水稻OsAKT2的鉀吸收功能及其在地上部分K+回流中的潛在作用
2022-09-22 08:23:56黃亞楠李俊林蘇彥華
植物營養(yǎng)與肥料學(xué)報 2022年7期
黃亞楠,李俊林,蘇彥華
(1 中國科學(xué)院南京土壤研究所/土壤與農(nóng)業(yè)可持續(xù)發(fā)展國家重點(diǎn)實(shí)驗(yàn)室,江蘇南京 210008;2 中國科學(xué)院大學(xué),北京 100049;3 山東省蠶業(yè)研究所,山東煙臺 264002)
K+是植物生長發(fā)育所必需的三大礦質(zhì)營養(yǎng)元素之一,也是植物體中含量最豐富的無機(jī)陽離子,可占植物干重的10%。K+在調(diào)節(jié)酶活性、蛋白質(zhì)合成、細(xì)胞滲透勢、細(xì)胞內(nèi)電荷平衡以及提高植物對環(huán)境脅迫的適應(yīng)能力等方面有重要作用[1–2]。土壤溶液中的K+濃度在0.01~20 mmol/L波動,而細(xì)胞質(zhì)中的K+濃度維持在50~100 mmol/L范圍內(nèi)且相對穩(wěn)定[3]。植物中有大量負(fù)責(zé)K+吸收和轉(zhuǎn)運(yùn)的功能部件,包括高親和的鉀轉(zhuǎn)運(yùn)體(在鉀濃度<0.2 mmol/L下起作用)和低親和的鉀離子通道(在鉀濃度>1 mmol/L下起作用),它們決定了植物體內(nèi)諸多功能和調(diào)控特性各異的鉀吸收系統(tǒng)[4]。一般鉀離子通道吸收的K+約占植物總吸鉀量的50%[5],其中Shaker型K+通道被認(rèn)為是介導(dǎo)植物鉀離子吸收、轉(zhuǎn)運(yùn)和細(xì)胞內(nèi)動態(tài)平衡最為重要的一類鉀通道[6]。
土壤溶液的K+經(jīng)植物根表皮和皮層細(xì)胞中的鉀離子通道吸收,再通過根中柱鞘和木質(zhì)部薄壁細(xì)胞中的鉀離子通道將K+運(yùn)輸?shù)街参锏厣喜俊H~片保衛(wèi)細(xì)胞作為K+發(fā)揮功能的終端,其中的鉀離子通道通過調(diào)節(jié)K+的吸收和外排來調(diào)控氣孔開關(guān)[4–7]。在植物的生長發(fā)育過程中,K+具有很強(qiáng)的移動性,葉片中多余的K+可沿韌皮部向下回流到根系中,衰老葉片組織中的K+也可重新分配到幼嫩及生長發(fā)育較旺盛的組織。植物體內(nèi)的這種鉀循環(huán)可表征地上部鉀營養(yǎng)狀況并調(diào)節(jié)根系對K+的吸收[8]。耐低鉀型大麥能夠高效地將K+轉(zhuǎn)運(yùn)至功能葉,從而使新生葉保持正常的光合作用和代謝活動[9]。煙草葉片的鉀含量是表征煙葉品質(zhì)的重要指標(biāo),由老葉向新葉的K+回流會導(dǎo)致成熟期煙草葉片中K+的流失,從而影響煙葉品質(zhì)[10]。葉片中K+的分布及含量都會對植物的生理功能產(chǎn)生一定的影響。作為重要的滲透調(diào)節(jié)物質(zhì),K+參與調(diào)控氣孔開關(guān)過程。保持葉片較高的K+濃度使得保衛(wèi)細(xì)胞可以吸收更多的水分,促進(jìn)氣孔的開放[11]。另外K+會影響光合作用所需要酶類的活性、ATP合成、葉綠素含量等。缺K+通常會導(dǎo)致植物光合作用效率下降,并影響光合作用產(chǎn)物的轉(zhuǎn)運(yùn)[12–13]。但迄今為止,植物地上部K+回流的結(jié)構(gòu)基礎(chǔ)和分子機(jī)制還有待進(jìn)一步深入探索。
