不同冬小麥品種(系)拔節期低溫脅迫生理反應及抗寒性評價
2022-09-23 11:59:56曹燕燕葛昌斌齊雙麗廖平安李雷雷喬冀良張振永
江蘇農業科學 2022年17期
曹燕燕, 葛昌斌, 齊雙麗, 廖平安, 黃 杰, 王 君, 李雷雷, 喬冀良, 張振永
(漯河市農業科學院/河南省抗赤霉優質小麥種質創制及新品種培育工程技術研究中心,河南漯河 462300)
小麥是我國主要糧食作物,小麥的可持續發展關系到我國的糧食安全、人民溫飽和社會穩定。近幾年,隨著極端天氣頻繁出現,低溫凍害已成為影響小麥生長發育的重要環境因素之一。特別是春季低溫,小麥正處于拔節至孕穗期,此時小麥組織器官幼嫩、含水量大,抵抗霜凍的能力較弱;同時小麥穗分化正處于雌雄蕊分化至小花原基分化期,對低溫反應尤其敏感,如遇低溫很容易造成傷害。黃淮麥區是我國冬小麥主產區之一,目前低溫冷害已成為該區主要氣象災害之一,發生頻率高、面積大、危害重,嚴重制約小麥生長,因此對黃淮麥區的小麥品種進行抗寒性鑒定研究具有重大意義,不僅關系到今后小麥的育種和推廣問題,也直接關系到國家的糧食安全和社會穩定等問題。
小麥遭受低溫脅迫后,植株會產生一種應激反應起到保護作用,主要表現在小麥體內可溶性糖、可溶性蛋白、游離脯氨酸等滲透調節物質的含量明顯增加,細胞膜結構與成分改變以及一些新酶的產生等。于晶等研究表明,抗寒性強的小麥品種脯氨酸和可溶性蛋白含量均高于抗寒性弱的品種;劉艷陽等研究表明,超氧化物歧化酶(SOD)活性、丙二醛(MDA)含量是小麥抗寒性的2個抗寒指標;余海波研究發現,小麥拔節期遭受低溫脅迫后,對SOD和過氧化物酶(POD)活性、脯氨酸和可溶性蛋白質含量、可溶性糖含量、MDA含量等都有影響;衛秀英等的研究發現,抗寒性弱的小麥品種MDA含量高于抗寒性強的品種,而SOD活性和POD活性均低于抗寒性強的品種。在生產中,單一的生理指標很難準確反映各個小麥品種抗寒性的強弱,因此可結合隸屬函數法和產量指標等進行綜合評價。
漯麥906于2019年通過河南省審定,審定編號為豫審麥20190064;漯麥47于2021年通過湖北省審定,審定編號為鄂審麥20210007;漯麥36已完成試驗程序,即將國審。本研究以小麥新品種漯麥47、漯麥906、對照周麥18、參試新品系漯麥36、漯麥6098、漯麥956和漯麥C101共7個試驗材料為研究對象,室外盆栽種植,至小麥拔節期連盆一起移入春化室模擬低溫處理,探索拔節期低溫對不同品種(系)小麥葉片生理生化特性的影響,并運用隸屬函數對各品種(系)抗寒性進行分析,結合產量表現對供試材料的抗寒性做出綜合評價,以期為小麥品種漯麥906和漯麥47,將審定品系漯麥36做出抗寒性評價以及后期示范推廣和育種親本的選擇等提供技術支撐。
1 材料與方法
1.1 試驗材料與設計
