愈創(chuàng)木基丙三醇-β-愈創(chuàng)木基醚對(duì)土壤中多環(huán)芳烴的歸趨的影響及微生物群落效應(yīng)的研究
2022-09-21 03:34:08季聰杰安雪暉陳禹竹
安徽農(nóng)業(yè)科學(xué) 2022年17期
尹 晗,季聰杰,安雪暉,孫 月,陳禹竹*
(1.中電建路橋集團(tuán)有限公司,北京 100048;2.清華大學(xué)水利水電工程系,北京 100084;3.中國(guó)科學(xué)院南京土壤研究所,江蘇南京 210008)
多環(huán)芳烴(PAHs)是含有2個(gè)或2個(gè)以上苯環(huán)連接在一起的烴類(lèi)化合物,隨著其環(huán)數(shù)增加、化學(xué)結(jié)構(gòu)的變化和疏水性的增強(qiáng),其化學(xué)穩(wěn)定性、持久性、抗生物降解能力和毒性都會(huì)增強(qiáng),多環(huán)芳烴已成為我國(guó)土壤中主要的有機(jī)污染物之一,被認(rèn)定為影響人類(lèi)健康的主要有機(jī)污染物。一些高分子量有機(jī)物不能直接作為大部分微生物的唯一碳源和能源,在有其他化合物作為一級(jí)底物提供碳源和能源時(shí),該有機(jī)物才能作為二級(jí)底物被降解,這種現(xiàn)象稱為共代謝作用,這類(lèi)一級(jí)底物被稱為共代謝底物。多環(huán)芳烴是一類(lèi)難降解的有機(jī)污染物,分子量高,很少有微生物可以直接利用其作為唯一碳源,因此,高分子量多環(huán)芳烴通常以共代謝方式進(jìn)行降解,共代謝過(guò)程已作為一種生物技術(shù)在芳香族化合物生物降解修復(fù)中得到廣泛應(yīng)用。
木質(zhì)素是一類(lèi)復(fù)雜的有機(jī)聚合物,其化學(xué)結(jié)構(gòu)與多環(huán)芳烴結(jié)構(gòu)類(lèi)似,已有研究表明,木質(zhì)素具有刺激微生物降解芳香族污染物的作用。木質(zhì)素解聚產(chǎn)生大量雙芳基或單芳基降解產(chǎn)物,它們的進(jìn)一步代謝會(huì)誘導(dǎo)微生物產(chǎn)生胞外酶等物質(zhì),這些可能會(huì)促進(jìn)多環(huán)芳烴降解。木質(zhì)素促進(jìn)芳香族污染物的降解過(guò)程,可能涉及共代謝機(jī)制,但其具體的微生物共代謝機(jī)制尚不明確。木質(zhì)素在土壤中的降解過(guò)程涉及多種微生物,降解的不同階段也具有不同的共代謝效應(yīng)。目前解決多環(huán)芳烴的污染問(wèn)題以及深入地研究木質(zhì)素促進(jìn)多環(huán)芳烴降解的微生物共代謝機(jī)制對(duì)研究多環(huán)芳烴污染土壤的修復(fù)具有重要意義。因此,該研究選擇了一種木質(zhì)素模型二聚體化合物愈創(chuàng)木基丙三醇-β-愈創(chuàng)木基醚(guaiacylglycerol-β-guaiacyl ether,GGE)為刺激底物,以多環(huán)芳烴苯并[a]蒽(Benz(a)anthracene,BaA)為模式污染物,同時(shí)結(jié)合真菌抑制劑放線菌酮(cycloheximide),設(shè)立微宇宙培養(yǎng)試驗(yàn),結(jié)合C同位素示蹤技術(shù)、定量PCR技術(shù)和擴(kuò)增子測(cè)序來(lái)分析GGE對(duì)BaA的歸趨的影響及其微生物群落效應(yīng)。
