植物乳桿菌ST-Ⅲ發酵苦蕎面團中蛋白質降解變化的研究
2022-09-28 04:55:42李云龍歐陽博雅周一鳴呂欣東佘宣明周小理
中國糧油學報 2022年8期
李云龍, 歐陽博雅, 周一鳴, 呂欣東, 向 茜, 佘宣明, 周小理,3
(山西農業大學;山西功能食品研究院1,太原 030031) (上海應用技術大學香料香精技術與工程學院2,上海 201418) (上海應用技術大學;美麗中國與生態文明研究院;上海高校智庫3,上海 201418)
酸面團是一種由谷物、水和乳酸菌、酵母菌等具有活性的微生物,經自然發酵或人工接菌制得的面團[1]。其發酵過程包括碳水化合物代謝、蛋白質代謝、胞外多糖的生成和揮發性代謝產物的生成等代謝反應[2,3]。尤其是乳酸菌通過發酵降解蛋白質所生成的小分子肽鏈、游離氨基酸等化合物,對改善面團的加工品質和產品的結構,尤其是焙烤類面制品,具有內部結構疏松細膩、風味豐富且令人愉悅,營養價值高并且在產品貨架期內不易產生雜菌等優點[4-6]。
苦蕎是一種富含多種營養成分和生物活性物質的雜糧,尤其是蛋白質質量分數較高,為8.5%~19%,高于玉米、水稻等作物,其中,苦蕎蛋白主要由清蛋白、球蛋白、谷蛋白和醇溶蛋白組成,清蛋白和球蛋白作為主要儲存蛋白,具有更高的營養價值和生物活性,而谷蛋白和醇溶蛋白主要與面制品的加工特性息息相關[7]。同時,相比于小麥來說,苦蕎氨基酸含量較高且配比更合理[8],其中不僅包含了人體所需的8種必需氨基酸,也富含有人體限制性氨基酸—賴氨酸,是大米、小麥的2倍以上,其生物價相當于脫脂奶粉生物價的92.3%、雞蛋全粉生物價的81.5%[9],因此,近年來苦蕎作為功能性營養食品的一種極好資源開始受到食品與營養界的極大關注[10,11]。然而,苦蕎不含麩質且纖維含量較高,其制備的餅干、面包等產品具有色澤暗淡、內部結構粗糙等缺點,將苦蕎粉應用于面制產品的制備中存在巨大挑戰[12,13]。發酵作為一種較好的谷物產品品質改良技術,目前在苦蕎發酵制品的基礎研究較少,已有研究表明,植物乳桿菌ST-Ⅲ是在酸面團發酵過程中分離出的一株具有代表性的乳酸菌,相比于其他乳酸菌來說,具有良好的耐酸和耐寒性,有利于加快發酵速度,適度的降解谷物中的蛋白質,提高發酵制品的品質及營養特性[14]。因此,本研究旨在利用植物乳桿菌ST-Ⅲ分別發酵小麥、苦蕎粉,通過研究植物乳桿菌ST-Ⅲ發酵面團比較小麥及苦蕎蛋白的降解和游離氨基酸的變化情況,為苦蕎酸面團發酵制品提供參考。