試驗于2020—2021年在河南省漯河市農業科學院試驗田網室進行,采用盆栽方法。供試材料為漯麥47、漯麥906、周麥18、漯麥36、漯麥6098、漯麥956和漯麥C101共7個品種(系)小麥,種子由漯河市農業科學院小麥研究所提供。盆栽用土為大田 0~30 cm耕層土,有機質含量為14.13 g/kg,全氮含量為0.86 g/kg,硝態氮含量為25.56 mg/kg,速效磷含量為42.59 mg/kg,速效鉀含量為168.99 mg/kg,pH值為7.47。盆直徑30 cm,高60 cm,底部用網袋包裹,埋入大田,盆內土壤表面與地面持平。播前每盆施復合基肥6 g,土壤水澆透沉實后于2020年10月15日播種。3葉期定苗,每盆留苗9株,每個品種種植9盆,其中3盆用于低溫取樣,3盆用于低溫脅迫后測產,3盆用于常溫對照,共計63盆。澆水量根據土壤墑情決定,每盆澆水量一致。其他管理同大田,2021年6月2日收獲。
1.2 低溫處理方法
2021年春季,各小麥品種生長至拔節期,通過顯微鏡觀察幼穂分化,各品種進入雌雄蕊原基分化期。將各品種6盆從大田移入春化室模擬春季低溫脅迫,春化室內溫度設置為白天0 ℃、夜間-4 ℃,溫度誤差為0.5 ℃,空氣相對濕度為70%。采用人工光源光照,光照時間為08:00—18:00。低溫處理0、1、2、3、4 d時取主莖最上部的展開葉用于生理指標檢測,每個品種取3盆,3個重復。各個品種剩余3盆在低溫處理后移至自然條件下,直至收獲,用于產量性狀方面的調查。
1.3 測定項目及方法
生理生化性狀的測定:每個供試材料測定3次取其平均值。采用硫代巴比妥酸比色法測定MDA含量;采用氮藍四唑(NBT)法測定SOD活性;采用愈創木酚法測定POD活性;采用過氧化氫法測定過氧化氫酶(CAT)活性;采用考馬斯亮藍G-250比色法測定可溶性蛋白質含量;采用酸性茚三酮法測定脯氨酸含量。
產量因子的測定:低溫脅迫處理4 d后,將未取樣的盆栽小麥放置到大田原位置,自然條件下自然恢復生長。蠟熟期后收獲,分別記載單株穗數、單株穗粒數和單株產量。
1.4 抗寒性評價及方法
應用隸屬函數法評價抗寒性,計算公式如下:
(1)與抗寒性呈正相關的各指標(SOD活性、POD活性、可溶性蛋白含量和脯氨酸含量等)用公式()=(-)/(-)計算。
(2)與抗寒性呈負相關的各指標(MDA含量) 用公式()=1-(-)/(-)計算。式中:()為第個品種、第天相對百分數的隸屬度;表示第個品種、第天某一指標測定值與其對應對照值的百分數;、為所有供試品種某一具體指標在某一天最大值和最小值的相對百分數值。對各個指標所求的隸屬函數值進行累加、求取平均值。之后按照從大到小的順序對各個品種的隸屬函數平均值進行排序,對應的得到各個品種抗寒性的強弱。
1.5 數據處理
利用WPS Office進行數據統計和作圖,利用DPS 15.10的Duncan’s新復極差法進行差異顯著性分析。
2 結果與分析
2.1 低溫脅迫對拔節期小麥葉片SOD活性的影響
