基于Cyt b序列的貴州境內(nèi)兩江流域月鱧群體遺傳結(jié)構(gòu)
2022-09-30 01:51:12吳俁學張顯波閔文武周其椿李世凱
貴州農(nóng)業(yè)科學 2022年9期
關鍵詞:貴州
吳俁學,張顯波*,閔文武,周其椿,李世凱
(1.貴州省水產(chǎn)研究所,貴州 貴陽 550025;2.貴州省特種水產(chǎn)工程技術中心,貴州 貴陽 550025)
0 引言
【研究意義】貴州位于我國西南部,地處云貴高原東側(cè)的大斜坡地帶,處于長江和珠江兩大流域上游交錯地段,河流數(shù)量較多,具有充沛獨特的水資源,加之豐富的地熱、余熱、冷泉水等資源以及特殊的喀斯特地形,孕育了種類繁多的水生生物,魚類區(qū)系組成復雜,具有豐富的生物多樣性。月鱧(Channaasiation)屬鱸形目(Perciformes)鱧科(Channidae)鱧屬(Channa),有七星魚、山花魚、點星魚、星光魚等俗稱,屬名貴淡水魚類,具有極高的經(jīng)濟價值和藥用價值,有保健魚之稱,為貴州原始土著魚類之一。開展貴州月鱧自然種群的遺傳背景研究,對貴州土著魚類資源保護和開發(fā)具有重要的現(xiàn)實意義。【前人研究進展】研究人員利用ITS序列和Cyt b序列分析比較了月鱧、烏鱧和斑鱧之間的進化關系[1-2],也有相關研究從形態(tài)學方面分析鱧屬魚類系統(tǒng)發(fā)育關系[3],主要是針對同屬不同種間的分析比較。Cytb基因是線粒體DNA上的編碼區(qū)之一,進化速率適中、易擴增,比較適合闡明種內(nèi)到種間的進化關系[4-6],廣泛地應用于各類物種的系統(tǒng)發(fā)育研究[7-9]。【研究切入點】目前尚無關于貴州月鱧群體遺傳學研究的相關報道,因此,以貴州境內(nèi)長江流域和珠江流域的4個月鱧群體為研究對象,測定和分析其Cytb基因序列,對月鱧群體的遺傳結(jié)構(gòu)進行研究。【擬解決的關鍵問題】弄清貴州境內(nèi)長江流域和珠江流域月鱧自然群體間的遺傳關系,探究貴州兩江流域月鱧自然種群的遺傳背景,為貴州土著魚類月鱧的種質(zhì)資源保護和開發(fā)利用奠定理論基礎。
1 材料與方法
1.1 材料
以貴州境內(nèi)長江流域和珠江流域分布的月鱧群體為主要研究對象,共收集到月鱧130尾,均為自然群體,具體采樣情況見表1。

表1 月鱧樣本采集情況Table 1 Sample information of C. argus populations
1.2 基因組DNA的提取與檢測
剪取月鱧樣本的尾鰭下葉組織,無水乙醇浸泡,—20℃保存?zhèn)溆谩⒄崭牧嫉谋椒覵DS法提取基因組DNA[10]。經(jīng)1%瓊脂糖凝膠電泳對提取的DNA進行完整性檢測,濃度及純度由NANODROP 2000分光光度儀測定,采用TE Buffer(pH=8.0)稀釋至60 ng/μL,—20℃儲存?zhèn)溆谩?/p>