1 材料與方法
1.1 實驗材料
苦蕎粉(黑豐1號);高筋小麥粉;植物乳桿菌ST-Ⅲ(CGMCC NO.0847),乳業生物技術國家重點實驗室提供。
1.2 實驗試劑
鹽酸、氫氧化鈉、硫酸、硫酸銅、氯化鈉、甲基紅、酚酞、乙醇、葡萄糖、甲醇、乙腈、乙酸、溴酚藍、硼酸、考馬斯亮藍等試劑均為分析純。蛋白分子量標準(10~150 ku);SDS-PAGE凝膠電泳試劑。
1.3 主要儀器與設備
TECAN Infinite M200PRO 多功能酶標儀,LC-10 AVP PLUS 高效液相色譜,FD-2 冷凍干燥機,Chem Studio 化學發光凝膠成像系統,S-3400N 掃描電子顯微鏡,K280R 冷凍離心機,FE20 pH計,LDZF-50KB-Ⅲ型高壓式蒸汽滅菌鍋,SPX-100B-Z 醒發箱,HH-4 數顯恒溫水浴鍋,DYCZ-24DN 電泳儀。
1.4 方法
1.4.1 植物乳桿菌ST-Ⅲ的活化及生長曲線的測定
無菌條件下取植物乳桿菌ST-Ⅲ于MRS肉湯培養基,37 ℃下培養20 h后以5%的接種量轉接到75 mL的MRS肉湯培養基中,繼續于37 ℃下培養8 h。培養完成后用無菌生理鹽水以4 500 r/min離心洗滌2次備用。參考楊森[1]的方法,對植物乳桿菌ST-III的生長曲線進行測定。
1.4.2 植物乳桿菌ST-Ⅲ發酵面團的制備
參考楊森[1]制備乳酸菌發酵酸面團的方法,按原料粉∶菌液質量比=2∶1的比例分別稱取小麥粉、苦蕎粉、混合粉(小麥粉∶苦蕎粉質量比=1∶1)和洗滌后的植物乳桿菌菌液(含量為8.0 logCFU/g),混勻后放入已滅菌的發酵罐中,于30 ℃醒發箱中進行發酵后,得到小麥發酵面團(WS)、苦蕎發酵面團(BS)、和混合發酵面團(WBS)。
1.4.3 面團發酵過程中中菌落密度的變化
參考楊森[1]的方法,于植物乳桿菌ST-Ⅲ發酵20 h內每隔2 h取樣3種酸面團,進行植物乳桿菌ST-Ⅲ菌落密度的生長計數測定。
1.4.4 面團發酵過程中酸化能力的研究
參照Bartkiene等[15]的方法,于植物乳桿菌ST-Ⅲ發酵20 h內每隔2 h取樣進行pH值和總酸的測定;參照高世陽[16]和張思佳等[17]的方法,于植物乳桿菌ST-Ⅲ發酵24 h內每隔6 h取樣,測定其中乳酸與乙酸的含量。