如圖1所示,拔節期持續低溫脅迫后,各小麥品種(系)SOD活性出現不同程度增加,以增強對逆境的適應能力,與常溫對照相比,均差異顯著。隨著低溫持續時間的延長,SOD活性整體表現為先升后降的趨勢。低溫處理1 d后,品種(系)間升高幅度存在明顯差異,漯麥906升幅最大(227.42%),漯麥C101升幅最小(101.09%),其余品種(系)升幅介于兩者之間;低溫處理2 d,除周麥18和漯麥C101外,其他品種(系)SOD活性出現下降;隨著低溫處理時間延長至3 d和4 d,大部分品種(系)SOD活性表現先上升后下降。由試驗可知,在持續低溫脅迫下,不同小麥品種(系)SOD活性存在差異,但都有較大幅度提高,以抵御低溫脅迫對植株造成的傷害。

2.2 低溫脅迫對拔節期小麥葉片POD活性的影響
如圖2所示,低溫脅迫后各品種(系)的POD活性均有升高,且升高程度不同,也都與對照差異顯著(除漯麥36處理2 d與對照差異不顯著外)。低溫脅迫1 d后,各品種(系)之間以漯麥47升幅最高(111.29%),其次為漯麥6098(109.74%);低溫脅迫2 d,除漯麥956活性繼續增加外,其余6個品種(系)POD活性均下降;低溫脅迫3 d和4 d,各品種(系)POD活性持續上升,與對照相比,增幅為68.33%~167.19%,其中漯麥47和漯麥6098增幅最大,分別為167.19%和115.24%。說明在長期的低溫脅迫下,漯麥47和漯麥6098的POD的抗氧化能力仍持續較高,而其他5個品種(品系)的抗氧化能力稍弱,但對活性氧自由基仍有一定的清除作用。
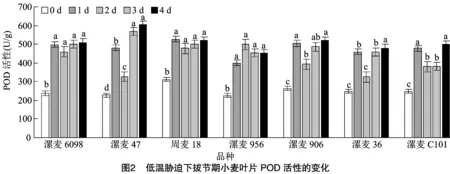
2.3 低溫脅迫對拔節期小麥葉片CAT活性的影響
如圖3所示,低溫脅迫后各品種(系)的CAT活性均有增加,也都與對照差異顯著。低溫脅迫處理1 d后,7個小麥葉片中的CAT活性平均升高328.74%,但品種(系)間升高幅度存在明顯差異;除漯麥956和漯麥C101外,隨著低溫脅迫時間的延長,各品種(系)CAT活性呈現“升高—降低—升高—升高”的趨勢;低溫處理 4 d 后,與對照相比,漯麥47增幅最高,為444.89%,漯麥C101增幅最少,為203.37%,其他材料介于兩者之間。這表明持續低溫脅迫后,各供試材料清除活性氧的能力有差異,但在低溫脅迫4 d后,除漯麥906和漯麥C101外,其他品種(系)CAT活性仍較高,可以將低溫對植物的傷害控制在一定范圍內。
2.4 低溫脅迫對拔節期小麥葉片丙二醛活性的影響
如圖4所示,低溫脅迫后各品種(系)的MDA含量均有增加,也都與對照差異顯著。低溫脅迫 2 d,漯麥47和漯麥C101的MDA含量下降,其他5個材料MDA含量略有上升;與處理3 d相比,低溫脅迫4 d后漯麥6098、漯麥47、周麥18和漯麥906的MDA含量出現下降,其他3個材料MDA含量則繼續上升。這說明低溫加速了各品種(系)膜脂過氧化,活性氧已傷害到各品種(系),但根據MDA含量的不同,各品種(系)間受傷害的程度存在差異。

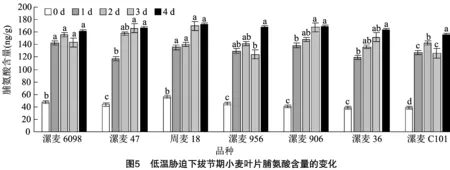
2.5 低溫脅迫對拔節期小麥葉片脯氨酸含量的影響