鉀離子通道AKT2主要在韌皮部表達(dá),兼具K+吸收和外排活性,但吸收活性占主導(dǎo)地位。韌皮部AKT2介導(dǎo)自上而下的K+運(yùn)輸,與植物地上部分K+的回流途徑相吻合;而其K+外排活性主要體現(xiàn)在對韌皮部細(xì)胞極化狀態(tài)(由H+內(nèi)流引起)的平衡方面,進(jìn)而協(xié)助蔗糖/H+共轉(zhuǎn)運(yùn)體將地上部光合同化產(chǎn)物蔗糖經(jīng)由韌皮部的向下運(yùn)輸過程[14–16]。擬南芥韌皮部AKT2缺失會降低細(xì)胞內(nèi)鉀離子和蔗糖含量以及膜電位對K+的依賴性,導(dǎo)致植株生長減緩、發(fā)育延遲[14–16]。AKT2的表達(dá)豐度受到光照和光合同化產(chǎn)物的誘導(dǎo),這也暗示其可能參與韌皮部蔗糖的運(yùn)輸[15]。脫落酸處理會增加AKT2的表達(dá)豐度,提示AKT2在植物應(yīng)對干旱脅迫時有重要作用[14]。在鹽脅迫條件下,水稻OsAKT2的缺失會使得植株韌皮部的K+降低,導(dǎo)致自上而下的K+再分配過程受損。此外,OsAKT2還調(diào)節(jié)K+和蔗糖從老葉到新葉中的轉(zhuǎn)運(yùn),并影響籽粒形成和產(chǎn)量[17]。
通過歸納分析本領(lǐng)域最新研究進(jìn)展及本課題組前期工作,發(fā)現(xiàn)水稻OsAKT2主要定位于地上部韌皮部細(xì)胞[18],且主要介導(dǎo)K+的吸收,推測其可能在地上部K+回流和再分配過程中發(fā)揮潛在作用。為此,通過蛙卵電生理技術(shù)研究了OsAKT2介導(dǎo)K+運(yùn)輸?shù)墓δ芴卣鳎ㄆ潆娏鞅旧硖匦浴⑩洕舛纫蕾囆浴㈦x子選擇性及對鉀通道抑制劑的響應(yīng)情況。進(jìn)一步模擬田間生長環(huán)境,利用實(shí)時熒光定量PCR試驗(yàn)測定OsAKT2基因表達(dá)的晝夜節(jié)律特征及對外界脅迫條件的響應(yīng)規(guī)律。
1 材料與方法
1.1 質(zhì)粒構(gòu)建
將水稻OsAKT2(GenBank: JN989970.1)基因經(jīng)酶切位點(diǎn)SmaI/NotI構(gòu)建到pCI載體上,用質(zhì)粒小提試劑盒(Axygen,AP-MN-P-250)提取質(zhì)粒,并濃縮至 1 μg/μL 待用。
1.2 蛙卵注射與電生理檢測
