CDKN2A 基因突變與腎細胞癌藥物敏感性和預后的關系
2022-09-26 08:35:28李松李瑞杰趙小磊
醫學信息 2022年16期
李松,李瑞杰,趙小磊
(1.河南大學淮河醫院泌尿外科,河南 開封 475000;2.鄭州市第二人民醫院綜合科,河南 鄭州 450000)
腎透明細胞癌(kidney renal clear cell carcinoma)是泌尿系統腎細胞癌的一個獨特亞型,根治性切除是腎細胞癌的主要治療方法,其次是化療和靶向治療。然而,即使手術治療成功,仍有30%的患者可能出現腫瘤復發或遠處轉移[1-3]。不同基因突變的患者可能對不同的藥物治療產生耐藥性,因此對不同基因突變的患者進行個體化治療更為重要[4]。因此,確定腎細胞癌進展和預后的潛在生物標志物和潛在相關分子機制是必要的,尋找潛在相關的靶向藥物對于個體化治療至關重要。Xu J等[5]研究證實,CDKN2A 啟動子甲基化發生在支氣管細胞癌變的早期。Hayashi T等[6]研究了胃癌患者的腫瘤組織,發現CDKN2A的失活通常是由5'CpG 島的純合缺失和高甲基化引起的,這在胃癌的發生發展中起著重要作用。Wang L等[7]發現,在77 例平滑肌肉瘤患者中,22%的患者存在CDKN2A 基因啟動子高甲基化,表明CDKN2A 啟動子高甲基化及其蛋白表達缺失與預后不良密切相關。然而,CDKN2A 在腎細胞癌中的作用尚不確定[8]。CDKN2A 突變狀態可能影響某些腎細胞癌患者的病情進展和治療。因此,探索CDKN2A 突變患者的一些相關信號通路將有助于進一步了解腎細胞癌進展的分子水平機制,并為患者的個體化治療策略提供指導。為了探索腎細胞癌預后和個體化治療反應的機制,本研究對腎細胞癌的RNA 測序數據集以分析基因突變,研究泛癌和腎細胞癌中CDKN2A 突變的藥物敏感性。
1 資料與方法
1.1 數據來源 從在線探索工具TCGA 數據庫下載(http://xena.ucsc.edu/)腎細胞癌基因表達譜(使用AgilentG4502A_07_3的腎細胞癌RNA 表達和突變基因)和臨床信息。采用開源網絡工具cBioPortal 分析Kaplan-Meier 圖和CDKN2A 突變(http://www.cbioportal.org/index.do)。BioPortal 數據庫(http://www.cbioportal.org)整合了各種基因數據類型,包括TCGA、UCSC、GDAC 等,從cBioPortal 數據庫獲得600份腎細胞癌樣本,通過數據庫篩選選項,提取患者相關資料,篩選條件:①“Cancer Type:KIRC”;②“Gene:CDKN2A”;③“Data Type:mRNA”。應用survival 選項分析600 份樣本中CDKN2A 突變情況與生存時間(總體生存和無疾病生存)關系圖,對3 種不同腎細胞癌研究中突變類型的mRNA 表達進行比較。