由圖5可以看出,低溫脅迫后各品種(系)的脯氨酸含量均有增加,也都與對照差異顯著。隨著低溫脅迫時間延長,漯麥47、周麥18、漯麥906和漯麥36的脯氨酸含量一直呈現上升趨勢;與脅迫1 d相比,漯麥6098、 漯麥956和漯麥C101的脯氨酸含量呈現先上升后降低再上升的趨勢。由此可以看出,低溫脅迫下,各品種(系)會積累較多的脯氨酸,以維護細胞膜內代謝的平衡和細胞膜結構的穩定性,以減輕低溫引起的傷害,但各品種(系)間脯氨酸含量變化趨勢存在差異。
2.6 低溫脅迫對拔節期小麥葉片可溶性蛋白質含量的影響
如圖6所示,各供試品種(系)在拔節期遭受低溫脅迫后,可溶性蛋白質含量均有不同程度增加,除漯麥36外,其他6個均與對照呈顯著性差異。低溫脅迫1 d后,各品種(系)之間以漯麥6098升幅最高(100.07%),周麥18升幅最低(43.98%),其他供試材料介于兩者之間;與低溫脅迫2 d相比,低溫脅迫 3 d,漯麥6098、周麥18和漯麥906可溶性蛋白質含量出現下降,其他4個持續上升;與低溫脅迫3 d相比,低溫持續脅迫4 d,漯麥6098、周麥18和漯麥C101可溶性蛋白質含量出現上升,其他4個品種(系)出現下降,漯麥6098表現顯著差異。由此可知,低溫脅迫時間越長,不同品種(系)間可溶性蛋白含量差異越大。

2.7 拔節期低溫脅迫對小麥籽粒產量構成因素的影響
由表1可知,與CK相比,小麥在拔節期受到低溫脅迫4 d后,各供試材料不孕小穗數顯著升高,單株分蘗數、穗粒數、結實小穗數、千粒質量和單穗產量均減少,不孕小穗數增加。其中漯麥47單株減產率最低,為39.57%,漯麥956減產率最高,達到75.73%。綜合產量各因素變化數據,在本試驗環境下,拔節期低溫脅迫對小麥不孕小穗數和單穗產量影響較大,而對結實小穗數和穗粒數影響比較小。
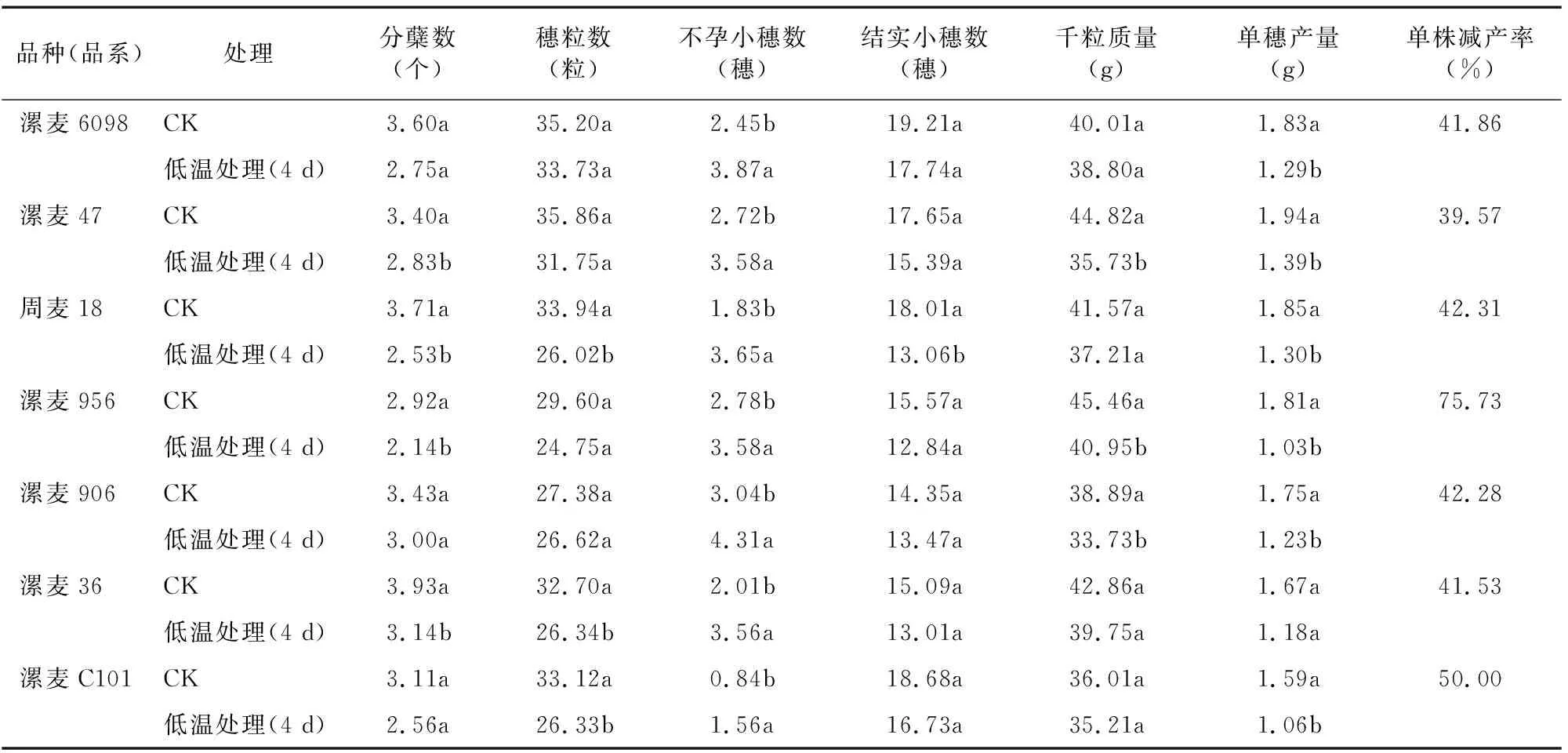
表1 拔節期低溫脅迫對單株小麥產量的影響
2.8 小麥品種抗寒性的隸屬函數值綜合分析
從表2可知,在低溫脅迫下,品種間隸屬函數值差異非常明顯,同一品種(系)不同指標的隸屬函數值也存在差異。每一品種(系)各項測定指標的隸屬值的平均綜合值越高,表明該品種(系)的抗寒性越強。根據綜合值,將7個供試品種(系)可分為3類:第Ⅰ類為漯麥6098、漯麥47、周麥18和漯麥906,屬于抗寒性強的品種(系);第Ⅱ類為漯麥36和漯麥C101,屬抗寒性較好的品系;第Ⅲ類為漯麥956,屬抗寒性弱的品系。試驗結果與各品種(系)的單株減產率基本一致,且與田間各品種(系)的抗寒性表現基本一致。

表2 低溫脅迫下小麥葉片各生理指標的隸屬值
3 討論與結論
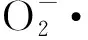
持續低溫脅迫下,當保護酶的清除能力不能及時清除多余的氧自由基時,就會引起膜脂不飽和脂肪酸發生過氧化,產生膜脂過氧化產物MDA,膜脂過氧化的程度隨溫度降低加重,同時與低溫脅迫時間呈正相關。本試驗中,低溫脅迫1 d后,各供試材料的MDA含量相比于對照,均顯著上升,表明低溫脅迫后,各供試材料抗氧化能力雖有所增強,但仍不能及時清除活性氧,引起膜脂過氧化,對小麥造成傷害;隨著低溫脅迫的持續,各供試材料MDA含量變化有升有降,說明各個品種(系)對低溫的適應調節能力存在差異;低溫脅迫4 d與脅迫 3 d 相比,抗寒性強的4個品種(系)漯麥6098、漯麥47、周麥18和漯麥906,MDA含量出現下降,說明抗寒性強的品種,在持續低溫脅迫后,能夠通過自身調節使質膜透性部分修復。