1.4.5 面團發酵過程中蛋白質各組分含量的測定
參考楊森[1]的方法,于植物乳桿菌ST-Ⅲ發酵24 h內每隔2 h取WS、BS和WBS各1 g,依次于不同溫度下溶于5 mL 1mol/L NaCl與50 mmol/L Tris-HCl(pH=8)緩沖液、5 mL 70%乙醇和5 mL 0.05 mol/L冰乙酸中,分級提取得到清蛋白、醇溶蛋白和谷蛋白,采用Bradford法[18]測定3種蛋白質含量。
1.4.6 面團發酵過程中蛋白各組分SDS-PAGE
參考楊森[1]的方法,利用SDS-PAGE凝膠電泳試劑盒對植物乳桿菌ST-Ⅲ發酵0、6、12、18、24 h時,WS、BS和WBS中的全蛋白、清蛋白和醇溶蛋白的分子量分布進行測定,凝膠染色及脫色后,拍照保存,圖像利用Quantity One軟件進行分析。
1.4.7 面團發酵過程中游離氨基酸含量的測定
根據SDS-PAGE實驗,選取WS、BS和WBS樣品各10 g,并采用張思佳等[17]的方法對樣品中的游離氨基酸含量進行測定。
1.5 數據處理
采用SPSS 20.0處理數據,每組數據重復3次,取其平均值,并進行顯著性分析,P<0.05表示差異顯著。同時,采用Origin 2019繪圖。
2 結果與分析
2.1 植物乳桿菌生長曲線及乳酸菌發酵面團菌落密度的變化
植物乳桿菌ST-Ⅲ在MRS肉湯培養基中的生長曲線如圖1所示,經過6 h的延滯期后在6~14 h進入對數期,快速生長,14 h后進入穩定期。微生物生長曲線的測定可以為發酵面團制備過程中植物乳桿菌的接種和乳酸菌在面團發酵過程中的生長分析提供依據[19]。3種發酵面團中植物乳桿菌ST-Ⅲ的接種量均為8.0 logCFU/g發酵面團。植物乳桿菌ST-Ⅲ經歷了2 h的延滯期后進入對數生長期;在穩定期,WS、BS和WBS中的植物乳桿菌ST-Ⅲ菌體密度分別達到11.751、12.410 、12.079 logCFU/g發酵面團;進入對數生長期后,相較于WS而言,植物乳桿菌ST-Ⅲ在BS和WBS中的生長更加旺盛,這可能是由于對數生長期時BS和WBS的酸性環境更加適宜植物乳桿菌ST-Ⅲ的生長,因而促進了植物乳桿菌ST-Ⅲ的繁殖代謝;發酵后期,植物乳桿菌ST-Ⅲ因3種酸面團中的營養物質缺乏而生長緩慢[20]。