非洲爪蟾(Xenopuslaevis)冰浴麻醉1 h后,用手術(shù)刀在其腹部切開小口取出蛙卵,置于1 mg/mL的膠原酶A中消解1~2 h,挑選出健康飽滿的蛙卵放入 ND96 溶液 (96 mmol/L NaCl、1.8 mmol/L CaCl2、2 mmol/L KCl、1 mmol/L MgCl2、5 mmol/L HEPES,pH7.4)中培養(yǎng)。通過微量注射儀(Nanol 2000,WPI,Sarasota,F(xiàn)L,USA),將 59.8 nL 的 pCIOsAKT2質(zhì)粒cDNA注射到蛙卵中,等量的無菌水注射到蛙卵中作為陰性對照。在相應(yīng)的處理?xiàng)l件下,用“注射OsAKT2的蛙卵產(chǎn)生的電流”減去“注射H2O的蛙卵產(chǎn)生的電流”得到“OsAKT2產(chǎn)生的凈電流”,以扣除各離子處理對蛙卵本身的影響。注射后的蛙卵置于19℃的ND96溶液中培養(yǎng)(含50 mg/L慶大霉素),每天更換一次培養(yǎng)液,并及時去除壞死細(xì)胞。2~3天后用雙電極電壓鉗放大器(Axoclamp 900A, Foster City, CA, USA)檢測蛙卵細(xì)胞電流。在檢測電流時,鉗制電壓為-40 mV,階越電壓為 10 mV,施加的電壓范圍從-160 mV 到+50 mV,刺激間隔為 2 s。基本檢測浴液含有 1.8 mmol/L CaCl2、1 mmol/L MgCl2、5 mmol/L HEPES-NaOH,pH 調(diào)整為7.4,KCl和NaCl濃度根據(jù)實(shí)驗(yàn)設(shè)置進(jìn)行調(diào)整(用于調(diào)整溶液中的恒定離子強(qiáng)度)。用pClampfit 10.3(Molecular Devices)初步分析電生理數(shù)據(jù)。
檢測所用處理溶液如下:
OsAKT2的鉀吸收特征檢測: 0、1、2、5、10、20、50、100 KCl (mmol/L)。
OsAKT2 的離子選擇性檢測: 100 LiCl、100 NaCl、100 RbCl、100 NH4Cl、100 KCl (mmol/L)。
通道抑制劑對OsAKT2的影響檢測: 1 BaCl2、1 CsCl、25 TEACl、50 KCl (mmol/L)。
1.3 水稻生長試驗(yàn)
以粳稻日本晴 (Oryzasativa.ssp.JaponicNipponbare)為供試水稻材料。水稻幼苗在改良的IRRI溶液中生長,光周期為16 h光照/8 h黑暗,溫度設(shè)定為27℃/25℃ (光照/黑暗),光強(qiáng)度為 400 μmol/(m2·s),相對濕度設(shè)定為70%。將10天苗齡(從水稻發(fā)芽后算起)的水稻幼苗移至不同的處理液中:0 mmol/L K+、1 mmol/L K+、20 mmol/L K+、0 mmol/L NH4+、1 mmol/L NH4+、10 mmol/L NH4+、180 mmol/L sorbitol、15% PEG-6000,分別處理 0、4、8、12、20 和 24 h后收獲植株,用于RNA提取。