1 材料與方法
土壤采集于南京市郊農(nóng)田,自然風(fēng)干,研磨后過(guò)2 mm網(wǎng)篩,保存于4 ℃,用于后續(xù)培養(yǎng)試驗(yàn)。土壤理化性質(zhì):pH 6.84、總碳12.7 g/kg、總氮1.3 g/kg、全磷0.57 g/kg、全鉀19.3 g/kg、田間持水量33.3%。
[7,12-C]苯并[a]蒽(C-BaA,10 mCi/mmol,99%純度),美國(guó)Radiolabeled Chemicals公司;苯并[a]蒽(C-BaA,98%純度),梯希愛(ài)(上海)化成工業(yè)發(fā)展有限公司;真菌抑制劑放線菌酮,西格瑪奧德里奇(上海)貿(mào)易有限公司。液體閃爍分析儀(LSC),美國(guó)貝克曼庫(kù)爾特有限公司;生物氧化燃燒儀(OX-500),德國(guó)Zinsser Analytic公司。
設(shè)立2組微宇宙培養(yǎng)試驗(yàn),2組處理相同,微宇宙培養(yǎng)試驗(yàn)在60 mL礦化管中進(jìn)行,每管加入5.0 g土壤樣品,土壤持水量為60%,處理組添加GGE的含量為500 μg/g,真菌抑制劑放線菌酮的含量為1 mg/g。
第一組通過(guò)同位素示蹤技術(shù),檢測(cè)GGE處理組和對(duì)照組在有無(wú)放線菌酮存在的情況下C-BaA的環(huán)境歸趨,每個(gè)處理設(shè)3個(gè)平行。添加C標(biāo)記的BaA至土壤樣品中,使其終濃度為50 μg/g,放射性為2.0×10DPM。在礦化管內(nèi)放置裝有1 mL 1 mol/L NaOH溶液的小瓶子,收集釋放的CO,培養(yǎng)時(shí)間為70 d,每7 d取樣測(cè)定培養(yǎng)過(guò)程中釋放的CO量,利用液體閃爍分析儀(LSC)測(cè)定NaOH溶液吸收的CO含量。
培養(yǎng)結(jié)束后,將礦化管內(nèi)的土壤樣品風(fēng)干,過(guò)2 mm網(wǎng)篩,取2.0 g用二氯甲烷在50 ℃下索氏提取24 h,將萃取物旋蒸后用環(huán)己烷熔解,經(jīng)LSC測(cè)定可提取態(tài)(DCM)含量。不可提取態(tài)(NER)進(jìn)一步分離為富里酸(FA)、腐殖酸(HA)和胡敏素(HU)結(jié)合態(tài)。參照Shan等的方法,將DCM提取后的土樣風(fēng)干,用0.1 mol/L無(wú)氧NaOH在250 r/min下振蕩提取24 h,12 000 r/min離心30 min,所得沉淀為胡敏素,上清液為富里酸和腐殖酸的混合溶液,在上清液中加入6 mol/L HCl調(diào)節(jié)混合溶液pH至1,4 ℃下沉淀24 h,7 000 r/min離心30 min,上清液為FA組分,利用LSC檢測(cè)FA和HA組分的放射性,得到FA、HA結(jié)合態(tài)的含量。將胡敏素沉淀冷凍干燥后,用生物氧化燃燒儀在900 ℃下完全氧化4 min,收集氧化后產(chǎn)生的C標(biāo)記的CO,并用LSC檢測(cè)HU組分放射性,得到HU結(jié)合態(tài)的含量。