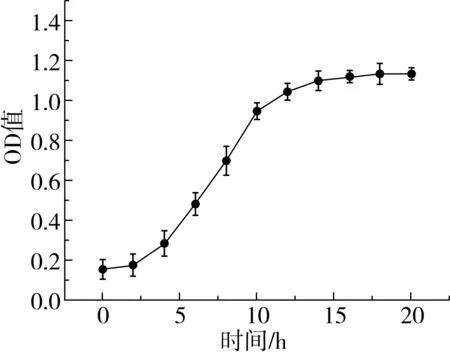
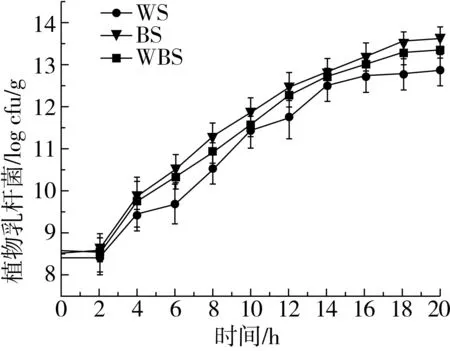
圖1 植物乳桿菌ST-Ⅲ生長曲線和酸面團中乳酸菌菌落密度變化
2.2 面團發酵過程酸化能力的變化
乳酸菌發酵面團中乳酸菌的生長可以使面團pH降低,總酸度升高[21]。綜合圖1和圖2可以看出,BS為弱酸性面團,經植物乳桿菌ST-Ⅲ發酵后,3種酸面團pH均隨著發酵時間的增加逐漸下降,TTA則相反。其中,在發酵初期3種酸面團酸化速率均較慢。但隨著植物乳桿菌ST-Ⅲ的不斷擴增,產酸速率加快,當發酵6~12 h時,BS的pH急劇下降,總酸度急劇上升,且下降變化顯著快于WS、WBS,這可能是由于處于對數生長期的植物乳桿菌ST-Ⅲ對苦蕎基質具有更強的適應性,因此代謝更為旺盛。當發酵進行18 h后,隨著底物不斷消耗,乳酸菌的生長和代謝受到抑制,3組酸面團pH均趨于穩定,但其TTA仍呈現緩慢上升趨勢,直至發酵20 h,WS、BS和WBS的TTA分別為16.28、24.39、22.26 mL,可能是由于苦蕎發酵基質灰分含量較高,對酸具有較強的緩沖能力,因此BS的TTA顯著高于WS、WBS。
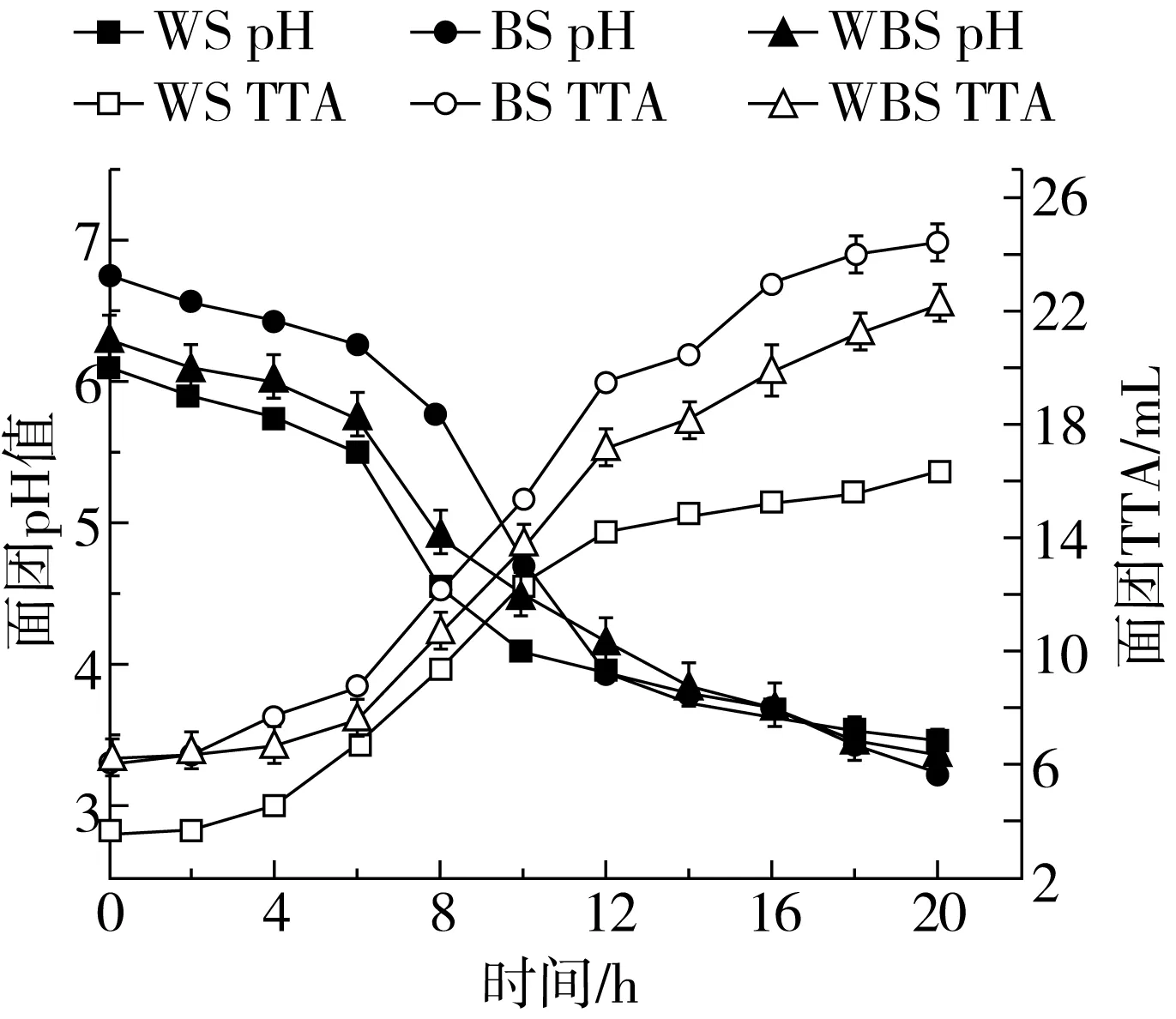
圖2 植物乳桿菌ST-Ⅲ發酵酸面團pH值與TTA的變化
2.3 面團發酵過程中有機酸含量的變化
植物乳桿菌ST-Ⅲ為兼性發酵乳桿菌,優先進行糖酵解途徑,即在發酵過程中利用葡萄糖產生乳酸,而后利用戊糖進行戊糖磷酸途徑產生乙酸[22]。圖3表明,BS中的乙酸含量在發酵12 h后分別為WS和WBS中乙酸含量的1.89、1.28倍;而BS中乳酸含量在發酵12 h分別為WS和WBS中乳酸含量的2.62、1.24倍,此現象與植物乳桿菌ST-Ⅲ的生長曲線及其在3種發酵面團的過程中其菌落密度變化趨勢有一定的聯系。植物乳桿菌ST-Ⅲ在發酵WS的過程中所產生的乳酸和乙酸含量均低于BS和WBS,其中,發酵24 h后BS中的乙酸含量分別為WS與WBS中乙酸含量的1.61、1.06倍;而BS中乳酸的含量分別為WS與WBS中乳酸含量的1.75、1.15倍。由植物乳桿菌ST-Ⅲ在發酵3種發酵面團過程中有機酸含量的變化,可推測BS中的蛋白質在發酵過程中能夠被更加廣泛地被水解,可能引起BS中游離氨基酸含量相對增幅較大。