其中改良的IRRI營養(yǎng)液配方為:0.5 mmol/L(NH4)2SO4、0.3 mmol/L KH2PO4、0.35 mmol/L K2SO4、1 mmol/L CaCl2·2H2O、1 mmol/L MgSO4·7H2O、0.5 mmol/L Na2SiO3、20 μmol/L NaFeEDTA、20 μmol/L H3BO3、0.32 μmol/L CuSO4·5H2O、9 μmol/L MnCl2·4H2O、0.77 μmol/L ZnSO4·7H2O、0.39 μmol/L Na2MoO4·2H2O、pH調(diào)至5.8。不同的銨處理濃度采用(NH4)2SO4調(diào)節(jié),其他離子成分保持不變;不同的鉀處理濃度使用KCl調(diào)節(jié),并分別用NaH2PO4和Na2SO4取代KH2PO4和K2SO4,其他離子成分保持不變。
1.4 水稻地上部分OsAKT2基因表達(dá)豐度的測定
使用總RNA提取試劑(R401-01,南京諾唯贊生物科技有限公司)對收獲后的水稻地上部樣品進(jìn)行總RNA提取。用反轉(zhuǎn)錄試劑HiScriptⅢRT-SuperMix for qPCR (+gDNA-wipper,R323-01,南京諾唯贊生物科技有限公司)將1 μg總RNA反轉(zhuǎn)錄成cDNA。實(shí)時熒光定量PCR酶為ChamQ Universal SYBR qPCR Master Mix (Q711-02,南京諾唯贊生物科技有限公司)。引物由南京金斯瑞生物科技有限公司合成。使用熒光定量PCR儀LightCycler 480 (Roche,Switzerland)檢測基因表達(dá)情況,反應(yīng)條件為:95℃(30 s),95℃ (10 s)、60℃ (15 s)、72℃ (15 s),44 個循環(huán)。以水稻的看家基因OsActin作為內(nèi)標(biāo),用2–ΔΔCT方法分析目的基因的相對表達(dá)量。
1.5 Shaker鉀通道的系統(tǒng)進(jìn)化樹分析與關(guān)鍵位點(diǎn)序列比對
鉀通道序列由NCBI (http://www.ncbi.nlm.nih.gov/)網(wǎng)站下載獲得,利用DNAMAN對這些鉀通道序列進(jìn)行比對分析,并構(gòu)建Shaker鉀通道系統(tǒng)發(fā)育樹。
1.6 數(shù)據(jù)處理與分析
使用SPSS 16.0進(jìn)行統(tǒng)計分析,采用SigmaPlot 12.5繪圖,所示數(shù)據(jù)為平均值±標(biāo)準(zhǔn)誤SE (n>3)。多重比較采用Duncan法,差異顯著水平為5%。
2 結(jié)果與分析
2.1 OsAKT2的氨基酸序列分析
由圖1a可看出,OsAKT2在通道孔區(qū)域具有特征性的序列TxxTxGYGD,屬于典型的Shaker型鉀離子通道。根據(jù)電壓依賴特征和鉀離子跨膜流動方向,Shaker型K+通道又可劃分為3類:內(nèi)向鉀離子通道(Kin),主要包括AKT1和KAT1類鉀通道;外向鉀離子通道(Kout),主要包括SKOR和GORK類鉀通道;以及弱整流型鉀離子通道(Kweak),主要是AKT2鉀通道。與內(nèi)向、外向鉀離子通道不同,弱整流型鉀離子通道AKT2在S4跨膜區(qū)上的關(guān)鍵氨基酸均為帶正電的賴氨酸K (圖1b),這可能對其整流性有重要作用。系統(tǒng)進(jìn)化樹的結(jié)果顯示,水稻OsAKT2在進(jìn)化上與單子葉植物玉米的ZMK2的親緣關(guān)系最近,大麥HvAKT2次之,與雙子葉植物擬南芥的AtAKT2 (56%)等AKT2家族成員也有較高的相似度(圖1c)。綜上,水稻鉀通道OsAKT2與AKT2的其它家族成員具有較高的同源性,且氨基酸序列在進(jìn)化上具有保守性。