脯氨酸通過其亞氨基促進蛋白質水合而成為重要的滲透調節物質之一,可溶性蛋白的增多有利于維持低溫狀態時酶的構象,因此脯氨酸是植物抗寒生理中的一個重要指標。本試驗中,在低溫脅迫1 d后,各供試品種(系)脯氨酸含量均有不同程度增加,均與對照呈顯著性差異。隨著低溫脅迫時間延長,漯麥47、周麥18、漯麥906和漯麥36的脯氨酸含量一直呈現上升趨勢;與脅迫處理1 d相比,漯麥6098、漯麥956和漯麥C101的脯氨酸含量呈現先上升后降低再上升的趨勢。說明漯麥47、周麥18、漯麥906和漯麥36這4個品種(系)在持續低溫脅迫下可以通過積累較多的脯氨酸來提高自身抗寒性。
可溶性蛋白作為滲透調節物質廣泛存在于植物細胞內,作用主要是為了調節細胞內滲透壓的平衡、維持細胞正常的生理代謝活動。研究表明,植株細胞內可溶性蛋白含量的高低與其自身抗寒性存在密切的關系。低溫脅迫后植株體內可溶性蛋白含量升高,以提高植物在低溫下的滲透調節能力和代謝功能,進一步提高植株的抗寒能力。本試驗中低溫持續脅迫4 d后,漯麥6098、周麥18和漯麥C101可溶性蛋白質含量出現上升,與脅迫處理3 d相比,漯麥6098表現顯著性差異。說明這3個供試材料在長期低溫脅迫下,通過提高體內可溶性蛋白的含量來增強自身的抗寒能力,漯麥6098表現更突出。隨著低溫脅迫時間的延長,其他試驗材料可溶性蛋白含量存在差異。
拔節期低溫凍害對小麥穗部發育和單株產量影響較大,不孕小穗數顯著提高,穗粒數減少,千粒質量降低,且各材料間單穗產量下降幅度不同。本試驗中漯麥47單株減產率最低,為39.57%,漯麥956減產率最高,達到75.73%。綜合各產量因素變化數據得出,拔節期低溫脅迫對小麥單株產量和不孕小穗數影響較大,而對結實小穗數和穗粒數影響相對較小。
通過隸屬函數綜合值分析評價小麥拔節期的抗寒性,供試材料被分為3類。漯麥6098、漯麥47、周麥18和漯麥906,屬于抗寒性強的品種(系);漯麥36和漯麥C101,屬抗寒性較好品系;漯麥956,屬抗寒性弱的品系。這是檢測各材料幾個生理生化指標綜合反應的結果,能夠較準確地反映出各品種(系)間抗寒性的差異,再結合各材料的產量指標,來綜合評價各供試材料的抗寒性,更加準確。
拔節期是小麥營養生長的重要時期,也是對低溫敏感的時期。低溫脅迫下,植物細胞多種生理生化指標發生改變以適應低溫條件和進行自我調節。小麥應對低溫逆境脅迫涉及一系列復雜因素,既有外部因素,也有內部因素。本研究根據試驗需要,篩選了7個供試材料,研究了拔節期持續低溫脅迫下生理生化指標和產量的變化,為收集和鑒定抗寒性小麥種質資源提供依據。目前分子生物學技術發展迅猛,植物耐受低溫脅迫機制的研究也從生理生化水平向分子水平轉變,下一步將開展相關抗凍基因表達等方面的研究,將分子育種手段有效融合到常規育種中,為培育抗寒、耐倒春寒的小麥新品種提供技術支撐。