1.2 GDSC 數據庫分析 癌癥藥物敏感性基因組學數據庫(GDSC)(https://www.cancerrxgene.org/)是癌癥臨床研究中最常用的數據庫之一,收集了大量關于腫瘤細胞藥物敏感性的信息,進入數據庫所有藥物的界面,將“CDKN2A”數據集數據與化合物敏感性相關聯,查看泛癌和腎細胞癌中CDKN2A 突變在藥物的細胞系IC50圖,并繪制和生成與相應藥物敏感性的火山圖和散點圖。
1.3 基因集富集分析 基因集富集分析(GSEA)用于評估按表型相關性排序的基因列表中預定義基因集中的基因分布,以確定它們對表型的作用(http://software.org/gsea/下載.jsp)。應用分子特征數據庫(MSigDB),基于標稱P值和標準化富集分數(NES)分析路徑富集。根據CDKN2A 基因表達量分為兩組,通過GSEA4.1.0 分析CENPF 基因表達水平對各種生物通路基因集的影響,按照默認富集加權統計方法,選擇MSigDB 數據庫的基因集(c2.cp.kegg.v7.2.symbols.gmt)作為參照基因集,置換次數為1000 次,計算基因富集得分(enrichment score,ES)。
1.4 差異基因(DEGs)的鑒定與富集分析 使用R軟件包在微陣列數據中選擇-Log10(Pvalue)>2 且∣Log2(fold change)∣<1的DEGs。由DAVID(http://david.ncifcrf.gov)對DEGs 進行KEGG 途徑和GO 富集分析,其中FDR<0.05,-LogFDR>1.302的前幾個通路被認為是顯著的。
1.5 統計學方法 采用Wilcoxon 試驗比較突變型和野生型之間的CDKN2A mRNA 表達水平;采用Kaplan-Meier 法和Log rank 檢驗,通過Graphpad 計算不同CDKN2A 組之間的臨床結果;edgeR 和GSEA中的FDR 分別采用Benjamini-Hochberg 程序進行多次試驗調整,以控制FDR。P<0.05 被認為差異有統計學意義。
2 結果
2.1 腎細胞癌中的CDKN2A 表達及突變 600 例腎細胞癌樣本中有5.00%的CDKN2A 突變病例,其余為野生型(圖1A)。CDKN2A 在腎細胞癌中的突變類型包括亞基突變、錯義突變、截斷突變和跨越整個基因的深度缺失(圖1B)。其中,3.00%為深度缺失,2.00%為錯義突變、截斷突變和inframe 突變(圖2A)。CDKN2A 突變類型為深度缺失的mRNA 表達低于野生型(圖2B);與突變型相比,野生型的CDKN2A 表達較高(圖2C);CDKN2A 在腎細胞癌中的表達高于正常腎組織。

圖1 腎細胞癌中CDKN2A的突變
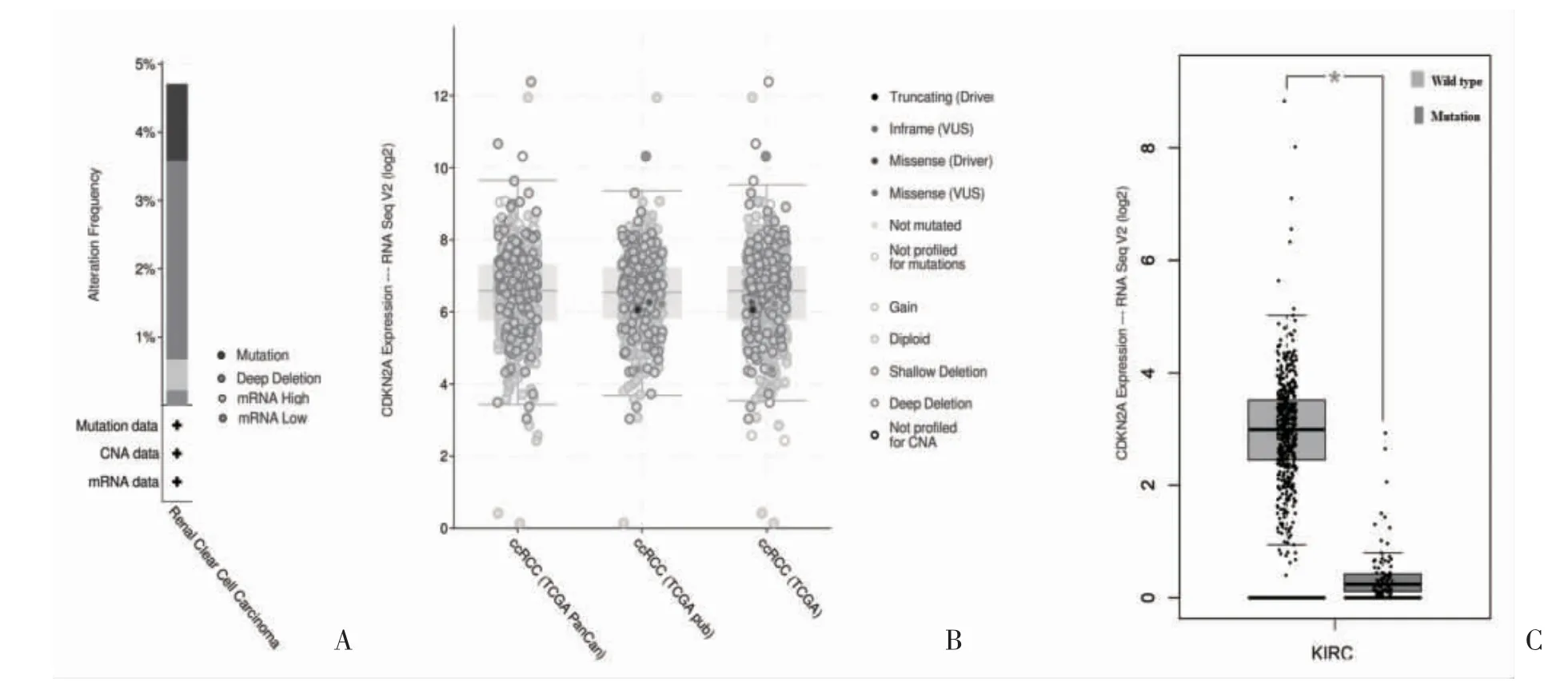
圖2 腎細胞癌中CDKN2A的突變表達