第二組添加未標(biāo)記的C-BaA至土壤樣品中,使其終濃度為50 μg/g,用于檢測(cè)培養(yǎng)過(guò)程中的微生物變化,每個(gè)處理6個(gè)重復(fù),分別在培養(yǎng)中期(第42天)和培養(yǎng)結(jié)束(第70天)時(shí),各取3支破壞性取樣。土壤樣品置于-20 ℃保存,用于后續(xù)微生物分析。使用FastDNA SPIN Kit for Soils土壤DNA提取試劑盒,參照試劑盒說(shuō)明書(shū)操作,提取土壤樣品中的DNA。測(cè)定DNA濃度后,置于-20 ℃保存。
采用定量PCR的方法,測(cè)定各樣品中細(xì)菌16S rRNA基因、18S rRNA基因、甲醇脫氫酶基因和革蘭氏陽(yáng)性菌多環(huán)芳烴雙加氧酶GP PAH-RHDα 基因表達(dá)豐度。土壤樣品總DNA稀釋至10濃度,作為定量模板,定量PCR反應(yīng)體系 (20 μL):Top Green qPCR SuperMix (SYBR Green) 10.0 μL,正、反向引物 (10 μmol/L)各0.4 μL,ddHO 7.2 μL,模板DNA 2.0 μL。采用細(xì)菌16S rRNA基因序列PCR通用引物Eub338F (5′-ACTCCTACGGGAGGCAGCAG-3′)和Eub518R (5′-ATTACCGCGGCTGCTGG-3′),擴(kuò)增目的片段;定量PCR擴(kuò)增程序:95 ℃ 3 min;95 ℃ 30 s,56 ℃ 30 s,72 ℃ 30 s,35 個(gè)循環(huán);72 ℃ 10 min。采用真菌18S rRNA基因序列PCR通用引物nu-SSU-0817 (5′-TTAGCATGGAATAATRRAATAGGA-3′)和nu-SSU-1536 (5′-ATTGCAATGCYCTATCCCCA-3′),擴(kuò)增目的片段;定量PCR擴(kuò)增程序:94 ℃ 3 min;94 ℃ 30 s,53 ℃ 30 s,72 ℃ 30 s,35 個(gè)循環(huán);72 ℃ 10 min。采用基因序列PCR通用引物mdh1 (5′-GCGGIWSCAICTGGGGYT -3′)、 mdh2 (5′-GCGGIWSGAICTGGGGYT -3′)和mdhR (5′-GAASGGYTCSYARTCCATGCA -3′),擴(kuò)增目的片段;定量PCR擴(kuò)增程序:95 ℃ 5 min;95 ℃ 1 min,53 ℃ 1 min,72 ℃ 1 min,40 個(gè)循環(huán);72 ℃ 10 min。采用GP PAH-RHDα基因序列PCR通用引物GP RHDα F (5′-CGGCGCCGACAAYTTYGTNGG -3′)和GP RHDα R (5′-GGGGAACACGGTGCCRTGDATRAA-3′),擴(kuò)增目的片段;定量PCR擴(kuò)增程序:95 ℃ 3 min;95 ℃ 20 s,54 ℃ 30 s,72 ℃ 30 s,40 個(gè)循環(huán);72 ℃ 10 min。
