫的負(fù)調(diào)節(jié)因子,通過激活介導(dǎo)的水楊酸(SA)信號來調(diào)節(jié)病原體防御反應(yīng)(Du et al.,2009);可能是小麥條銹病防御反應(yīng)的負(fù)調(diào)節(jié)因子,病毒誘導(dǎo)基因沉默(VIGS)后小麥的抗性增強(qiáng)。
本試驗中,對葉片、花、果實(shí)中不同發(fā)育時期的表達(dá)量變化分析表明,辣椒CAMTA 基因家族在組織的發(fā)育過程中同樣發(fā)揮了作用。在其他植物中,同樣有報道基因表達(dá)的空間差異與植物生長和發(fā)育有關(guān)(劉文宇 等,2021)。在擬南芥花粉發(fā)育過程中,和可以增強(qiáng)在花粉中的特異性表達(dá)(Li et al.,2005)。在煙草中過表達(dá)基因(),葉片和花瓣出現(xiàn)早衰的表型,這意味著CAMTA 參與了發(fā)育調(diào)節(jié)(Yang &Poovaiah,2000)。本試驗結(jié)果可以為辣椒CAMTA 基因家族成員的生物學(xué)功能研究與分子機(jī)制研究提供參考,同時為分子育種提供方向。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