1.3 線粒體Cyt b序列片段的擴增和測序
利用Primer blast設計該片段的引物,得到上游引物Cytb-CDS-F1:atggccaatctacgaaaaacac,下游引物Cytb-CDS-R1:ttaacctccgacgtccggtt,引物由生工生物工程(上海)有限公司合成。PCR反應在PTC-200型PCR儀上進行,反應體系:上下游引物各2 μL,模板DNA 4 μL,MIX 20 μL,加無菌去離子水至40 μL;反應程序:95℃預變性3 min;94℃變性45 s,55℃復性45 s,72℃延伸1 min,并進行35個循環(huán);72℃延伸10 min。擴增產(chǎn)物由生工生物工程(上海)有限公司測序。
1.4 數(shù)據(jù)分析
采用Clustal X軟件將測序結(jié)果進行對位排序,并輔以人工校對。利用MEGA 5.0軟件計算各種群間的遺傳距離,運用基于遺傳距離的鄰接法(Neighbor-joining method,NJ)和基于信息位點的最大簡約法(Maximum parsimonious criterion,MP)構(gòu)建系統(tǒng)進化樹,同時采用自舉檢驗估計系統(tǒng)樹中節(jié)點的置信度。用DNAsp5.0軟件對各群體的單倍型多樣性(Haplotype diversity,Hd)進行分析,估計不同種群的遺傳多樣性,并進行Tajima’s D檢驗統(tǒng)計。利用NCBI中 GenBank提供的生物信息資源共同進行數(shù)據(jù)分析。
2 結(jié)果與分析
2.1 4個月鱧群體的Cyt b序列特征
對貴州境內(nèi)長江流域不同區(qū)域的2個群體和珠江流域不同區(qū)域的2個群體共130尾月鱧的Cytb基因的946 bp序列分析顯示,A+T含量為53.7%,G+C含量為46.4%,其堿基組成差異不大。
長江流域烏江群體和清水江群體的堿基組成特征表現(xiàn)為保守位點940個,占核苷酸總數(shù)的99.37%,變異位點6個,其中,簡約信息位點4個,單態(tài)突變位點2個;轉(zhuǎn)換位點6個,無顛換位點。珠江流域貓營河群體和都柳江群體的堿基組成特征表現(xiàn)為保守位點939個,占核苷酸總數(shù)的99.26%,變異位點7個,其中,簡約信息位點7個,無單態(tài)突變位點;轉(zhuǎn)換位點6個,顛換位點1個。
2.2 4個月鱧群體的單倍型及核苷酸多樣性
2.2.1 群體的單倍型多樣性 從表2看出,長江流域和珠江流域4個月鱧群體130個樣本共定義6種單倍型,其中,以單倍型Hap 4的個體分布最多,4個群體無共享單倍型,清水江群體獨為1個單倍型。
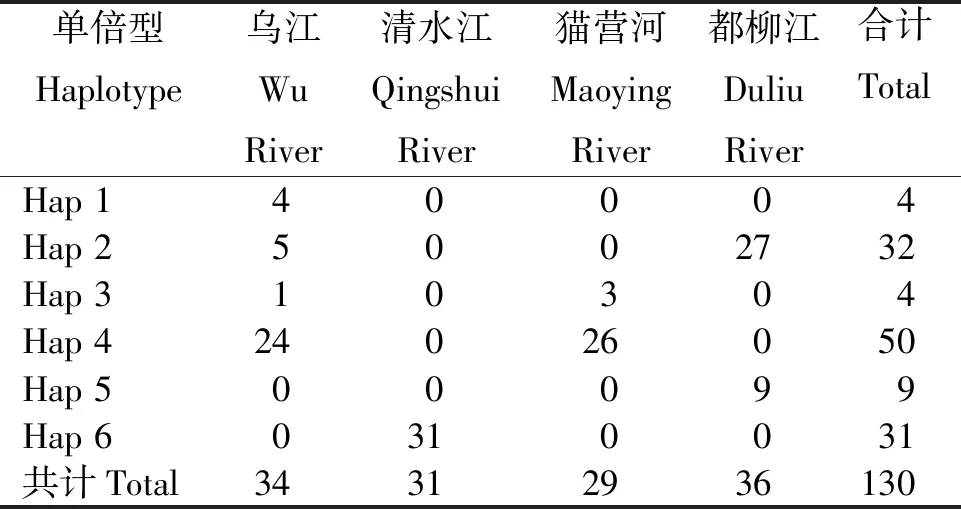