利用Illumina Miseq高通量測(cè)序技術(shù),對(duì)細(xì)菌16S rRNA基因和真菌ITS基因測(cè)序,分析土壤細(xì)菌和真菌群落組成。測(cè)序工作送至上海美吉生物醫(yī)藥科技有限公司完成。所得原始數(shù)據(jù)使用QIIME (V1.9.1)和USEARCH (V8.1.1861)流程進(jìn)行分析,序列經(jīng)拼接、過(guò)濾和質(zhì)控后,在97%相似性水平劃分操作分類(lèi)單元(operational taxonomic unit,OTU),細(xì)菌序列通過(guò)與SILVA數(shù)據(jù)庫(kù)(https://www.arb-silva.de)比對(duì)確定系統(tǒng)學(xué)分類(lèi),真菌序列通過(guò)與Unite數(shù)據(jù)庫(kù)(http://unite.ut.ee/)比對(duì)確定系統(tǒng)學(xué)分類(lèi),得到每個(gè)樣品的OTUs和物種注釋的基本分析結(jié)果,同時(shí)對(duì)物種注釋在各個(gè)分類(lèi)水平上進(jìn)行組成結(jié)構(gòu)的統(tǒng)計(jì)分析。再對(duì)OTUs進(jìn)行豐度、多樣性指數(shù)等分析。
利用SPSS 20.0、 Excel 2019和Graphpad Prism 8對(duì)數(shù)據(jù)進(jìn)行整理分析與作圖,利用Galaxy平臺(tái)(http://huttenhower.sph.harvard.edu/galaxy/)進(jìn)行LefSe分析。
云工作流引擎的設(shè)計(jì)目標(biāo)主要是將工作流引擎從應(yīng)用系統(tǒng)中解耦出來(lái),形成一個(gè)獨(dú)立的服務(wù),從而降低系統(tǒng)的耦合性,提供更加靈活的服務(wù)支持,實(shí)現(xiàn)復(fù)雜度可控、獨(dú)立部署、技術(shù)選型靈活、服務(wù)容錯(cuò)和按需獨(dú)立擴(kuò)展等目標(biāo)。
2 結(jié)果與分析
通過(guò)同位素示蹤技術(shù),測(cè)定C-BaA在土壤中的歸趨情況,如圖1所示。在培養(yǎng)的前期階段,礦化率呈現(xiàn)CK>GGE>CK-cycloheximide>GGE-cycloheximide的趨勢(shì),在70 d的培養(yǎng)結(jié)束后,GGE的礦化率最高,達(dá)到4.61%,CK的礦化率為4.43%,略低于GGE。除礦化部分外,大部分的C-BaA可被二氯甲烷提取,CK組和GGE處理組中的可提取態(tài)C-BaA分別為71.17%和70.23%。不可提取態(tài)進(jìn)一步細(xì)分為富里酸(FA)、腐殖酸(HA)和胡敏素(HU)結(jié)合態(tài),在CK組中,與富里酸、腐殖酸和胡敏素結(jié)合的比例分別為3.37%、6.55%和14.10%,在GGE處理組中,與富里酸、腐殖酸和胡敏素結(jié)合的比例分別為3.13%、6.78%和14.75%。
在CK組和GGE處理組中,真菌抑制劑放線菌酮(cycloheximide)的添加對(duì)BaA在土壤中的歸趨有不同程度的影響。CK組受到放線菌酮的影響較小,礦化基本未受影響,除腐殖酸結(jié)合態(tài)減少了13.13%,其他組分變化很小。在GGE處理組中,加入放線菌酮后,礦化率減少,僅為3.50%,礦化率降低了22.35%,富里酸結(jié)合態(tài)增加了29.71%,胡敏素結(jié)合態(tài)增加了15.39%。