2.2 CDKN2A 基因突變對腎細胞癌患者預后和藥物敏感性的影響 腎細胞癌患者的臨床特征見表1,生存分析顯示,具有CDKN2A 突變的腎細胞癌患者總體生存率和無病生存率較差(圖3),在泛癌和腎細胞癌中,CDKN2A 突變型可能比野生型增加對Tenovin-6 等藥物的耐藥性,并且Tenovin-6 對各種癌癥和腎細胞癌中的野生型具有選擇性(圖4)。

圖3 CDKN2A 基因突變對腎細胞癌患者預后的影響

圖4 CDKN2A 基因突變對腎細胞癌患者藥物敏感性的影響

表1 腎細胞癌患者的臨床特征
2.3 基于GSEA的CDKN2A 突變信號通路分析GSEA 分析顯示,CDKN2A 突變信號通路主要富集在Wnt-catenin 信號、泛素介導的蛋白水解、緊密連接、TGF-β 信號、磷脂酰肌醇信號、erbb 信號和粘附連接(圖5)。2.4 DEGs的鑒定和富集分析 共有350 個基因被鑒定為DEGs,包括170 個下調基因和180 個上調基因。DEGs的火山圖見圖6A。使用DAVID 對DEGs進行富集分析,GO 分析結果表明,DEGs 主要富集于RNA 聚合酶Ⅱ啟動子、轉錄、細胞核、細胞質、轉錄因子復合物和序列特異性DNA 結合的轉錄負調控,見圖6B。KEGG 通路分析顯示,DEGs 主要富集于細胞周期、癌癥、慢性髓系白血病、癌癥中的microRNA 和p53 信號通路,見圖6C。

圖5 腎細胞癌CDKN2A 突變的GSEA 富集分析

圖6 DEGs的鑒定和富集分析
3 討論
CDKN2A 基因位于染色體9p21 上,受調控的CDKN2A 蛋白包含156 個氨基酸,在細胞周期中調節細胞增殖并抑制腫瘤的產生,該基因的失活可導致細胞分離控制和惡性進展為腫瘤[9]。目前研究發現,該基因在許多腫瘤中丟失或發生突變。CDKN2A基因失活的機制包括啟動子甲基化、純合缺失和點突變,平均頻率分別為33%、22%和15%;另外,研究還發現9p21 區域雜合性缺失與腫瘤發生有關[10]。
CDKN2A 在各種腫瘤的惡性轉化中突變類型包括純合缺失(HD)、雜合性缺失(LOH)、p16INK4a/p14ARF 基因啟動子的突變和異常甲基化[11]。Ibrahim I等[12]研究了肺癌、卵巢癌和頭頸部鱗狀細胞癌的89 個細胞系中246 個位點的純合缺失,發現CDKN2A 基因的純合缺失最常見(26%),且與疾病預后的關系不同。本研究發現,腎細胞中約5.00%的患者攜帶CDKN2A 突變,包括亞基突變、錯義突變、截斷突變和跨越整個基因的深度缺失,其中3.00%為深度缺失,2.00%為錯義突變、截斷突變和近端突變。Thompson ED等[13]發現,CDKN2A的總突變率為93.3%,啟動子異常甲基化占46.7%,純合缺失占36.7%,突變占13.3%。