表2 4個月鱧群體Cyt b基因的單倍型分布Table 2 Distribution of the Cyt b haplotypes in C. argus population with four-month-age 尾
2.2.2 核苷酸多樣性 從表3看出,長江流域烏江群體34個樣本定義了4種單倍型,核苷酸多樣性指數(shù)(π)為0.001 65,單倍型多樣性指數(shù)(Hd)為0.480;長江流域清水江群體31個樣本僅定義1種單倍型。珠江流域貓營河群體29個樣本定義2種單倍型,π為0.000 61,Hd為0.192;珠江流域都柳江群體36個樣本定義2種單倍型,π為0.002 04,Hd為0.386。

表3 4個月鱧群體的遺傳多樣性指數(shù)Table 3 Genetic diversity indexes of C. argus population with four-month-age
4個月鱧群體總體的π為0.001 98,Hd為0.734。長江流域月鱧群體出現(xiàn)5種單倍型,珠江流域月鱧群體出現(xiàn)4種單倍型。其中,烏江群體的遺傳多樣性最豐富,有4種單倍型;清水江群體遺傳多樣性水平最低,其π和Hd均為0,且只有1種單倍型。
2.3 4個月鱧群體的遺傳距離與遺傳分化指數(shù)
2.3.1 遺傳距離 根據(jù)DNAsp的計算結(jié)果,4個月鱧群體間平均遺傳距離(D)為0.002 29。從表4看出,貓營河-都柳江群體的D值最大,為0.003 08;烏江-貓營河群體的D值最小,為0.001 31;烏江-清水江群體、烏江-貓營河群體、清水江-都柳江D值低于平均水平,其他群體間的遺傳距離值均大于平均遺傳距離;但各群體間D值均小于0.01,表明群體間遺傳差異較小,推測各群體間可能存在頻繁的基因交流[11]。

表4 基于序列分析月鱧群體間的遺傳分化Table 4 Genetic differentiation among C. argus populations based on sequence analysis
2.3.2 遺傳分化 參照群體遺傳分化標準(遺傳分化指數(shù)Fst>0.25)[12],長江流域群體與珠江流域群體間基因存在一定的遺傳分化。根據(jù)一般判斷標準(Fst<1)[12],群體間未出現(xiàn)生殖隔離,各群體間僅烏江-貓營河群體的基因流(Nm)>1。根據(jù)線粒體DNA堿基突變率每百萬年(Ma)1%~2.5%估算[13],種群間分化時間以貓營河-都柳江群體的分化時間最早(0.06 Ma~0.15 Ma),烏江-貓營河群體最晚(0.03 Ma~0.07 Ma),各群體間分化時間交叉在0.06 Ma~0.07 Ma。
2.4 4個月鱧群體的聚類