圖1 培養(yǎng)過(guò)程中的14CO2礦化情況(A)以及培養(yǎng)結(jié)束時(shí)14C-BaA的歸趨(B) Fig.1 The mineralization of 14CO2 during the incubation(A) and the distribution of 14C-BaA at the end of incubation (B)
采用定量PCR的方法,檢測(cè)了土壤中細(xì)菌16S rRNA基因、18S rRNA基因和革蘭氏陽(yáng)性細(xì)菌多環(huán)芳烴雙加氧酶GP PAH-RHDα基因,結(jié)果如圖2所示。不添加放線菌酮的情況下,細(xì)菌16S rRNA基因中,在CK組和GGE處理組的培養(yǎng)中期的基因拷貝數(shù)分別為1.02×10和8.89×10copies/g,培養(yǎng)結(jié)束后的基因拷貝數(shù)分別為3.57×10和3.92×10copies/g,CK和GGE處理之間未見(jiàn)顯著差異;真菌18S rRNA基因中,GGE處理組相對(duì)于CK組提高了基因拷貝數(shù),從1.92×10copies/g提高至2.70×10copies/g,培養(yǎng)結(jié)束后,拷貝數(shù)均明顯下降,且處理間相差不大;與CK組相比,GGE處理組也提高了甲醇脫氫酶的基因拷貝數(shù),在培養(yǎng)期間從1.09×10copies/g提高至1.79×10copies/g,培養(yǎng)結(jié)束后整體上拷貝數(shù)減少;在GP PAH-RHDα基因中,隨著培養(yǎng)的進(jìn)行,CK組的基因拷貝數(shù)逐漸減少,從1.78×10copies/g減少至1.17×10copies/g,而GGE處理組的拷貝數(shù)逐漸增加,從1.33×10copies/g增加至1.75×10copies/g。放線菌酮添加的情況下,整體上沒(méi)有明顯減少土壤中細(xì)菌16S rRNA基因、真菌18S rRNA基因和基因的數(shù)量,但在GP PAH-RHDα基因中,放線菌酮的添加顯著抑制了GGE處理組中基因豐度。
對(duì)細(xì)菌16S rRNA基因的高通量測(cè)序結(jié)果進(jìn)行分析,一共獲得了55萬(wàn)條序列,經(jīng)質(zhì)量控制,單個(gè)樣品的序列數(shù)在15 978~34 374,在97%序列相似性水平上獲得24 750個(gè)OTU。對(duì)真菌ITS基因的高通量測(cè)序結(jié)果進(jìn)行分析,一共獲得了78萬(wàn)條序列,經(jīng)質(zhì)量控制,單個(gè)樣品的序列數(shù)在24 393~71 441,在97%序列相似性水平上獲得1 579個(gè)OTU。