圖1 不同Shaker型K+通道的關(guān)鍵序列比對及系統(tǒng)進(jìn)化樹分析Fig.1 Sequence alignment and phylogenetic tree analysis of Shaker K+ channels
2.2 OsAKT2的鉀濃度響應(yīng)
弱整流型擬南芥AtAKT2通道可以同時介導(dǎo)K+的吸收和外排,且電壓依賴性較弱[14,19]。借助雙電極電壓鉗電生理技術(shù),發(fā)現(xiàn)水稻OsAKT2主要介導(dǎo)K+的吸收,但是缺少明顯的外排活性(圖2a)。從電流-電壓的關(guān)系可看出,當(dāng)外界鉀離子濃度高于5 mmol/L時,OsAKT2的電流開始顯著增加,而K+濃度達(dá)到100 mmol/L時電流基本接近飽和值(圖2b和c),反映了其對外界K+的吸收具有反饋調(diào)節(jié)機(jī)制。在 50 mmol/L K+且膜電位大于–50 mV 時,記錄到明顯的內(nèi)向電流,且隨著膜電位的降低,電流變得更大(圖2c)。OsAKT2的電流具有濃度依賴性和電壓依賴性。盡管OsAKT2在生物信息學(xué)和結(jié)構(gòu)分析上屬于AKT2通道,但是OsAKT2的電流性質(zhì)與已報道的擬南芥弱整流型AtAKT2明顯不同。
用米氏方程擬合OsAKT2的K+吸收動力學(xué)曲線,發(fā)現(xiàn)在各個膜電位下,Km值(達(dá)到1/2最大吸收速率時的鉀濃度)均大于 43 mmol/L (圖 2c),是一個典型的低親和力鉀離子通道(在大于1 mmol/L K+濃度下介導(dǎo)K+吸收) (圖2d)。在葉片衰老過程中,老葉中的K+要運(yùn)輸至生長旺盛的葉子、果實(shí)及種子中,可能會造成細(xì)胞內(nèi)K+釋放,導(dǎo)致老葉中K+瞬時增加達(dá)到10 mmol/L[20],而OsAKT2正是在此濃度范圍下發(fā)揮最適作用,揭示OsAKT2可能在地上部分K+回流和分配中有重要作用。
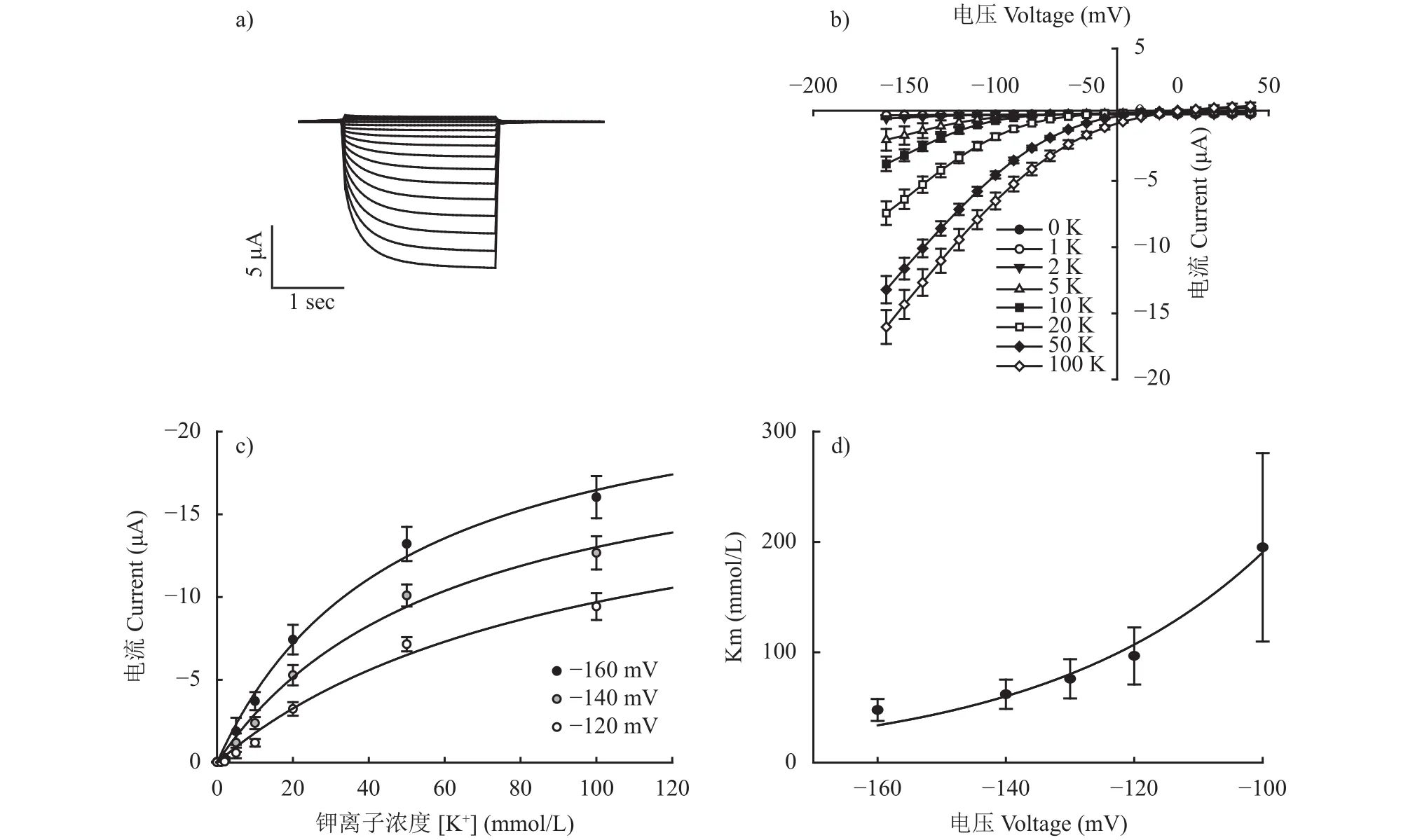
圖2 OsAKT2的鉀吸收特征Fig.2 K+ transport kinetics in OsAKT2 expressing Xenopus oocytes
2.3 OsAKT2的離子選擇性
對比與K+半徑相近的堿金屬陽離子通透性發(fā)現(xiàn),在不同膜電位下,OsAKT2對K+的吸收具有較高的選擇性,基本不能通透Li+和Na+,對Rb+有輕微的通透能力(圖3)。OsAKT2對NH4+有很大的通透性,約占K+吸收量的22%左右(以O(shè)sAKT2對K+的吸收量為標(biāo)準(zhǔn),100%),推測OsAKT2可能參與體內(nèi)NH4+的再分配,在緩解體內(nèi)NH4+毒害方面有一定作用。