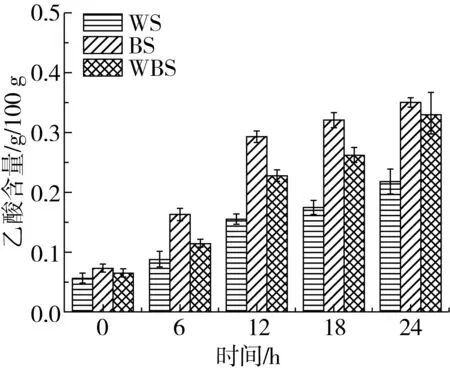
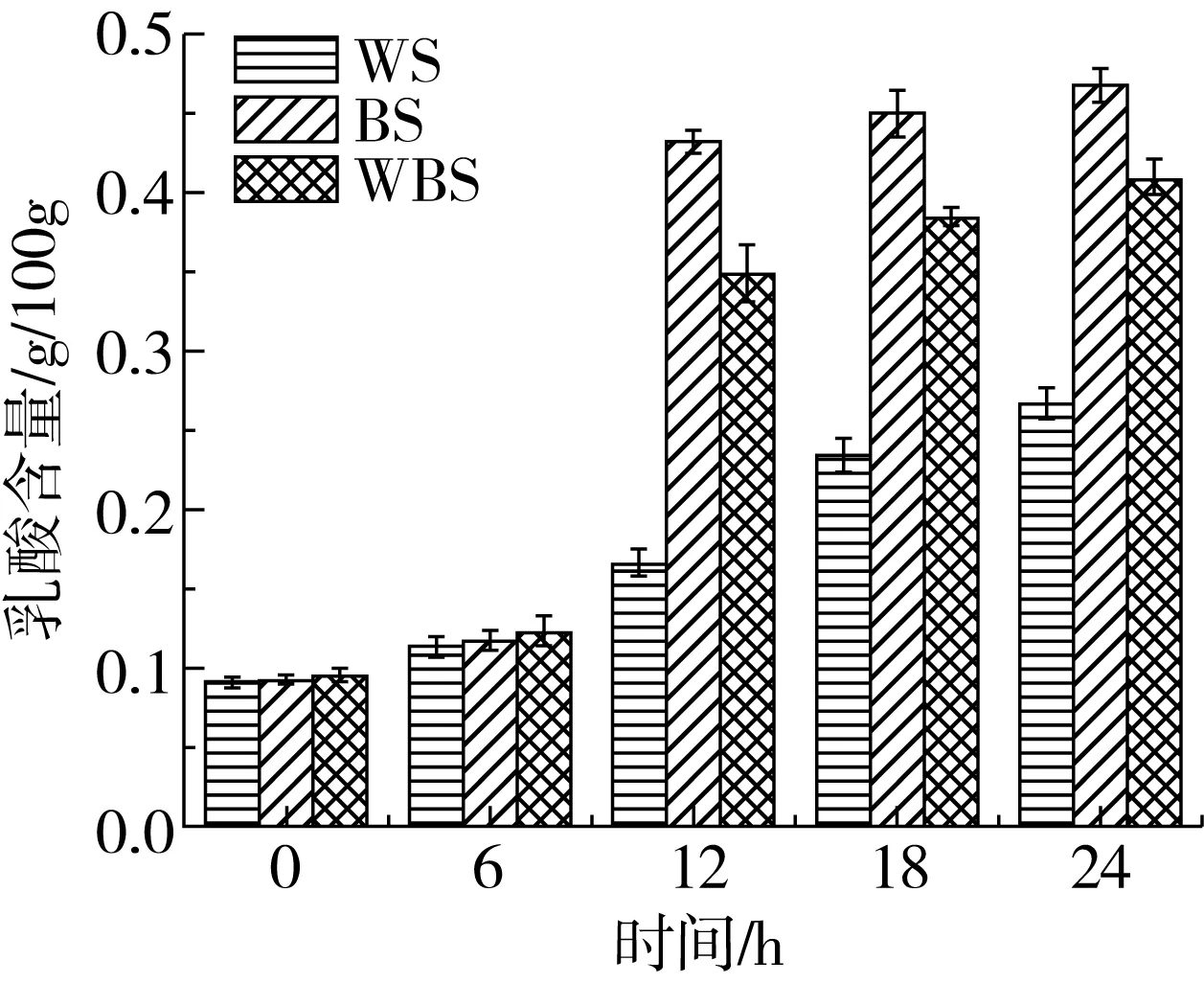
圖3 植物乳桿菌ST-Ⅲ發酵酸面團中乙酸與乳酸含量的變化
2.4 蛋白質各組分含量變化
酸面團對面團中蛋白質降解程度不同,這可能是乳酸菌發酵不同谷物基質過程中,代謝產生有機酸的種類和含量差異造成的[23]。由表1可知,WS、BS、WBS中清蛋白,醇溶蛋白,谷蛋白含量隨著發酵的進行具有顯著差異。在發酵過程中,各組面團中的清蛋白含量均呈現下降的趨勢。經過24 h的發酵,BS、WBS的清蛋白分別有30%、15.8%被降解,而WS的清蛋白則有90%被降解。植物乳桿菌ST-Ⅲ在發酵過程中對小麥面團中可溶性蛋白具有極強的降解作用,且對酸面團中清蛋白的降解主要集中在4~16 h。而含有苦蕎的酸面團中清蛋白隨著發酵時間的延長,降解速率較慢。
與清蛋白相比,WS、BS和WBS中的醇溶蛋白含量隨著3種酸面團的發酵均呈現出先增加后減少的趨勢。3種酸面團中醇溶蛋白的含量隨著植物乳桿菌ST-Ⅲ發酵的進行均有不同程度的增長,這一增長趨勢分別持續至WS發酵至16 h時,而當BS和WBS發酵至18 h時,醇溶蛋白含量才呈現下降的趨勢。在發酵24 h后,WS、BS和WBS中醇溶蛋白較發酵初期上升,這可能是由于植物乳桿菌ST-Ⅲ的繁殖和代謝產酸促進了酸面團中諸如清蛋白等可溶性蛋白的溶解從而導致醇溶蛋白含量的增加[24]。由于醇溶蛋白的變化可能對面團的延伸性具有較大的影響,因此從醇溶蛋白的變化趨勢可推測,利用植物乳桿菌ST-Ⅲ發酵苦蕎酸面團能夠改善苦蕎面制品的加工品質。
面團發酵過程中乳酸菌的代謝產物對面團中谷蛋白具有較強的降解作用,對面團的勁度、韌性有較大的影響[24]。植物乳桿菌ST-Ⅲ發酵對3種發酵面團中的谷蛋白均有著明顯地降解作用,且隨發酵時間的進行具有顯著差異(P<0.05);經過24 h的發酵后,WS中4.38%的谷蛋白被降解,而BS和WBS中的谷蛋白則分別被降解了4.77%、4.45%。
2.5 面團發酵過程中全蛋白SDS-PAGE圖譜變化