利用Genbank中斑鱧(Channamaculata,GenBank:MN057083.1)、烏鱧(Channaargus,GenBank:KU852447.1)作為同屬外群同源序列,對所有月鱧單倍型進行NJ聚類和UPMGA聚類(圖1)。從單倍型NJ樹可看出,6個倍型分別聚為2支,其中,Hap3、Hap4、Hap5聚為1支,Hap1、Hap2、Hap6聚為1支,最后2支聚在一起。以布展值50為劃分標準,長江流域與珠江流域月鱧種群內(nèi)部單倍型遺傳分化不明顯,結(jié)合各群體單倍型分布情況及各群體間遺傳距離的分析結(jié)果,長江流域及珠江流域月鱧群體之間種群分化不明顯。月鱧群體與同屬外種群明顯分開,但與斑鱧遺傳關系更近,與楊春英等[14]的研究結(jié)果一致。
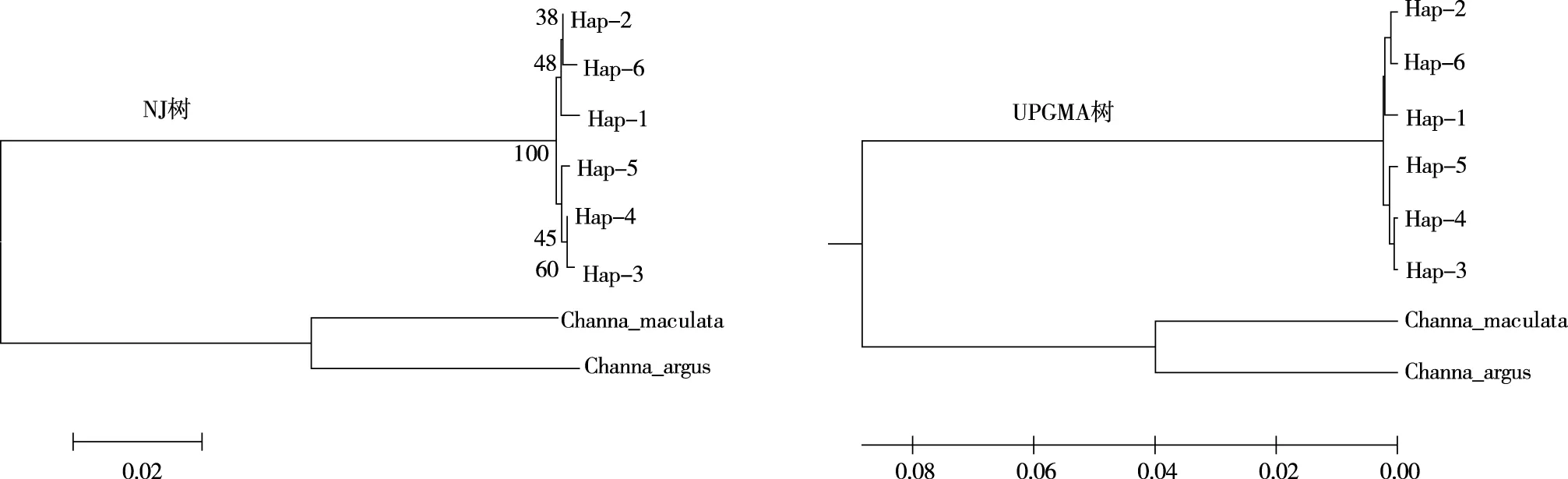
圖1 4個月鱧群體基于cytb單倍型的NJ樹及UPGMA樹Fig.1 NJ tree and UPGMA tree of C. argus population with four-month-age based on Cyt b haplotype
月鱧單倍型UPGMA樹聚類趨勢和NJ樹聚類趨同,均支持貴州月鱧種群的遺傳關系劃分為兩大支系。
3 討論
核苷酸多樣性指數(shù)是判斷群體遺傳多樣性水平的重要指標。研究結(jié)果顯示,貴州境內(nèi)長江流域(烏江、清水江)和珠江流域(都柳江、貓營河)4個月鱧群體遺傳多樣性水平呈烏江群體>貓營河群體>都柳江群體>清水江群體,總體呈長江流域>珠江流域。從單倍型數(shù)量上看,烏江群體出現(xiàn)4種單倍型,貓營河群體與都柳江群體均為2種單倍型,而清水江群體僅1種。根據(jù)GRANT等[15]對核苷酸多樣性指數(shù)(π)<0.005則群體遺傳多樣性水平低的判斷,貴州月鱧群體(π=0.001 98)整體遺傳多樣性處于較低水平。從近年來對相關水系魚類遺傳多樣性的研究結(jié)果看,貴州境內(nèi)月鱧遺傳多樣性低于小口白甲魚[16]、清水江鯉[17]、都柳江鲇、光倒刺鲃[18]等經(jīng)濟魚類。生物種群瓶頸效應、建群效應、棲息地破碎化以及遺傳漂變等因素都有可能引起生物遺傳多樣性偏低,20世紀90年代有水產(chǎn)科技人員指出,月鱧面臨資源衰減的困境[19]。根據(jù)本次對貴州月鱧采樣的分布結(jié)果看,月鱧的保有數(shù)量相比20年前大大減少,人類活動很大程度上影響月鱧種群數(shù)量,這有可能是造成月鱧遺傳多樣性水平低的原因。清水江月鱧種群的主要遺傳多樣性參數(shù)數(shù)值均為0,且倍型單一,除人類活動外可能與采集到的月鱧為稻田養(yǎng)殖區(qū)的樣本有關。黔東南地區(qū)月鱧主要在稻田養(yǎng)殖區(qū),稻田養(yǎng)魚相較河流有更大的局限性,造成月鱧群體的小范圍地理隔離,從而影響群體遺傳多樣性水平。目前沒有貴州月鱧遺傳多樣性的相關研究報道,隨著長江流域開展“十年禁捕”工作,相信月鱧資源會得到一定恢復,今后可適時增加樣品采集地以及使用其他分子標記如CoI、D-loop、ITS等再次對月鱧遺傳多樣性開展研究。