對(duì)相對(duì)豐度>1%的細(xì)菌門(mén)/綱類(lèi)進(jìn)行分析,結(jié)果發(fā)現(xiàn)(圖3A),在第42天,CK組中主要為α-變形菌綱(Alpha-proteobacteria)、β-變形菌綱(Beta-proteobacteria)、酸桿菌門(mén)(Acidobacteria)、放線菌門(mén)(Actinobacteria)、厚壁菌門(mén)(Firmicutes)和γ-變形菌綱(Gamma-proteobacteria),占比分別為14.47%、17.07%、21.47%、12.14%、5.93%和4.97%。GGE處理組增加了α-變形菌門(mén)、擬桿菌門(mén)(Bacteroidetes)的相對(duì)豐度,而β-變形菌綱和芽單胞菌門(mén)(Gemmatimonadetes)則有所減少。各類(lèi)細(xì)菌門(mén)/綱的相對(duì)豐度在培養(yǎng)時(shí)間上也存在著一定的變化,第70天與第42天相比,CK和GGE處理下酸桿菌門(mén)和α-變形菌綱的相對(duì)豐度明顯增加,β-變形菌綱、放線菌門(mén)和厚壁菌門(mén)的比例均有減少。
對(duì)真菌在門(mén)水平上進(jìn)行分析,結(jié)果發(fā)現(xiàn)(圖3B),在第42天時(shí),CK組中主要真菌為子囊菌門(mén)(Ascomycota)、擔(dān)子菌門(mén)(Basidiomycota)和壺菌門(mén)(Chytridiomycota),占比分別為63.67%、23.60%和2.28%,GGE處理組與CK組相比,子囊菌門(mén)的相對(duì)豐度增加至73.85%,擔(dān)子菌門(mén)的相對(duì)豐度減少,為11.58%。各類(lèi)真菌的相對(duì)豐度在培養(yǎng)時(shí)間上也存在著一定的變化,到第70天時(shí),CK和GGE處理中的子囊菌門(mén)的相對(duì)豐度有所減少,CK組與第42天相比,被孢霉門(mén)(Mortierellomycota)明顯增加,GGE處理組中被孢霉門(mén)和球囊菌門(mén)(Glomeromycota)的相對(duì)豐度明顯增加。
在OUT水平上,對(duì)細(xì)菌和真菌的群落進(jìn)行非度量多維尺度(NMDS)分析(基于Bray-Curtis 距離),如圖4所示。在細(xì)菌群落中,第42天和第70天的樣品存在很明顯的分離,表明樣品在培養(yǎng)期間存在時(shí)間上的演替,GGE處理組的大部分樣品能相對(duì)集中在一起,且與CK組之間呈現(xiàn)出一定的分離,表明GGE的添加對(duì)細(xì)菌群落組成產(chǎn)生了一定的影響。在真菌群落中,GGE處理對(duì)真菌群落影響較小,2組樣品未發(fā)生分別聚類(lèi),與CK組的群落結(jié)構(gòu)相似,在不同的培養(yǎng)時(shí)間分布上,CK組未出現(xiàn)明顯分離,而GGE處理組呈現(xiàn)了一定程度的分離。
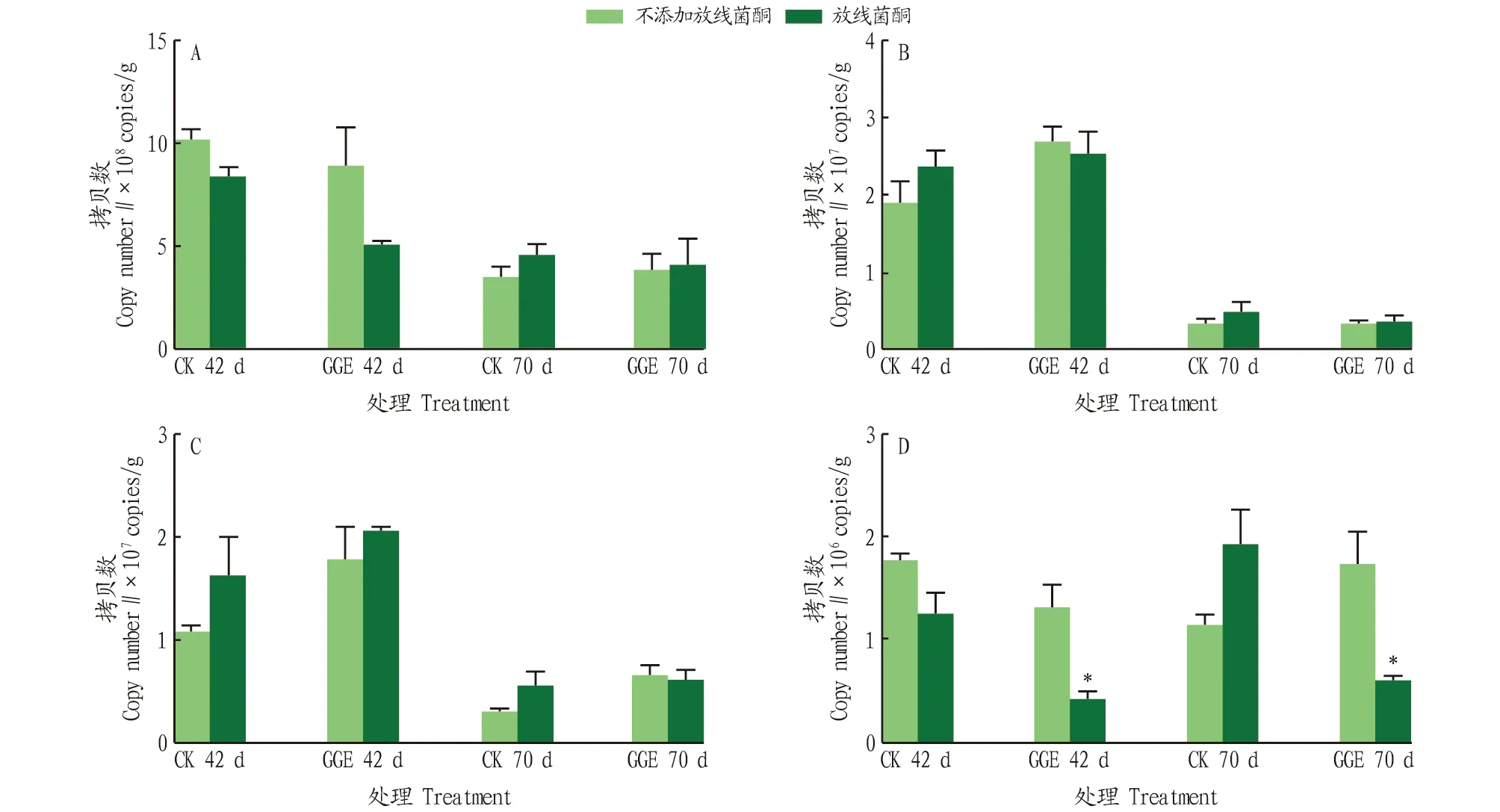
注:*表示在0.05水平上放線菌酮處理間差異顯著 Note:* indicates significant difference between cycloheximide treatments at the 0.05 level圖2 細(xì)菌(A)、真菌(B)、甲醇脫氫酶mxaF/xoxF(C)和 GP PAH-RHDα(D)基因的定量PCR結(jié)果Fig.2 Quantitative PCR results of bacteria(A) ,fungi (B),methanol dehydrogenase mxaF/xoxF(C) and GP PAH-RHDα genes (D)