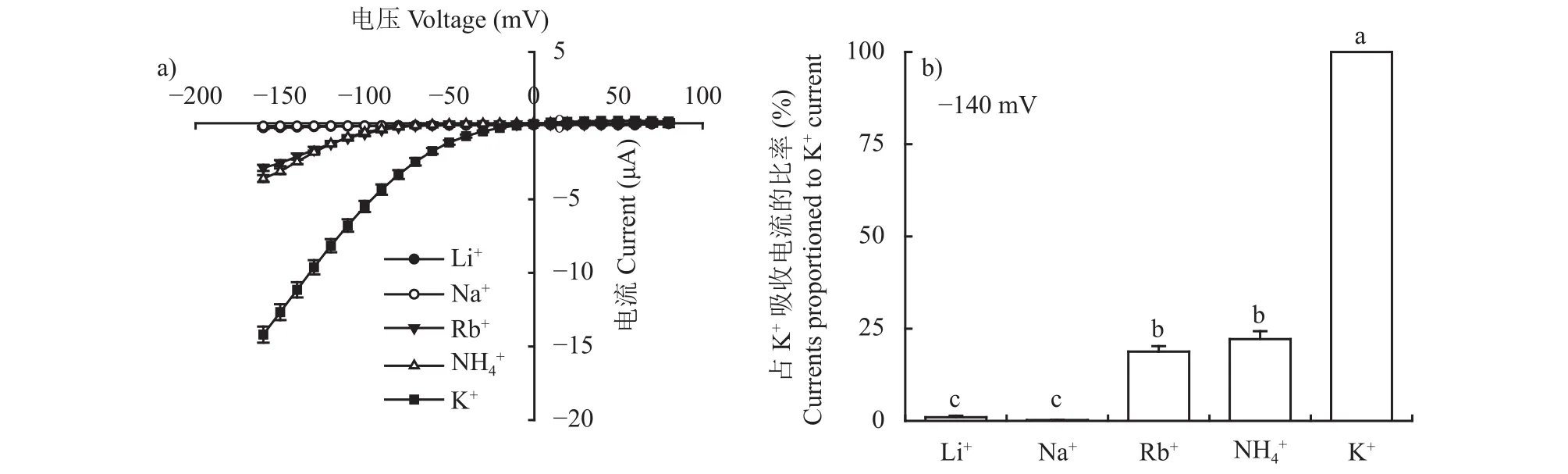
圖3 OsAKT2對一價陽離子的吸收電流?電壓曲線(a)和一價陽離子吸收電流相對于K+吸收電流的比例(b)Fig.3 I?V curves of monovalent ion uptake by OsAKT2 (a) and the current proportion of monovalent ions relative to that of K+ (b)
2.4 鉀離子通道抑制劑對OsAKT2的影響
我們檢測了K+通道專性抑制劑Ba2+、Cs+和TEA對OsAKT2通道活性的影響,以驗(yàn)證在蛙卵表達(dá)系統(tǒng)中產(chǎn)生的電流是否為專一性的鉀吸收電流。結(jié)果顯示,Ba2+、Cs+對OsAKT2電流的抑制作用具有電壓依賴性(圖4a),膜電位越負(fù),抑制效果越顯著,在膜電位從–80 mV到–160 mV的范圍內(nèi),Cs+對OsAKT2電流的抑制率從42%增加到了94%;Ba2+對OsAKT2電流的抑制率雖然從42%增加到了78%(圖4b),但較其他高度專一的K+通道來說抑制率較低,這與圖3中OsAKT2對離子的通透性特征相吻合(主要通透K+,但是也可以通透一定量的NH4+)(圖4c)。而TEA對OsAKT2電流的抑制作用沒有電壓依賴性,在膜電位從–80 mV到–160 mV的范圍內(nèi),TEA的抑制率基本維持在70%~79% (圖4d)。總之,OsAKT2通道活性受K+通道抑制劑較高程度的抑制,表明OsAKT2產(chǎn)生的電流為專一性的鉀吸收電流(圖4)。

圖4 OsAKT2在鉀通道抑制劑Ba2+、Cs+和TEA處理下的電流?電壓曲線(a)和穩(wěn)態(tài)電流(b、c、d)Fig.4 I?V curves (a) and steady currents (b, c, d) of the OsAKT2 by the external application of K+ channel blockers Ba2+, Cs+ and TEA
2.5 不同供銨、供鉀及脅迫處理對OsAKT2基因表達(dá)豐度的影響
為了進(jìn)一步揭示OsAKT2的作用條件及分子機(jī)制,測定了不同供銨、供鉀及干旱脅迫(模擬葉片衰老的過程)條件下OsAKT2的表達(dá)特征。模擬晝夜光照變化,發(fā)現(xiàn)OsAKT2的基因表達(dá)受光照條件影響,在光照條件下其表達(dá)豐度較低,而在黑暗中OsAKT2表達(dá)水平較高(圖5),具有一定的晝夜節(jié)律。在處理20 h時OsAKT2基因的表達(dá)豐度達(dá)到最高,因此在該處理時間進(jìn)行各處理的基因豐度比較。缺鉀(圖5a)和缺銨(圖5b)處理均會顯著增加OsAKT2的表達(dá)量,而OsAKT2主要介導(dǎo)地上部K+的吸收(圖2),推測OsAKT2在水稻氮、鉀營養(yǎng)虧缺時可促進(jìn)地上部分離子的回流以保障植物完整的生長發(fā)育進(jìn)程。此外,相比于正常條件,利用sorbitol(山梨醇)處理模擬滲透脅迫增加了OsAKT2基因的表達(dá)豐度,PEG處理無顯著效果(圖5c)。在不同脅迫處理?xiàng)l件下均未改變OsAKT2的晝夜節(jié)律變化,這可能是植株應(yīng)對脅迫條件的一種策略。