研究顯示,貴州境內(nèi)長江流域和珠江流域月鱧群體有一定的遺傳分化,但群體間遺傳距離不大,種群分化不明顯。根據(jù)種群間分岐時間估算,貴州月鱧群體間分化時間最早在15萬年前,晚于苗嶺山脈成為長江流域和珠江流域諸水系分水嶺的時間[20],說明地理隔離是造成兩江流域月鱧群體遺傳分化的重要因素。烏江月鱧群體和貓營河月鱧群體之間遺傳分化程度不高,存在一定程度的基因交流,這可能是因為貓營河處于珠江流域和長江流域分水嶺,兩流域水系時有相互掠水現(xiàn)象,地理隔離不明顯;也可能與貴州境內(nèi)曾經(jīng)開展過月鱧的人工養(yǎng)殖有關,推測造成兩群體遺傳分化的主要原因可能是生殖隔離。月鱧遺傳分化程度高,基因交流不頻繁,還可能與其生活習性相關。月鱧是廣溫、雜食性魚類,對環(huán)境適應能力強,游泳距離短,不需要生殖洄游、索餌洄游及越冬洄游,甚至在稻田里面就能生存,造成一定范圍的地理隔離和生殖隔離。
4 結(jié)論
采用PCR技術對貴州境內(nèi)長江流域(烏江、清水江)和珠江流域(都柳江、貓營河)的4個月鱧群體(共130尾月鱧樣本)進行遺傳關系及遺傳背景研究。結(jié)果表明,貴州月鱧Cytb基因946 bp序列的A+T含量為53.7%,高于G+C含量(46.3%),其堿基組成表現(xiàn)為C的含量最高、G的含量最低,呈較強的反G偏倚性。在130個月鱧樣本中存在6種單倍型,總體核苷酸多樣性指數(shù)為0.001 98,單倍型多樣性指數(shù)為0.734。4個月鱧群體間平均遺傳距離為0.002 29,以貓營河-都柳江群體的遺傳距離最大(0.003 08),烏江-貓營河群體的最小(0.001 31)。總體看,貴州兩江流域境內(nèi)月鱧遺傳多樣性水平低,長江流域和珠江流域月鱧各群體種群分化不明顯。可以利用各月鱧群體間遺傳分化程度高的有利條件,加強各群體間親本的交叉繁殖,盡量避免群體內(nèi)部近親繁殖,提升月鱧整體遺傳多樣性水平,達到保護貴州土著魚類資源及開發(fā)利用的目的。
猜你喜歡
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
鄉(xiāng)村地理(2019年2期)2019-11-16 08:49:28
鄉(xiāng)村地理(2018年3期)2018-11-06 06:51:08
鄉(xiāng)村地理(2018年3期)2018-11-06 06:51:06
鄉(xiāng)村地理(2018年3期)2018-11-06 06:50:10
貴茶(2018年6期)2018-05-30 09:53:50
貴茶(2018年6期)2018-05-30 09:53:36
鄉(xiāng)村地理(2018年4期)2018-03-23 01:54:04
鄉(xiāng)村地理(2016年2期)2016-06-15 20:29:24
貴州師范學院學報(2016年8期)2016-02-27 12:48:14