利用Quantity One對圖4的SDS-PAGE譜圖進行分析得到蛋白質的質量分數(表2)。植物乳桿菌ST-Ⅲ發酵過程中,3種發酵面團中全蛋白含量均呈現下降趨勢,這表明植物乳桿菌ST-Ⅲ發酵苦蕎酸面團有助于將大分子苦蕎蛋白降解為小分子,從而提高苦蕎蛋白的利用率。結合圖1可知,植物乳桿菌ST-Ⅲ在WS和WBS發酵至第6 h時開始劇烈繁殖代謝,而其在BS發酵第8 h后開始代謝產酸。BS中的苦蕎蛋白,尤其是36.5 ku左右的苦蕎蛋白在植物乳桿菌ST-Ⅲ發酵苦蕎酸面團的過程中被降解地較為徹底,而WS與WBS中的蛋白質被降解的幅度則相對較小,這驗證了BS的弱酸性環境更加適宜植物乳桿菌ST-Ⅲ的生長繁殖代謝,進一步使其產有機酸量增加,從而增進了對蛋白質的降解。
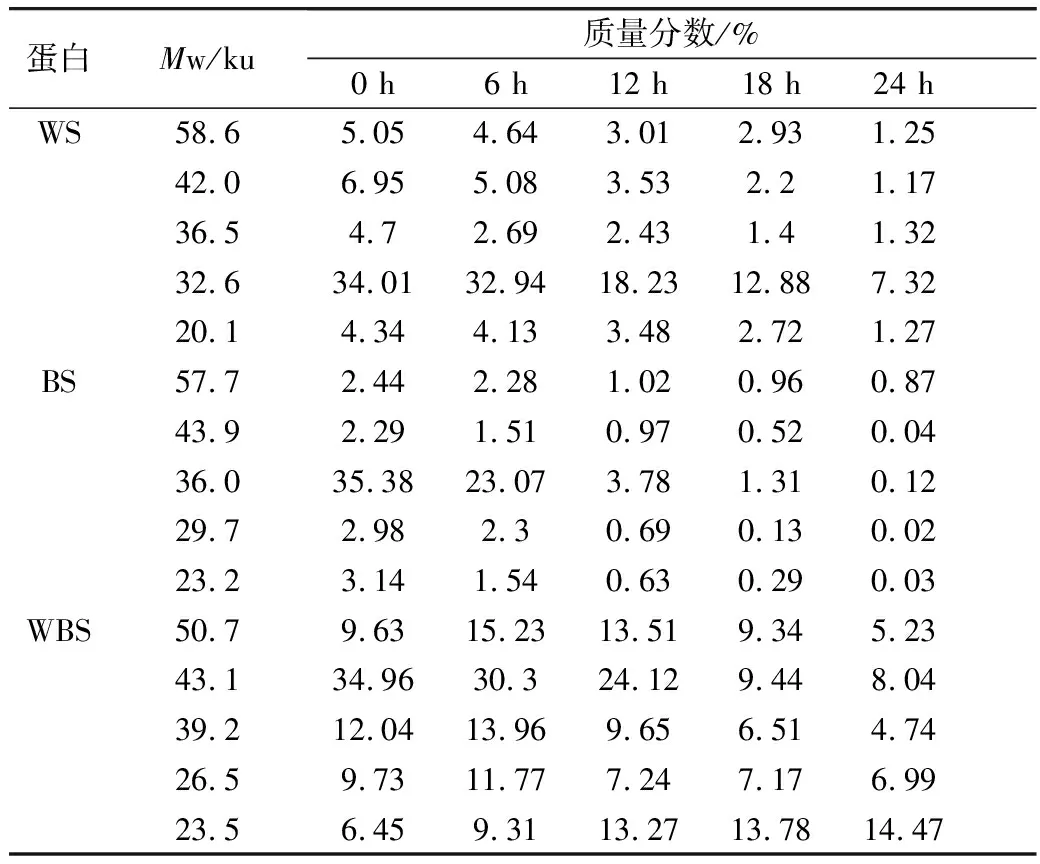
表2 植物乳桿菌ST-Ⅲ發酵3種發酵面團中全蛋白質量分數變化
2.6 植物乳桿菌發酵酸面團過程中清蛋白和醇溶蛋白SDS-PAGE圖譜變化
對圖5的SDS-PAGE譜圖進行分析得到蛋白質質量分數(表3)。經乳酸菌發酵后,苦蕎清蛋白分子量集中在23~56.7 ku之間,即1條組成2S蛋白的單鏈多肽鏈[25]。在植物乳桿菌ST-Ⅲ發酵3種酸面團的過程中,面團中的各主要分子段的清蛋白均被降解,同時結合表1來看,發酵12 h時清蛋白降解速度變緩。其中,BS中的清蛋白,尤其是36.2 ku段的蛋白質在發酵24 h后從35.38%降低至3.86%,而29.7 ku段的蛋白質則從19.01%被降解至0.07%。23.0 ku段的分子則出現積聚;由此可見,在植物乳桿菌ST-Ⅲ發酵苦蕎面團有助于將面團中的部分大分子蛋白降解為小分子蛋白,從而促進人體對苦蕎蛋白的吸收。
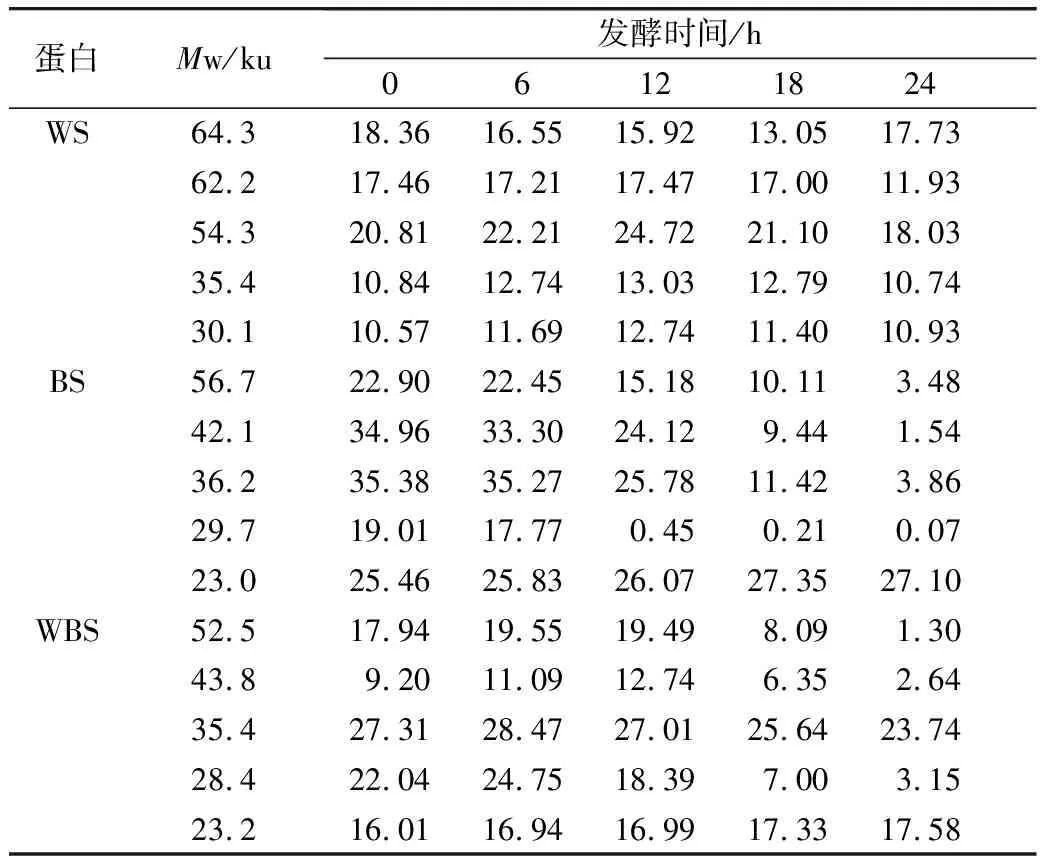
表3 植物乳桿菌ST-Ⅲ發酵WS過程中清蛋白質量分數變化/%
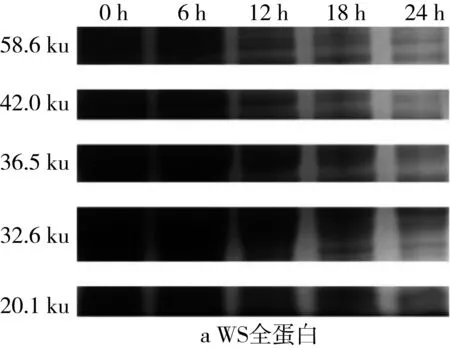
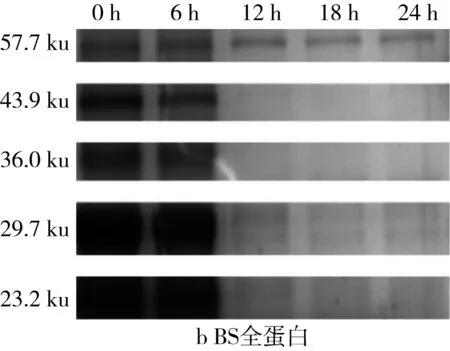
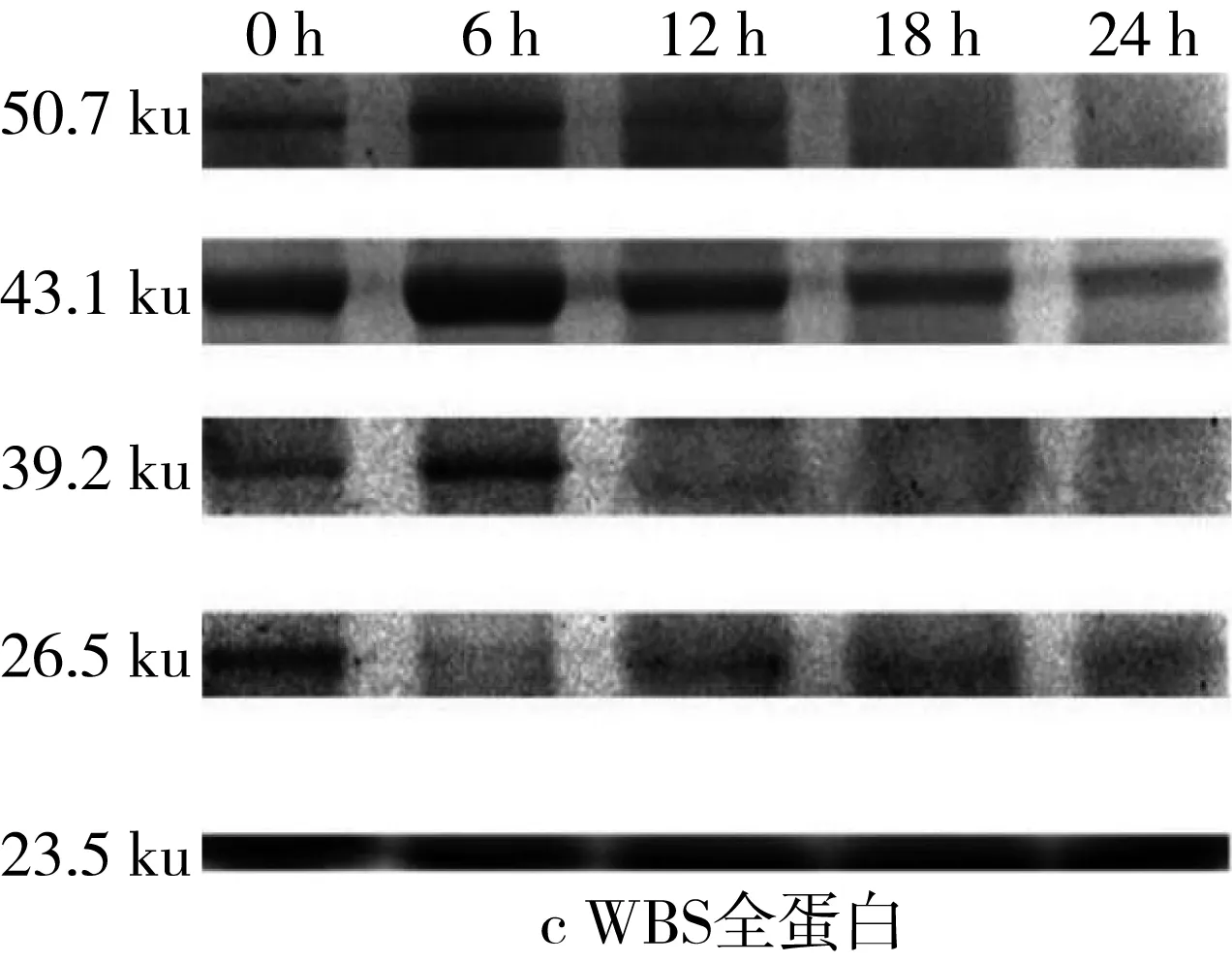
圖4 植物乳桿菌ST-Ⅲ發酵酸面團過程中全蛋白SDS-PAGE