圖5 不同處理?xiàng)l件下OsAKT2基因的表達(dá)豐度Fig.5 OsAKT2 gene expression level in response to different treatments
3 討論與結(jié)論
植物Shaker型鉀通道是一類電壓門控的K+離子通道,在K+的吸收、轉(zhuǎn)運(yùn)以及維持細(xì)胞內(nèi)離子動態(tài)平衡等方面發(fā)揮重要作用[21]。盡管OsAKT2在結(jié)構(gòu)和基本性質(zhì)上都與典型的AKT2鉀通道相似,如二者在氨基酸序列上高度同源,且表現(xiàn)出較高的鉀離子選擇性,同屬于弱整流型Shaker類鉀通道(圖1),但是其也表現(xiàn)出一些特殊電流性質(zhì)。利用非洲爪蟾卵母細(xì)胞作為外源表達(dá)系統(tǒng),發(fā)現(xiàn)OsAKT2主要介導(dǎo)K+的吸收,缺少明顯的外向活性,且表現(xiàn)出明顯的電壓依賴性(圖2),這與已知的擬南芥AtAKT2的“雙向”電流明顯不同[14]。AtAKT2的外向活性(不受電壓影響)有其特殊的生理意義:K+的外流能夠阻止韌皮部細(xì)胞內(nèi)由于H+內(nèi)流引起的過度去極化,進(jìn)而通過蔗糖/H+共轉(zhuǎn)運(yùn)體驅(qū)動蔗糖的運(yùn)輸[22]。而水稻OsAKT2缺少明顯的外排活性,意味著其可能缺少蔗糖運(yùn)輸?shù)墓δ堋A硗猓芯勘砻鱋sAKT2主要定位在韌皮部[18],且負(fù)責(zé)K+的吸收,那么OsAKT2很可能參與了將植株地上部分多余的K+經(jīng)由韌皮部運(yùn)往根系,一方面給韌皮部負(fù)載提供所需的營養(yǎng)物質(zhì),另一方面也作為地上部鉀營養(yǎng)狀況的重要信號對根系吸收K+進(jìn)行反饋調(diào)節(jié)。植株地上部分的K+回流有利于水稻等糧食作物維持其生長部位的功能,但是對于農(nóng)業(yè)生產(chǎn)中需要保持葉片高鉀水平的作物如煙草等較為不利。
OsAKT2對于維持細(xì)胞內(nèi)K+濃度穩(wěn)定發(fā)揮著關(guān)鍵作用。本研究發(fā)現(xiàn)OsAKT2對外界K+濃度的變化很敏感,且顯示出典型的低親和力特性(Km為43 mmol/L,遠(yuǎn)大于高親和系統(tǒng)介導(dǎo)的0.2~1 mmol/L K+的吸收[23]),與已知的其它Shaker型鉀通道一致[24]。植物細(xì)胞質(zhì)中的K+濃度一般維持在50~100 mmol/L范圍內(nèi)[25],這種最佳的K+濃度有利于細(xì)胞質(zhì)中酶的活化[9]。水稻OsAKT2對K+具有較高的選擇性(圖3),且會抑制根系對Na+的吸收[17],表明OsAKT2可能有助于提高植物的抗鹽能力。此外,水稻OsAKT2對NH4+表現(xiàn)出一定的通透性(圖3),且缺銨處理會顯著上調(diào)OsAKT2的表達(dá)水平,說明OsAKT2可能對水稻氮素營養(yǎng)吸收轉(zhuǎn)運(yùn)具有潛在的貢獻(xiàn)。植株內(nèi)多余的NH4+或可通過OsAKT2再回流到根系并進(jìn)行氨同化,降低植株體內(nèi)游離NH4+含量,從而緩解NH4+毒害。這一結(jié)果對于將來研究水稻體內(nèi)NH4+回流及再分配有一定的參考價值。