Ibrahim IS等[14]發現,CDKN2A 啟動子甲基化與直腸癌患者的復發和預后相關,表明CDKN2A 啟動子甲基化可作為預測直腸癌預后的指標。Potjer TP等[15]發現,胰腺癌患者的腫瘤組織中存在一種特殊的CDKN2A 突變,表明CDKN2A 突變可能被用作胰腺癌的標志物。本研究顯示,CDKN2A 突變的腎細胞癌患者總體生存率和無病生存率較差,這表明CDKN2A 突變可能有助于腎細胞癌的進展。除此之外,降低CDKN2A 表達水平已被證明可增加腫瘤細胞對化療或放療的耐受性[16]。因此,CDKN2A 可以作為新的抗癌治療策略的靶點。本研究發現,與野生型相比,CDKN2A 突變對Tenovin-6 等藥物的敏感性較低,這表明Tenovin-6 突變在治療上的預后較差,并且在各種癌癥和腎細胞癌中,Tenovin-6 對野生型具有顯著的選擇性。隨著組蛋白去乙酰化酶(HDAC)的發展,HDAC 抑制劑(HDACi)已成為抗腫瘤藥物的研究熱點[17]。Tenovin-6 是近年來開發的一種HDACi,是脫乙酰酶SIRT1的抑制劑,選擇性地抑制Ⅲ類HDACs 中最常見的SIRT1 和SIRT2,并激活P53,具有更大的臨床應用價值。Tenovin-6 可以抑制腫瘤細胞的增殖,促進細胞凋亡和周期阻滯[18]。這些研究為腎細胞癌患者個體化應用特異性抗腫瘤藥物提供了額外證據,并為進一步研究提供了基礎。
本研究發現,腎細胞癌患者CDKN2A 突變信號通路主要富集Wnt-catenin 信號、泛素介導的蛋白水解、緊密連接、TGF-β 信號、磷脂酰肌醇信號、erbb信號和粘附連接。已有研究證實[12],CDKN2A 基因的負調控途徑是RB1 途徑。CDKN2A 蛋白包含四個連續的錨蛋白重復序列,它們與CDK4 和CDK6 結合,而CDK4 和CDK6 是一種雙周期依賴性蛋白激酶,作用于CDK4-6的非催化側,可抑制CDK 和細胞周期蛋白D1 復合物的催化活性[19]。CDKN2A 突變可能通過影響多種途徑促進腎細胞癌的進展,CDKN2A突變患者的腫瘤可能更容易進展和遠處轉移[20]。另外,本研究還發現,GO 分析中差異基因主要富集在RNA 聚合酶Ⅱ啟動子、轉錄的負調控中。
綜上所述,多個基因和通路途徑可能在CDKN2A 突變中起關鍵作用。本研究確定了與腎細胞癌中CDKN2A 突變相關的主要途徑,這可能通過制定腎細胞癌特定CDKN2A 突變亞型的治療策略來改善腎細胞癌的預后。另外,腎細胞癌中CDKN2A突變的機制和驗證仍需在臨床和分子生物學實驗中進一步研究。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
電子制作(2018年18期)2018-11-14 01:48:24
電子制作(2018年11期)2018-08-04 03:25:42
財經(2017年2期)2017-03-10 14:35:35
山東工業技術(2016年15期)2016-12-01 05:31:22
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