圖3 細(xì)菌(A)和真菌(B)在門(mén)/綱水平上的相對(duì)豐度Fig.3 Relative abundance of bacteria (A)and fungi(B) at phylum or class level

圖4 細(xì)菌 (A)和真菌(B)的非度量多維尺度(NMDS)分析Fig.4 Non-metric multidimensional scaling (NMDS) analysis of bacteria (A) and fungi (B)

注:圖中結(jié)點(diǎn)從內(nèi)圈至外圈代表各個(gè)分類(lèi)水平(界、門(mén)、綱、目、科、屬),不同顏色代表在各自處理中顯著富集的菌屬 Note:The nodes in graphs from the inner ring to the outer ring represent the different taxonomic levels (domain,phylum,class,order,family and genus),different colors represent the significant enrichment in respective treatments圖5 不同處理的微生物群落LefSe分析Fig.5 LefSe analysis of microbial communities from different treatments
3 討論與結(jié)論
木質(zhì)素是一種芳香性聚合物,被解聚后會(huì)有不同的芳基單體產(chǎn)生,因其與多環(huán)芳烴都含有芳香性結(jié)構(gòu),可能會(huì)成為多環(huán)芳烴降解過(guò)程中的共代謝底物,從而促進(jìn)污染物的代謝。在該研究中,通過(guò)比較對(duì)照(CK)組和添加木質(zhì)素的一種模式化合物的GGE處理組對(duì)土壤中BaA歸趨的影響,并通過(guò)添加真菌抑制劑,來(lái)驗(yàn)證污染物歸趨過(guò)程中細(xì)菌和真菌的貢獻(xiàn)。從BaA的歸趨結(jié)果來(lái)看,CK組與GGE處理組的礦化率相近,在CK組中加入放線菌酮對(duì)礦化幾乎沒(méi)有影響,而與CK組相比,GGE處理受到了放線菌酮的抑制,礦化率減少了22.35%。礦化過(guò)程主要與微生物群落有關(guān),認(rèn)為在CK組中真菌起到的作用不大,但是加入了GGE之后,真菌的作用增大,盡管礦化結(jié)果并沒(méi)有顯著變化,但結(jié)合微生物群落的變化,GGE的加入在一定程度上使得真菌的貢獻(xiàn)在提升。
目前已知真菌與細(xì)菌使用完全不同的途徑對(duì)多環(huán)芳烴進(jìn)行轉(zhuǎn)化,真菌轉(zhuǎn)化多環(huán)芳烴的途徑主要是胞內(nèi)P450氧化酶系轉(zhuǎn)化途徑和胞外木質(zhì)素氧化酶轉(zhuǎn)化途徑;而細(xì)菌對(duì)多環(huán)芳烴的轉(zhuǎn)化一般屬于好氧降解途徑,其中雙加氧酶途徑是細(xì)菌降解多環(huán)芳烴最常見(jiàn)的方式。