OsAKT2基因的表達(dá)呈現(xiàn)出明顯的避光性,與AtAKT2基因受光照誘導(dǎo)的特性相反,說明水稻和擬南芥兩種植物具有不同的晝夜節(jié)律和光合產(chǎn)物分配機(jī)制。擬南芥AtAKT2鉀通道參與的蔗糖分配過程具有光依賴性[15],AKT2的一個重要作用就是激活韌皮部質(zhì)膜協(xié)助光合產(chǎn)物的向下運(yùn)輸,從而有效的裝載糖類物質(zhì)[26]。而水稻OsAKT2基因在光照下表達(dá)較弱(圖5),且OsAKT2功能缺失并不影響水稻地上部植株的蔗糖含量[17],表明OsAKT2可能不參與協(xié)助蔗糖的運(yùn)輸,這也與OsAKT2缺少明顯的外向活性及其可能對應(yīng)的蔗糖運(yùn)輸功能這一特征相一致(圖2)。黑暗條件下,氣孔關(guān)閉不再提供蒸騰拉力,植物為了滿足正常的生長需求,其體內(nèi)存在一定的離子循環(huán),此時OsAKT2的表達(dá)豐度升高(圖5),顯示OsAKT2具有潛在的K+回流作用。這些不同植物之間的特異性節(jié)律變化可能是植物表現(xiàn)出的適應(yīng)性進(jìn)化,具體生理機(jī)制有待進(jìn)一步探索。本研究中基因表達(dá)試驗(yàn)采用的是10天苗齡(2~3葉期)的水稻幼苗,該結(jié)果可以在一定程度上反映OsAKT2介導(dǎo)地上部分的K+回流,而老葉到新葉的K+回流可能需在4~5葉期的幼苗中進(jìn)行驗(yàn)證,在此時期,第一片葉已開始進(jìn)入衰老階段,便于新的完全展開葉和老葉之間進(jìn)行比較。
缺氮、缺鉀以及干旱等脅迫環(huán)境因素會加速葉片老化,縮短植物生育期。本研究發(fā)現(xiàn)在這類脅迫條件下OsAKT2基因的表達(dá)水平會顯著上調(diào),這可能有助于K+從衰老器官運(yùn)輸?shù)缴L旺盛的新器官,反映了植物對外界環(huán)境的適應(yīng)性生存策略。相關(guān)研究也發(fā)現(xiàn),過表達(dá)大麥HvAKT2和HvHAK1能夠通過調(diào)節(jié)葉片H+內(nèi)穩(wěn)態(tài)和細(xì)胞信號傳導(dǎo)來提高大麥的耐旱性[27]。而對于喜鉀作物煙草來說,保持葉片充足的K+是其品質(zhì)保證,沈方科等[28]發(fā)現(xiàn)環(huán)割或斷根處理可以阻斷物質(zhì)在韌皮部的運(yùn)輸,從而減少K+向地下部分回流,提高煙葉鉀含量。綜上,水稻OsAKT2可能在地上部K+的再分配過程中發(fā)揮重要作用,這為通過超表達(dá)OsAKT2來提高植物抗逆性提供了可能。
猜你喜歡
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