由于苦蕎中醇溶蛋白含量極少,因此本研究旨在探討乳酸菌發酵對于小麥、苦蕎中醇溶蛋白含量變化的影響。由于醇溶蛋白通過Quantity One對圖6的SDS-PAGE譜圖進行分析從而得到蛋白質的質量分數(表3)。發酵BS過程中,37.2 ku段的蛋白質在發酵12 h時出現積聚,而78.8、62.4、48.3k、33.2 ku段的蛋白質在出現短暫積聚后繼續被降解。對于WS而言,32.9 ku段的蛋白質在發酵6 h時出現積聚,而其他大分子段蛋白均被緩慢降解。但相較于全蛋白和清蛋白而言,WS、BS、WBS的醇溶蛋白質量分數隨著發酵時間的延長變化較為微小。
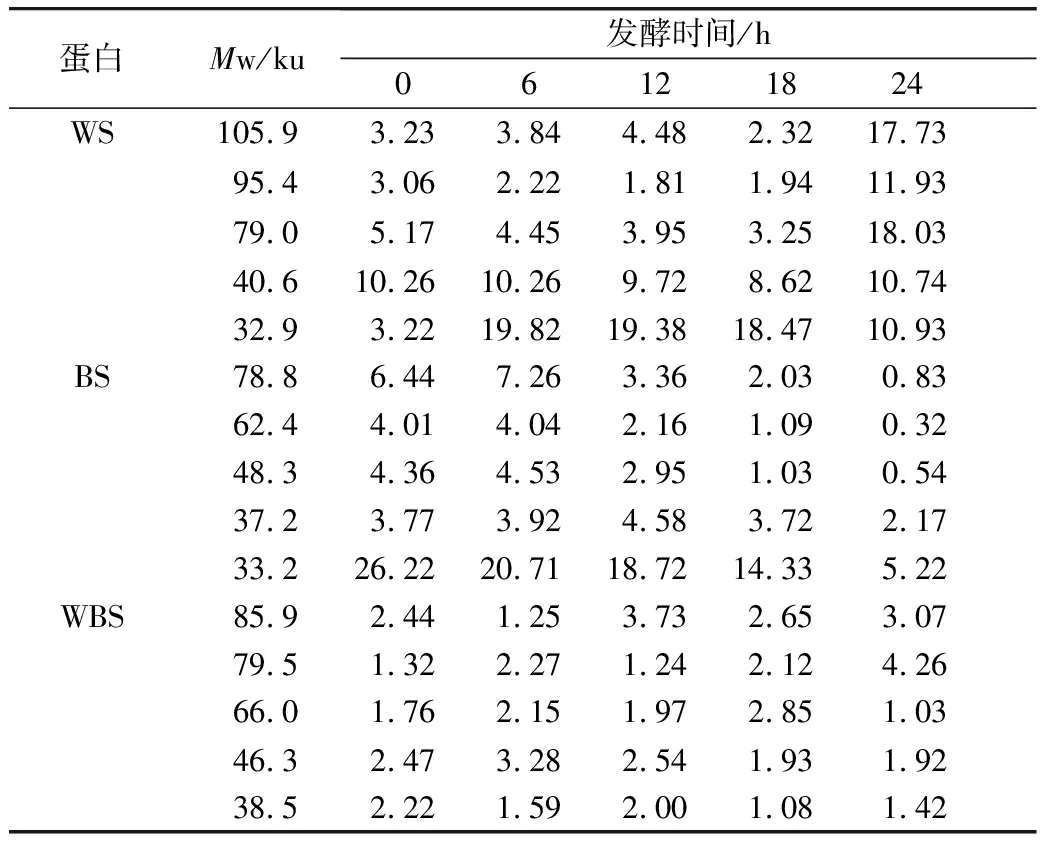
表4 植物乳桿菌ST-Ⅲ發酵WS過程中醇溶蛋白質量分數變化/%
2.7 發酵面團中游離氨基酸的變化
乳酸菌在發酵酸面團過程中能夠通過其自身的生長代謝和產有機酸等方式對面團中蛋白質進行降解,將大分子蛋白降解為小分子游離氨基酸,提高酸面團中蛋白質利用率和最終產品中游離氨基酸的含量,均衡產品營養價值[26]。植物乳桿菌ST-Ⅲ發酵酸面團過程中游離氨基酸含量變化見表5。不同發酵底物作用效果差別明顯。BS中的游離氨基酸含量隨著發酵的進行始終呈上升趨勢,發酵至18 h時,BS中的必需氨基酸含量較8 h時上升了22.22%,而非必需氨基酸含量則上升了16.92%;植物乳桿菌ST-Ⅲ發酵BS的前12 h游離氨基酸含量上升幅度較小,這可能是因為BS中的蛋白質首先被降解為小肽鏈,這些小肽鏈在12 h后被進一步地降解至游離氨基酸,同時苦蕎粉中的內源性蛋白酶和淀粉酶被激活,也利于促進游離氨基酸的代謝。而WS和WBS中的游離氨基酸含量均成先上升后下降的趨勢,且在植物乳桿菌ST-Ⅲ發酵18 h后酸面團中的總游離氨基酸含量均低于發酵初期,進一步說明植物乳桿菌ST-Ⅲ發酵苦蕎相對于小麥而言,更有利于蛋白質水解成氨基酸。
苦蕎蛋白富含一般谷物蛋白缺乏賴氨酸和精氨酸,其賴氨酸含量是大米、小麥的2倍以上[27]。隨著發酵時間的延長,BS中賴氨酸、精氨酸含量升高,此現象與預期相符,也印證了發酵可以將部分蛋白質進一步降解成具有營養價值的氨基酸。賴氨酸能夠增強人體的免疫力、并且能夠緩解緊張等的負面情緒,同時其能夠協同并促進營養素的吸收,使得這些營養素能夠更好的在人體內發揮其有益功效[28]。由此可以看出,植物乳桿菌ST-Ⅲ發酵對BS中的某些必需氨基酸和非必需氨基酸的增加有著明顯的促進作用,將由植物乳桿菌發酵的BS添加入烘焙類產品的制作當中,能夠有效地提升產品的營養價值。
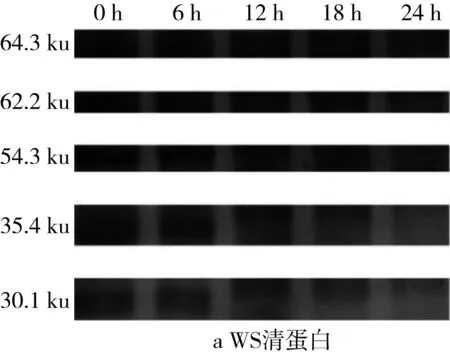