這些不同導(dǎo)致 PAHs 在轉(zhuǎn)化的中間產(chǎn)物和轉(zhuǎn)化效率等方面存在很大差異。該研究土壤微宇宙中BaA的礦化可能涉及真菌和細(xì)菌的協(xié)同作用。從定量的分析結(jié)果可以看到,在CK處理中加入放線菌酮,并未對(duì)多環(huán)芳烴雙加氧酶GP PAH-RHDα基因的豐度產(chǎn)生明顯影響,而在GGE處理中加入放線菌酮,則顯著抑制了雙加氧酶基因的豐度。真菌對(duì)PAHs的作用是多樣的,但真菌較少能直接利用高分子量的多環(huán)芳烴,更多是以共代謝機(jī)制降解PAHs,即利用其他碳源生長(zhǎng)的同時(shí),在次級(jí)代謝過(guò)程中轉(zhuǎn)化PAHs。培養(yǎng)體系中GGE的加入可能會(huì)刺激土壤中的真菌,使其先被利用,轉(zhuǎn)化成某些中間體化合物,它們的進(jìn)一步代謝可能會(huì)誘導(dǎo)土壤中的細(xì)菌表達(dá)雙加氧酶等,這些酶類(lèi)會(huì)作用于污染物,促進(jìn)降解過(guò)程。底物基質(zhì)的添加可能會(huì)引起多環(huán)芳烴的不同微生物降解機(jī)制。
根據(jù)該研究對(duì)微生物群落的分析結(jié)果,GGE的添加相比于真菌對(duì)細(xì)菌群落的影響更大。定量PCR的結(jié)果顯示,在GGE處理下,基因有一定程度的增加,甲醇脫氫酶是微生物甲基代謝中的關(guān)鍵酶之一,LEfSe的分析結(jié)果也表明,GGE處理組顯著富集了Hyphomicrobiaceae,已有研究發(fā)現(xiàn)為一種甲基營(yíng)養(yǎng)菌,一些甲基營(yíng)養(yǎng)菌已被證明可降解石油碳?xì)浠衔锖头枷阕逄細(xì)浠衔铮€可以利用廢氣中的多環(huán)芳香烴,甲基營(yíng)養(yǎng)菌這類(lèi)潛在降解菌的富集,可能也會(huì)對(duì)多環(huán)芳烴的降解有潛在能力。GGE處理組還顯著富集了Rhodospirillaceae、Sinobacteraceae、Acidobacteriaceae、Burkholderiaceae等與有機(jī)物降解相關(guān)的細(xì)菌科,真菌的富集主要集中在子囊菌門(mén)糞殼菌綱。已有研究表明,子囊菌門(mén)是污染環(huán)境中的主要真菌,具有轉(zhuǎn)化或去除PAHs的能力。張娟琴等從工廠附近的土壤中分離到一株可以降解PAHs的伯克氏菌(),且菌株具有較高的雙加氧酶活性。Rhodospirillaceae和Acidobacteriaceae等菌科也被研究發(fā)現(xiàn)是PAHs的潛在降解菌。這些微生物的富集可能會(huì)在整體上提升土壤中的芳香族化合物的代謝功能,增強(qiáng)對(duì)多環(huán)芳烴類(lèi)污染物的降解能力。
現(xiàn)階段,鑒于微生物群落的復(fù)雜響應(yīng),GGE處理對(duì)土壤微生物群落的調(diào)控與多環(huán)芳烴的轉(zhuǎn)化關(guān)系仍需進(jìn)一步分析,希望隨著研究的深入,將木質(zhì)素及其代謝產(chǎn)物的共代謝刺激作用機(jī)制進(jìn)一步的完善,為利用微生物群落降解環(huán)境污染物提供更多的科學(xué)依據(jù)。