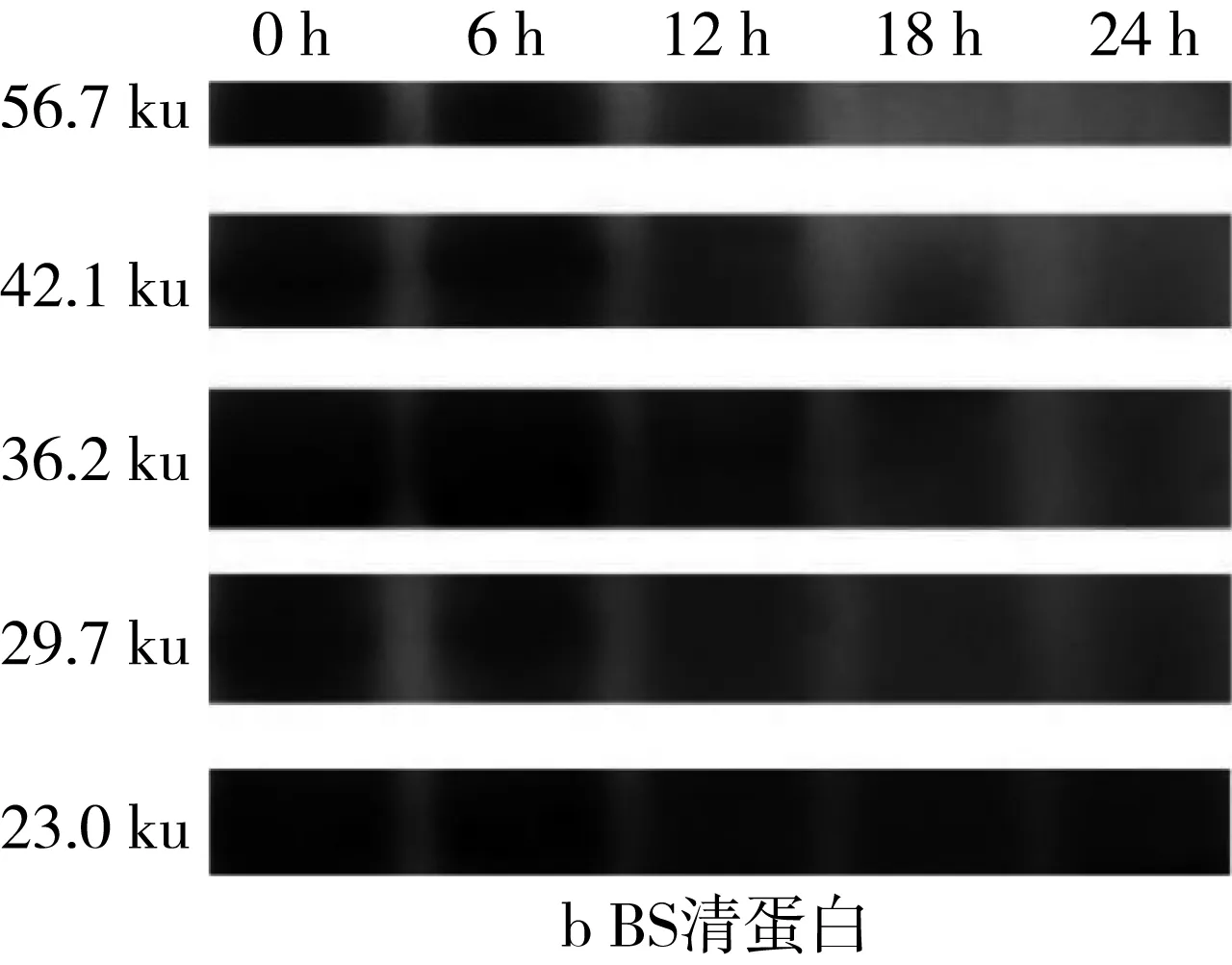
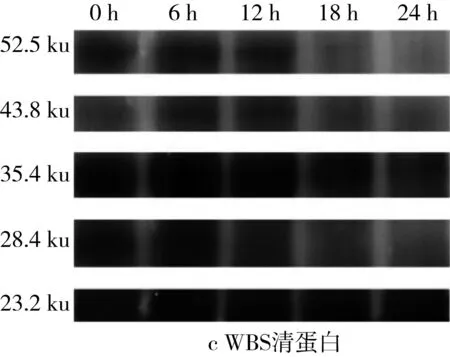
圖5 植物乳桿菌ST-Ⅲ發酵酸面團過程中清蛋白SDS-PAGE

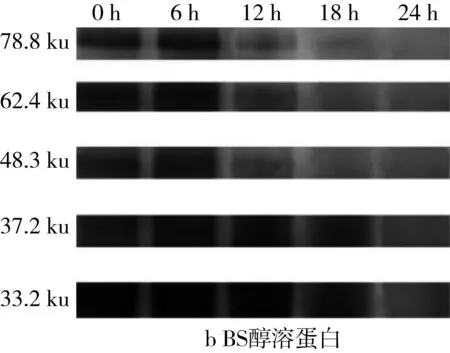
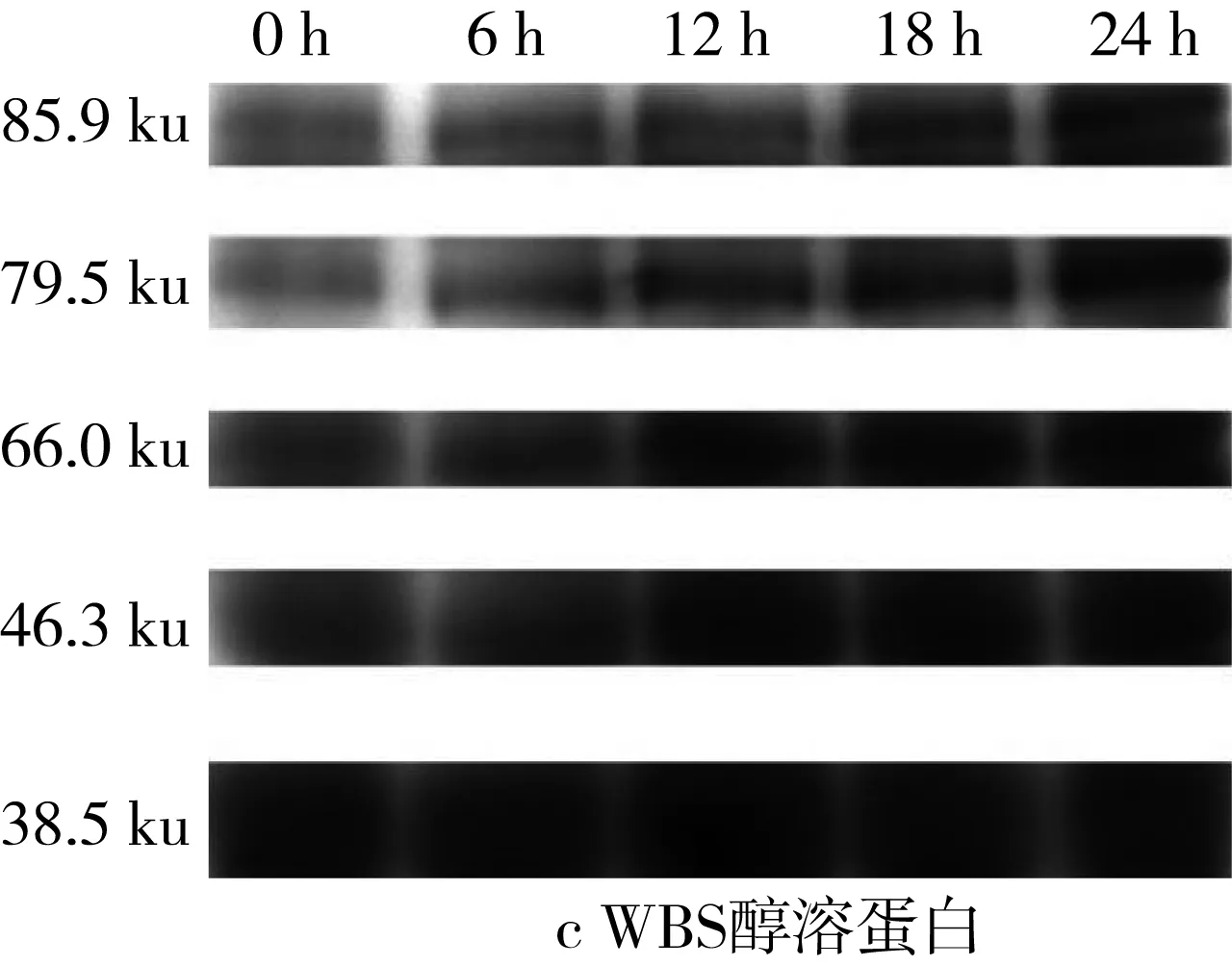
圖6 植物乳桿菌ST-III發酵3種酸面團過程中醇溶蛋白SDS-PAGE變化
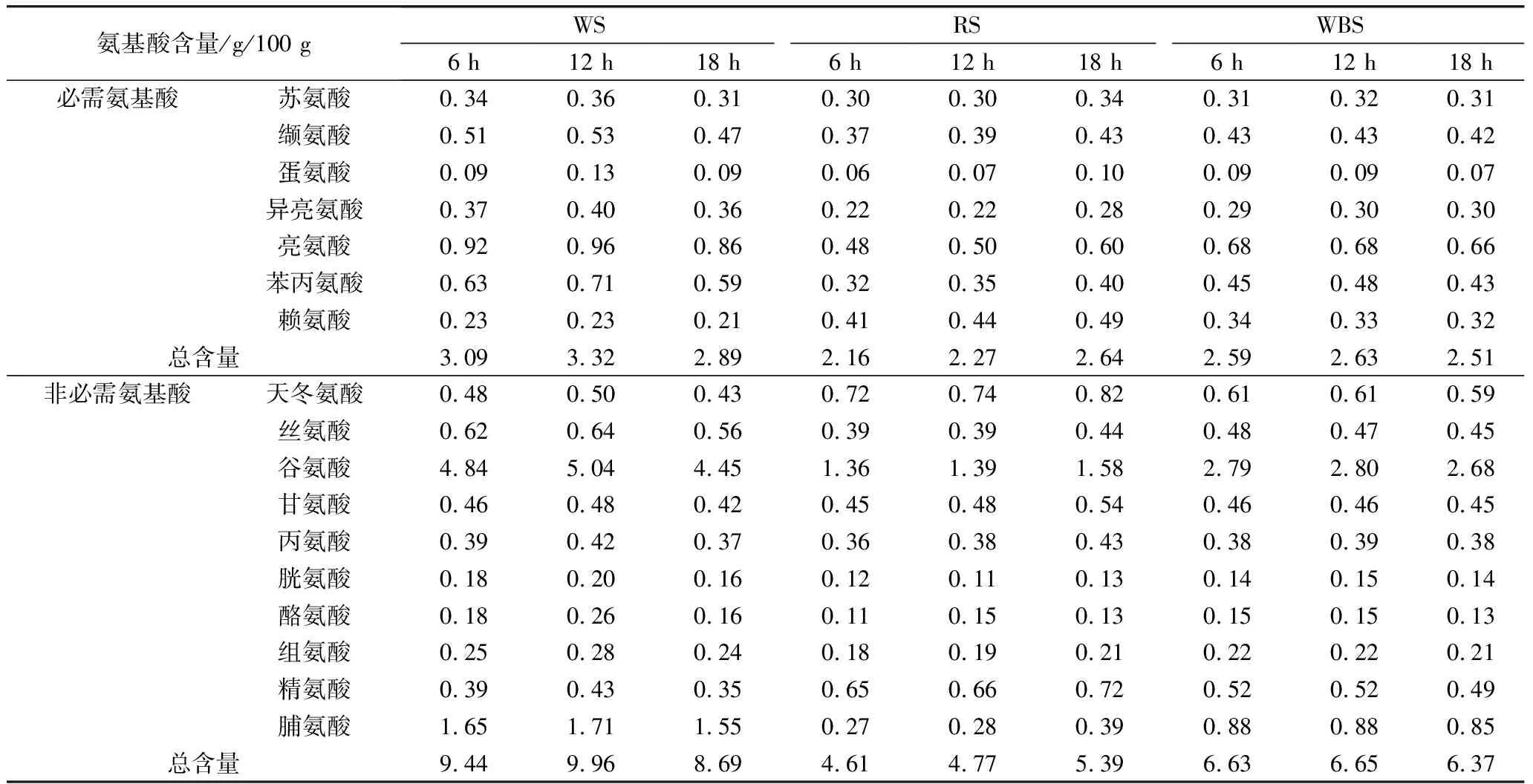
表5 植物乳桿菌ST-III發酵面團過程中游離氨基酸組成及含量變化
3 結論
將植物乳桿菌ST-Ⅲ添加到小麥、苦蕎及小麥-苦蕎混合粉中進行酸面團的制備,研究酸面團發酵過程中蛋白質的降解變化。結果表明,在發酵過程中,植物乳桿菌ST-Ⅲ在含有苦蕎基質的酸面團中生長較為旺盛,代謝產生大量乙酸和乳酸,特別是在發酵6~12 h時,BS的pH急劇下降,總酸度上升,且下降變化顯著快于WS、WBS,酸化能力較強。發酵12~18 h時,植物乳桿菌ST-III發酵對BS中苦蕎蛋白適度的降解效果最為明顯,且游離氨基酸含量也隨著發酵的進行呈上升趨勢。植物乳桿菌ST-Ⅲ在以苦蕎為基質的面團中生長良好,產酸及耐酸能力較強,利于促進蛋白質的降解。并且,利用植物乳桿菌ST-Ⅲ適度發酵苦蕎面團,能夠促進大分子量蛋白降解為小分子蛋白和多肽,且對BS中的某些必需氨基酸和非必需氨基酸的增加有著明顯的促進作用。因此,利用植物乳桿菌ST-Ⅲ發酵技術提高苦蕎面制品中的蛋白質利用率。
猜你喜歡
農產品加工(2023年14期)2023-08-17 10:32:28
青年歌聲(2018年5期)2018-10-29 03:18:40
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
廣東飼料(2016年3期)2016-12-01 03:43:12
家庭百事通·健康一點通(2016年4期)2016-04-22 09:16:57
林業與生態(2016年2期)2016-02-27 14:24:01
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
